
High-Resolution Transect Sampling and Multiple Scale Diversity Analyses for Evaluating Grassland Resilience to Climatic Extremes

 , , , , , ,
, , , , , ,
Abstract
1. Introduction
- Q1
- Do temporal diversity patterns (trends or fluctuations) respond significantly to the temporal variation of weather characteristics?
- Q2
- Are these patterns synchronized and dependent on the spatial scale of observation?
- Q3
- Do plant functional groups differ in responses, and how do site conditions and the type of plant communities modulate the related patterns?
2. Materials and Methods
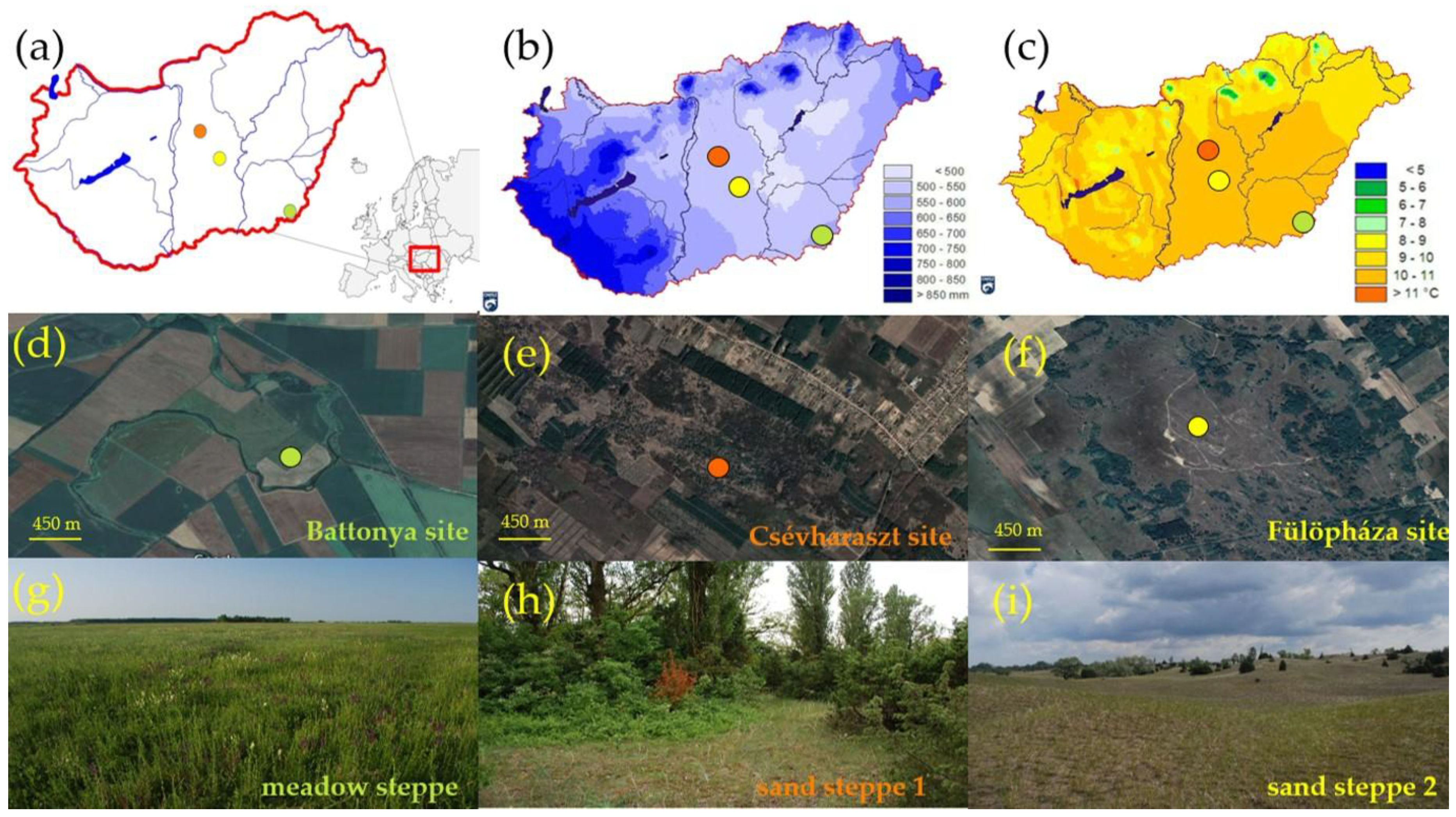
2.1. Study Sites
2.2. Vegetation Monitoring and Computerized Sampling for Species Richness Data
2.3. Meteorological Data
2.4. Analyses
2.4.1. Testing for Trends in Weather and Temporal Diversity Patterns
2.4.2. Testing for Spatial Synchrony of Temporal Patterns among and within Sites
2.4.3. Testing Relationships between Species Richness and Weather Characteristics
3. Results
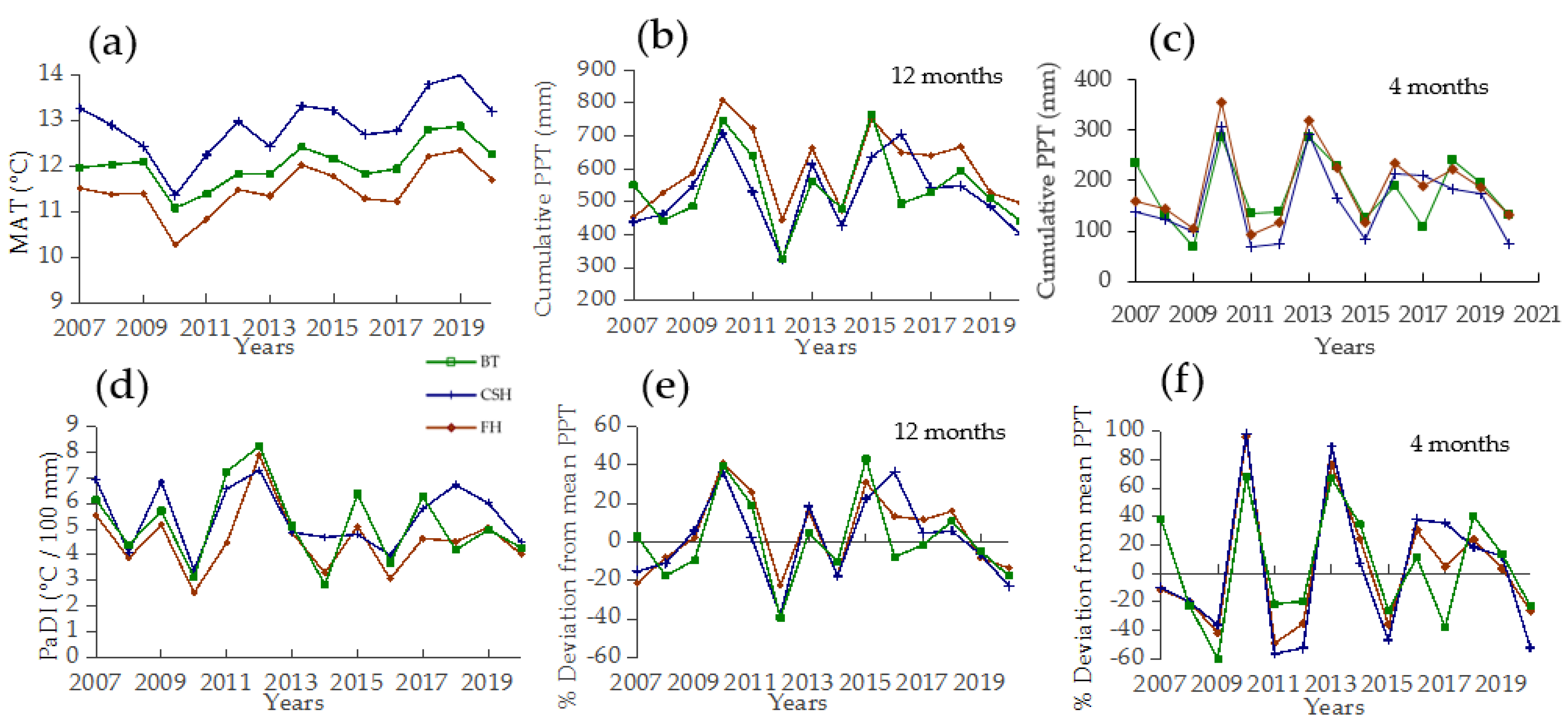
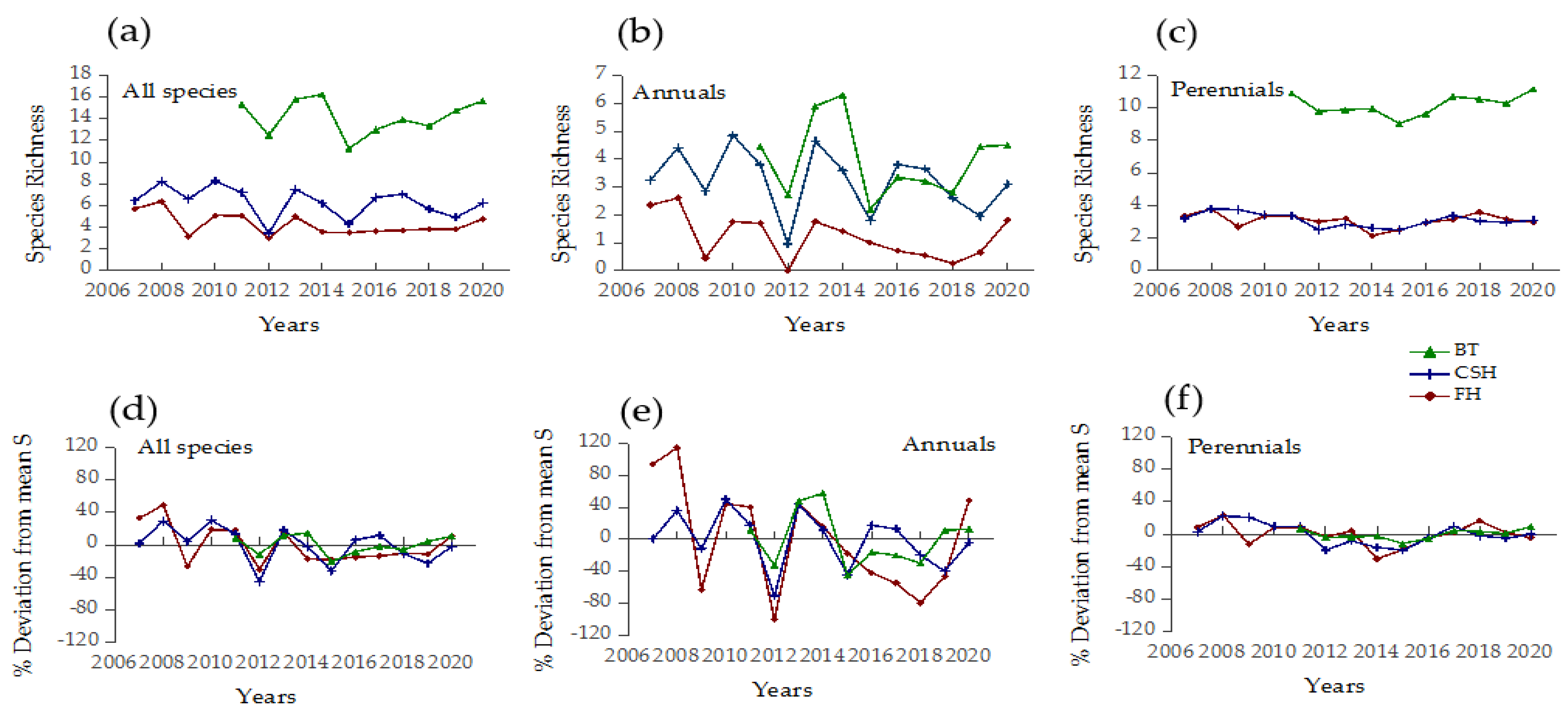
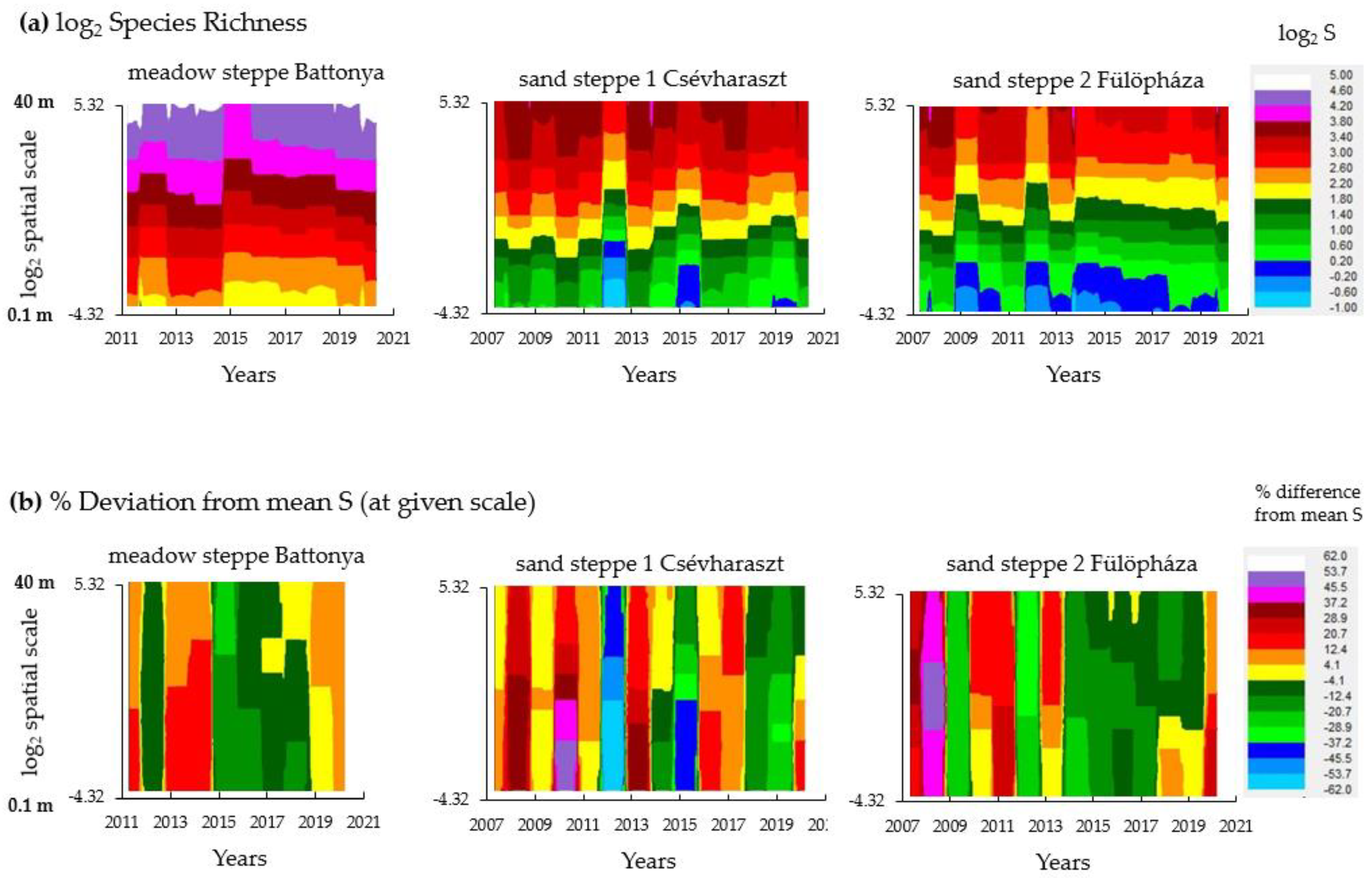
3.1. Weather Fluctuations and Temporal Diversity Patterns
3.2. Spatial Synchrony among Sites
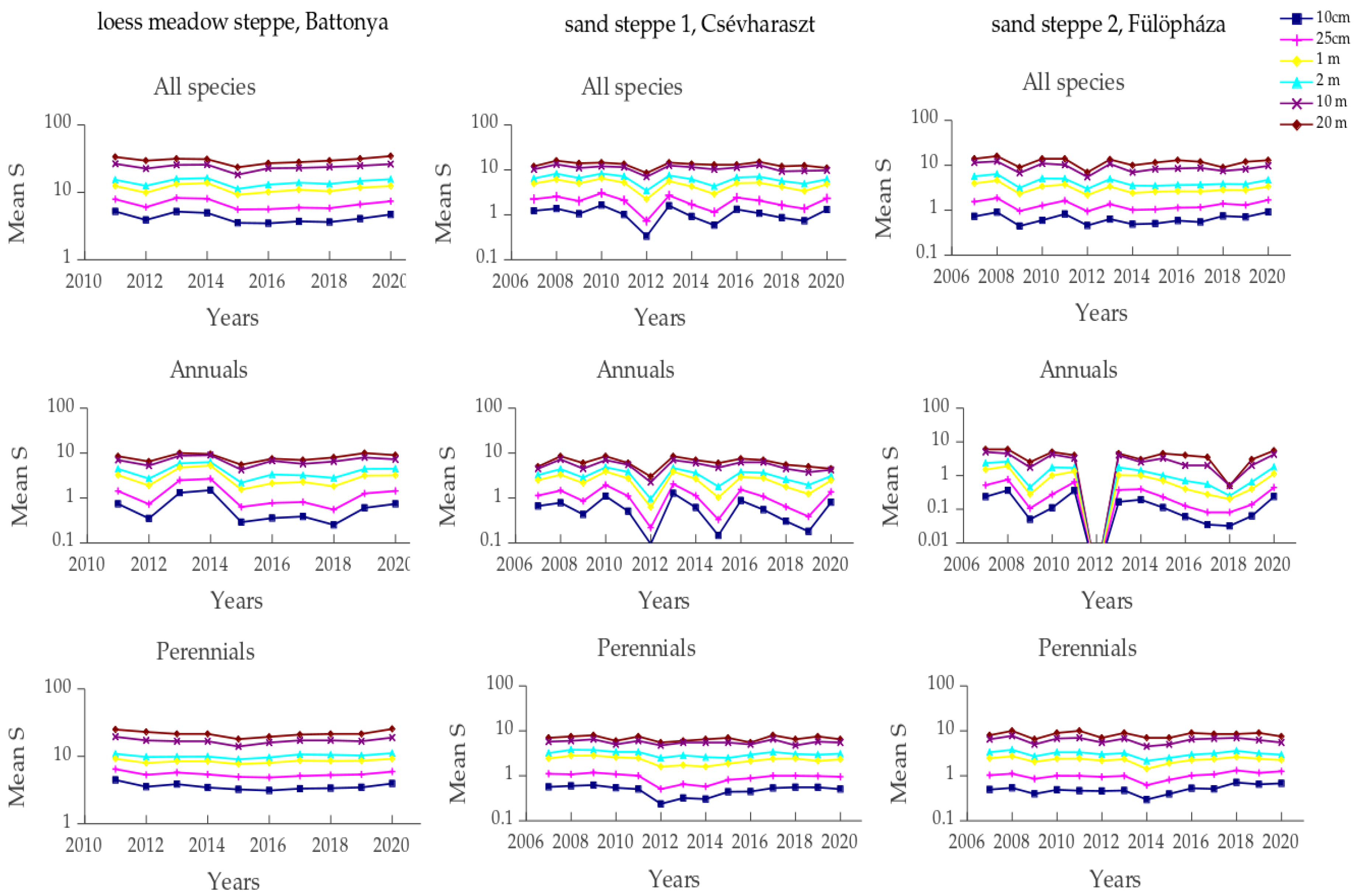
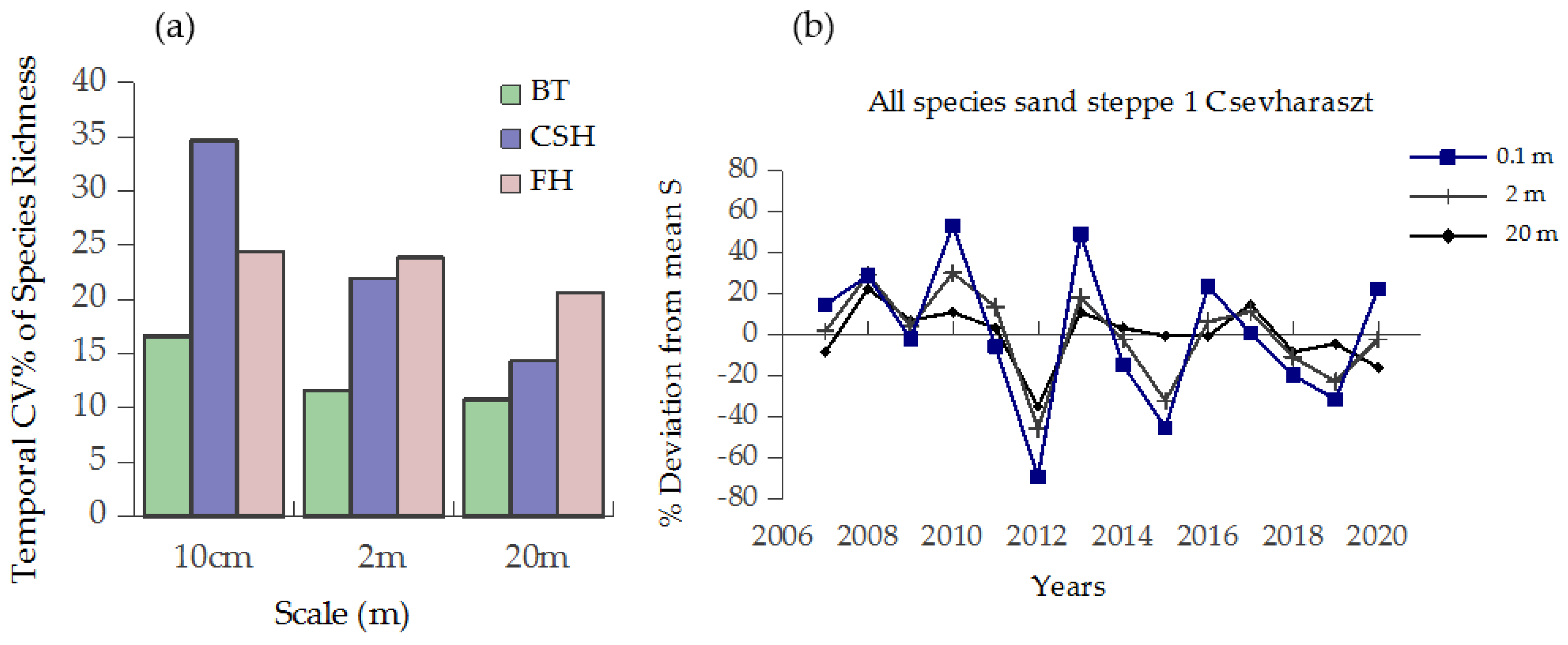
3.3. Synchrony of Species Richness Within-Site over Different Spatial Scales
3.4. Relationships between Weather Fluctuations and Diversity Patterns
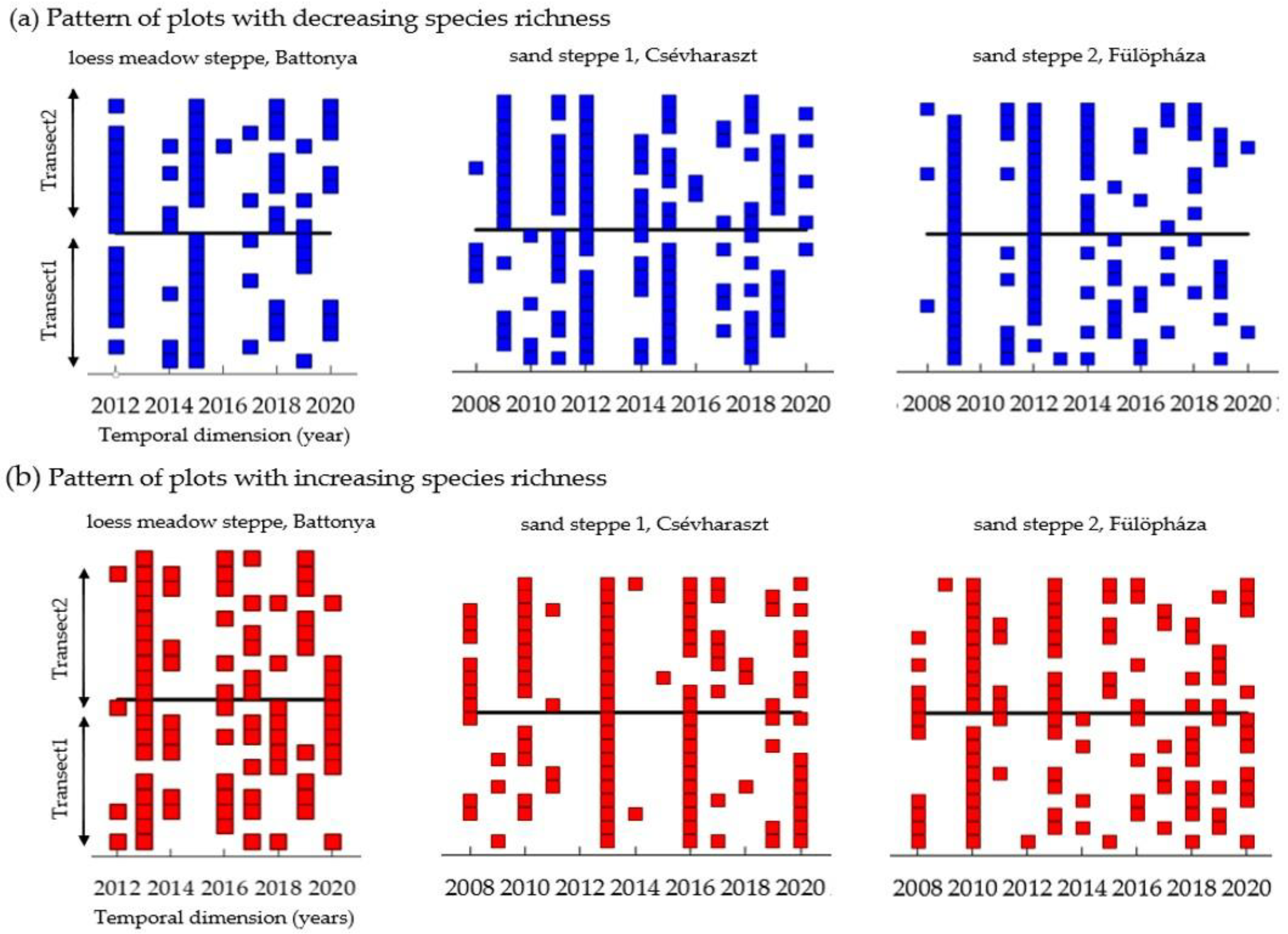
3.5. Micro-Scale Spatial Synchrony of Inter-Annual Diversity Changes
4. Discussion
4.1. Diversity Patterns Indicate Stable Vegetation despite Extreme Weather Fluctuations
4.2. Scale Dependence of Diversity Responses
4.3. Synchrony between Weather Fluctuations and Diversity Changes
4.4. Climatic Drivers and Vegetation Responses
4.5. Implication for Future Monitoring
4.6. Limitations and Needs for Future Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blair, J.; Nippert, J.; Briggs, J. Grassland Ecology. In Ecology and the Environment; The Plant Sciences 8, Chapter 14; Monson, R.K., Ed.; Springer: New York, NY, USA, 2014; pp. 389–423. [Google Scholar] [CrossRef]
- Chesson, P.; Gebauer, R.L.E.; Schwinning, S.; Huntly, N.; Wiegand, K.; Ernest, M.S.K.; Sher, A.; Novoplansky, A.; Weltzin, J.F. Resource pulses, species interactions, and diversity maintenance in arid and semi-arid environments. Oecologia 2004, 141, 236–253. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.L.; Chung, Y.A.; Baur, L.E.; Hallmark, A.; Ohlert, T.J.; Rudgers, J.A. Press–pulse interactions and long-term community dynamics in a Chihuahuan Desert grassland. J. Veg. Sci. 2020, 31, 722–732. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change 2013; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; p. 1535. Available online: https://www.ipcc.ch/site/assets/uploads/2018/02/WG1AR5_all_final.pdf (accessed on 16 December 2021).
- Magurran, A.E.; Baillie, S.R.; Buckland, S.T.; Dick, J.M.P.; Elston, D.A.; Scott, E.M.; Smith, R.I.; Somerfield, P.J.; Watt, A.D. Long-term datasets in biodiversity research and monitoring: Assessing change in ecological communities through time. Trends Ecol. Evol. 2010, 25, 574–582. [Google Scholar] [CrossRef] [PubMed]
- De Bello, F.; Valencia, E.; Ward, D.; Hallett, L. Why we still need permanent plots for vegetation science. J. Veg. Sci. 2020, 31, 679–685. [Google Scholar] [CrossRef]
- Adler, P.B.; Levine, J.M. Contrasting relationships between precipitation and species richness in space and time. Oikos 2007, 116, 221–232. [Google Scholar] [CrossRef]
- Cleland, E.E.; Collins, S.L.; Dickson, T.L.; Farrer, E.C.; Gross, K.L.; Gherardi, L.A.; Hallett, L.M.; Hobbs, R.J.; Hsu, J.S.; Turnbull, L.; et al. Sensitivity of grassland plant community composition to spatial vs. temporal variation in precipitation. Ecology 2013, 94, 1687–1696. [Google Scholar] [CrossRef]
- Harrison, S.P.; Gornish, E.S.; Copeland, S. Climate-driven diversity loss in a grassland community. Proc. Natl. Acad. Sci. USA 2015, 112, 8672–8677. [Google Scholar] [CrossRef]
- Jonas, J.L.; Buhl, D.A.; Symstad, A.J. Impacts of weather on long-term patterns of plant richness and diversity vary with location and management. Ecology 2015, 96, 2417–2432. [Google Scholar] [CrossRef]
- Wilcox, K.R.; Shi, Z.; Gherardi, L.A.; Lemoine, N.P.; Koerner, S.E.; Hoover, D.L.; Bork, E.; Byrne, K.M.; Cahill, J., Jr.; Collins, S.L.; et al. Asymmetric responses of primary productivity to precipitation extremes: A synthesis of grassland precipitation manipulation experiments. Glob. Chang. Biol. Bioenergy 2017, 23, 4376–4385. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, L.; Tang, Z.; Wu, S. A synthesis of ecosystem aboveground productivity and its process variables under simulated drought stress. J. Ecol. 2019, 107, 2519–2531. [Google Scholar] [CrossRef]
- Zhang, F.; Quan, Q.; Ma, F.; Tian, D.; Hoover, D.L.; Zhou, Q.; Niu, S. When does extreme drought elicit extreme ecological responses? J. Ecol. 2019, 107, 2553–2563. [Google Scholar] [CrossRef]
- Gruner, D.S.; Bracken, M.E.S.; Berger, S.A.; Eriksson, B.K.; Gamfeldt, L.; Matthiessen, B.; Moorthi, S.; Sommer, U.; Hillebrand, H. Effects of experimental warming on biodiversity depend on ecosystem type and local species composition. Oikos 2017, 126, 8–17. [Google Scholar] [CrossRef]
- Piseddu, F.; Bellocchi, G.; Picon-Cochard, C. Mowing and warming effects on grassland species richness and harvested biomass: Meta-analyses. Agron. Sustain. Dev. 2021, 41, 74. [Google Scholar] [CrossRef]
- Korell, L.; Auge, H.; Chase, J.M.; Harpole, W.S.; Knight, T.M. Responses of plant diversity to precipitation change are strongest at local spatial scales and in drylands. Nat. Commun. 2021, 12, 2489. [Google Scholar] [CrossRef]
- Morris, W.F.; Pfister, C.A.; Tuljapurkar, S.; Haridas, C.V.; Boggs, C.L.; Boyce, M.S.; Bruna, E.M.; Church, D.R.; Coulson, T.; Doak, D.F.; et al. Longevity can buffer plant and animal populations against changing climatic variability. Ecology 2008, 89, 19–25. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Vicca, S.; Roy, J.; Nijs, I.; Milcu, A.; Kreyling, J.; Jentsch, A.; Chabbi, A.; Campioli, M.; Callaghan, T.; et al. Global change experiments: Challenges and opportunities. BioScience 2015, 65, 922–931. [Google Scholar] [CrossRef]
- Leuzinger, S.; Luo, Y.; Beier, C.; Dieleman, W.; Vicca, S.; Körner, C. Do global change experiments overestimate impacts on terrestrial ecosystems? Trends Ecol. Evol. 2011, 26, 236–241. [Google Scholar] [CrossRef]
- Arribas, L.P.; Gutiérrez, J.L.; Bagur, M.; Soria, S.A.; Penchaszadeh, P.E.; Palomo, M.G. Variation in aggregate descriptors of rocky shore communities: A test of synchrony across spatial scales. Mar. Biol. 2019, 166, 44. [Google Scholar] [CrossRef]
- Chase, J.M.; Knight, T.M. Scale-dependent effect sizes of ecological drivers on biodiversity: Why standardised sampling is not enough. Ecol. Lett. 2013, 16, 17–26. [Google Scholar] [CrossRef]
- Podani, J. Analysis of mapped and simulated vegetation patterns by means of computerized sampling techniques. Acta Bot. Hung. 1984, 30, 403–425. [Google Scholar]
- Bartha, S.; Campatella, G.; Canullo, R.; Bódis, J.; Mucina, L. On the importance of fine-scale spatial complexity in vegetation restoration. Int. J. Ecol. Environ. Sci. 2004, 30, 101–116. Available online: https://ecolres.hu/sites/default/files/IJEES2004.pdf (accessed on 16 December 2021).
- Buonaccorsi, J.P.; Elkinton, J.S.; Evans, S.R.; Liebhold, A.M. Measuring and testing for spatial synchrony. Ecology 2001, 82, 1668–1679. [Google Scholar] [CrossRef]
- Liebhold, A.; Koenig, W.D.; Bjornstad, O.N. Spatial synchrony in population dynamics. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 467–490. [Google Scholar] [CrossRef]
- Walter, J.A.; Shoemaker, L.G.; Lany, N.K.; Castorani, M.C.N.; Fey, S.B.; Dudney, J.C.; Gherardi, L.; Portales-Reyes, C.; Rypel, A.L.; Cottingham, K.L.; et al. The spatial synchrony of species richness and its relationship to ecosystem stability. Ecology 2021, 102, e03486. [Google Scholar] [CrossRef]
- Li, Z.; Ma, W.; Liang, C.; Liu, Z.; Wang, W.; Wang, L. Long-term vegetation dynamics driven by climatic variations in the Inner Mongolia grassland: Findings from 30-year monitoring. Landsc. Ecol. 2015, 30, 1701–1711. [Google Scholar] [CrossRef]
- Aronson, J.; Shmida, A. Plant species diversity along a Mediterranean-desert gradient and its correlation with interannual rainfall fluctuations. J. Arid Environ. 1992, 23, 235–247. [Google Scholar] [CrossRef]
- Chelli, S.; Simonetti, E.; Campetella, G.; Chiarucci, A.; Cervellini, M.; Tardella, F.M.; Tomasella, M.; Canullo, R. Plant diversity changes in a nature reserve: A probabilistic sampling method for quantitative assessments. Nat. Conserv. 2019, 34, 145–161. [Google Scholar] [CrossRef]
- Mezősi, G.; Bata, T.; Meyer, B.C.; Blanka, V.; Ladányi, Z. Climate change impacts on environmental hazards on the Great Hungarian Plain, Carpathian Basin. Int. J. Disaster Risk Sci. 2014, 5, 136–146. [Google Scholar] [CrossRef][Green Version]
- Molnár, Z.; Biró, M.; Bartha, S.; Fekete, G. Past trends, present state and future prospects of Hungarian forest-steppes. In Eurasian Steppes. Ecological Problems and Livelihoods in a Changing World; Werger, M., Ed.; Springer: Dordrecht, The Netherlands, 2012; Chapter 7; pp. 209–252. [Google Scholar]
- Csathó, A.J.; Csathó, A.I. The flora list of the Külső-gulya meadow of Battonya-Tompapuszta (SE Hungary). Crisicum 2009, 5, 51–70. Available online: https://docplayer.hu/20471202-A-battonya-tompapusztai-kulso-gulya-floralistaja.html (accessed on 16 December 2021).
- Kovács-Láng, E.; Kröel-Dulay, G.; Kertész, M.; Fekete, G.; Bartha, S.; Mika, J.; Dobi-Wantuch, I.; Rédei, T.; Rajkai, K.; Hahn, I. Changes in the composition of sand grasslands along a climatic gradient in Hungary and implications for climate change. Phytocoenologia 2000, 30, 385–407. [Google Scholar] [CrossRef]
- Virágh, K.; Horváth, A.; Bartha, S.; Somodi, I. A multiscale methodological approach for monitoring the effectiveness of grassland management. Community Ecol. 2008, 9, 237–246. [Google Scholar] [CrossRef]
- Pálfai, I. Description and forecasting of droughts in Hungary. In Proceedings of the 14th Congress on Irrigation and Drainage (ICID), Rio de Janeiro, Brazil, 30 April–4 May 1990; Volume 1-C, pp. 151–158. [Google Scholar]
- Pálfai, I. Probability of drought occurence in Hungary. Q. J. Hung. Meteorol. Serv. 2002, 106, 265–275. [Google Scholar]
- Kozák, P.; Pálfai, I.; Herceg, Á. Palfai Drought Index (PaDI)—The extension of the adaptability of Pálfai Index (PAI) for South East Europe. In DMCSEE—Summary of the Project Results; Bihari, Z., Ed.; Hungarian Meterological Service: Budapest, Hungary, 2012. (In Hungarian) [Google Scholar]
- R Core Team. R: A language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2020. Available online: https://www.R-project.org/ (accessed on 12 December 2020).
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: Berlin/Heidelberg, Germany, 2000; pp. 235–397. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Core Team_nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3. 2020, pp. 1–144. Available online: https://CRAN.R-project.org/package=nlme (accessed on 12 December 2020).
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version 1.43.17. 2020. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 13 December 2020).
- Juhász-Nagy, P.; Podani, J. Information theory methods for the study of spatial processes and succession. Vegetatio 1983, 51, 129–140. [Google Scholar] [CrossRef]
- Juhász-Nagy, P. Spatial dependence of plant populations. Part 2. A family of new models. Acta Bot. Acad. Sci. Hung. 1984, 30, 363–402. [Google Scholar]
- Tsakalos, J.L. An R Package to Analyze within-Community Spatial Organization Using Species Combinations. GitHub Repository. 2022. Available online: https://github.com/jamestsakalos/comspat (accessed on 16 December 2021).
- Tsakalos, J.L.; Chelli, S.; Campetella, G.; Canullo, R.; Simonetti, E.; Bartha, S. An R package to analyze within-community spatial organization using species combinations. Ecography 2022. Manuscript Resubmitted after Minor Revision. [Google Scholar]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. Available online: https://www.jstor.org/stable/4615733 (accessed on 16 December 2021).
- Tilman, D.; El Haddi, A. Drought and biodiversity in grasslands. Oecologia 1992, 89, 257–264. [Google Scholar] [CrossRef]
- Doležal, J.; Altman, J.; Jandová, V.; Chytrý, M.; Conti, L.; Méndez-Castro, F.E.; Klimešová, J.; Zelený, D.; Ottaviani, G. Climate warming and extended droughts drive establishment and growth dynamics in temperate grassland plants. Agric. For. Meteorol. 2022, 313, 108762. [Google Scholar] [CrossRef]
- Jentsch, A.; Kreyling, J.; Beierkuhnlein, C. A new generation of climate-change experiments: Events, not trends. Front. Ecol. Environ. 2007, 5, 365–374. [Google Scholar] [CrossRef]
- Smith, M.D. An ecological perspective on extreme climatic events: A synthetic definition and framework to guide future research. J. Ecol. 2011, 99, 656–663. [Google Scholar] [CrossRef]
- Jentsch, A.; Kreyling, J.; Elmer, M.; Gellesch, E.; Glaser, B.; Grant, K.; Hein, R.; Lara, M.; Mirzae, H.; Nadler, S.E.; et al. Climate extremes initiate ecosystem-regulating functions while maintaining productivity. J. Ecol. 2011, 99, 689–702. [Google Scholar] [CrossRef]
- Matesanz, S.; Brooker, R.W.; Valladares, F.; Klotz, S. Temporal dynamics of marginal steppic vegetation over a 26-year period of substantial environmental change. J. Veg. Sci. 2009, 20, 299–310. [Google Scholar] [CrossRef]
- Jones, S.K.; Ripplinger, J.; Collins, S.L. Species reordering, not changes in richness, drives long-term dynamics in grassland communities. Ecol. Lett. 2017, 20, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J.; Shimadzu, H.; Dornelas, M.; McGill, B.; Moyes, F.; Magurran, A.E. Community-level regulation of temporal trends in biodiversity. Sci. Adv. 2017, 3, e1700315. [Google Scholar] [CrossRef]
- Kröel-Dulay, G.; Ransijn, J.; Schmidt, I.K.; Beier, C.; De Angelis, P.; De Dato, G.; Dukes, J.S.; Emmett, B.; Estiarte, M.; Garadnai, J.; et al. Increased sensitivity to climate change in disturbed ecosystems. Nat. Commun. 2015, 6, 6682. [Google Scholar] [CrossRef]
- Bartha, S.; Horváth, F. Application of long transects and information theoretical functions to pattern detection. I. Transects versus isodiametric sampling units. Abstr. Bot. 1987, 11, 9–26. Available online: https://www.jstor.org/stable/43519147 (accessed on 16 December 2021).
- Dengler, J.; Matthews, T.J.; Steinbauer, M.J.; Wolfrum, S.; Boch, S.; Chiarucci, A.; Conradi, T.; Dembicz, I.; Marcenò, C.; García-Mijangos, I.; et al. Species–area relationships in continuous vegetation: Evidence from Palaearctic grasslands. J. Biogeogr. 2020, 47, 72–86. [Google Scholar] [CrossRef]
- Zhang, J.; Gillet, F.; Bartha, S.; Alato, J.M.; Biurrun, I.; Dembicz, I.; Grytnes, J.-A.; Jaunatre, R.; Pielech, R.; Van Meerbeek, K.; et al. Scale dependence of species–area relationships is widespread but generally weak in Palaearctic grasslands. J. Veg. Sci. 2021, 32, e13044. [Google Scholar] [CrossRef]
- Dembicz, I.; Dengler, J.; Steinbauer, M.J.; Matthews, T.J.; Bartha, S.; Burrascano, S.; Chiarucci, A.; Filibeck, G.; Gillet, F.; Janišová, M.; et al. Fine-grain beta diversity of Palaearctic grassland vegetation. J. Veg. Sci. 2021, 32, e13045. [Google Scholar] [CrossRef]
- Güler, B.; Jentsch, A.; Apostolova, I.; Bartha, S.; Bloor, J.M.; Campetella, G.; Canullo, R.; Házi, J.; Kreyling, J.; Pottier, J.; et al. How plot shape and dispersion affect plant species richness counts: Implications for sampling design and rarefaction analyses. J. Veg. Sci. 2016, 27, 692–703. [Google Scholar] [CrossRef]
- Bartha, S.; Meiners, S.J.; Pickett, S.T.A.; Cadenasso, M.L. Plant colonization windows in a mesic old field succession. Appl. Veg. Sci. 2003, 6, 205–212. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Koncz, P.; Vadász-Besnyői, V.; Csathó, A.I.; Nagy, J.; Szerdahelyi, T.; Tóth, Z.; Pintér, K.; Fóti, S.; Papp, M.; Balogh, J.; et al. Carbon uptake changed but vegetation composition remained stable during transition from grazing to mowing grassland management. Agric. Ecosyst. Environ. 2020, 304, 107161. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Likens, G.E. Adaptive monitoring: A new paradigm for long-term research and monitoring. Trends Ecol. Evol. 2009, 24, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, B.; Collins, S.L. Multiscale monitoring of a multispecies case study: Two grass species at Sevilleta. Plant Ecol. 2005, 179, 149–154. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Observed Synchrony | Null Model Synchrony | p-Value |
|---|---|---|---|
| Mean annual temperature | 0.790 | 0.509 | 0.000 |
| Pálfai aridity index | 0.930 | 0.503 | 0.000 |
| Precipitation 4 months | 0.860 | 0.505 | 0.000 |
| Precipitation 12 months | 0.789 | 0.500 | 0.000 |
| Spatial Scale | Characteristics | Observed Synchrony | Null Model Synchrony | p-Value | Adjusted p-Value |
|---|---|---|---|---|---|
| 0.1 m | All species | 0.630 | 0.498 | 0.181 | 1.000 |
| Annuals | 0.704 | 0.489 | 0.049 | 0.441 | |
| Perennials | 0.630 | 0.489 | 0.164 | 1.000 | |
| 2 m | All species | 0.778 | 0.495 | 0.012 | 0.108 |
| Annuals | 0.778 | 0.486 | 0.013 | 0.117 | |
| Perennials | 0.704 | 0.475 | 0.046 | 0.419 | |
| 40 m | All species | 0.444 | 0.409 | 0.415 | 1.000 |
| Annuals | 0.519 | 0.392 | 0.160 | 1.000 | |
| Perennials | 0.296 | 0.359 | 0.751 | 1.000 |
| Sites | Characteristics | Observed Synchrony | Null Model Synchrony | p-Value | Adjusted p-Value |
|---|---|---|---|---|---|
| Battonya | All species | 0.837 | 0.497 | 0.000 | 0.000 |
| meadow | Annuals | 0.778 | 0.488 | 0.000 | 0.000 |
| steppe | Perennials | 0.622 | 0.454 | 0.007 | 0.007 |
| Csévharaszt | All species | 0.867 | 0.489 | 0.000 | 0.000 |
| sand | Annuals | 0.949 | 0.476 | 0.000 | 0.000 |
| steppe1 | Perennials | 0.621 | 0.446 | 0.004 | 0.008 |
| Fülöpháza | All species | 0.790 | 0.486 | 0.000 | 0.000 |
| sand | Annuals | 0.774 | 0.473 | 0.000 | 0.000 |
| steppe2 | Perennials | 0.785 | 0.454 | 0.000 | 0.000 |
| Species Richness Type | Weather Characteristic | Observed Association | Null Model Association | p-Value | p after Correction |
|---|---|---|---|---|---|
| All species | Precipitation 4 months | 2.467 | 1.515 | 0.0000 | 0.0000 |
| Precipitation 12 months | 2.082 | 1.308 | 0.0010 | 0.0040 | |
| Pálfai aridity index | 1.680 | 1.223 | 0.0190 | 0.0380 | |
| Annuals | Precipitation 4 months | 2.625 | 1.629 | 0.0000 | 0.0000 |
| Precipitation 12 months | 1.944 | 1.390 | 0.0070 | 0.0210 | |
| Pálfai aridity index | 2.133 | 1.319 | 0.0000 | 0.0000 | |
| Perennials | Precipitation 4 months | 2.348 | 1.277 | 0.0000 | 0.0000 |
| Precipitation 12 months | 1.964 | 1.109 | 0.0000 | 0.0000 | |
| Pálfai aridity index | 1.393 | 1.036 | 0.0551 | 0.0551 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartha, S.; Szabó, G.; Csete, S.; Purger, D.; Házi, J.; Csathó, A.I.; Campetella, G.; Canullo, R.; Chelli, S.; Tsakalos, J.L.; et al. High-Resolution Transect Sampling and Multiple Scale Diversity Analyses for Evaluating Grassland Resilience to Climatic Extremes. Land 2022, 11, 378. https://doi.org/10.3390/land11030378
Bartha S, Szabó G, Csete S, Purger D, Házi J, Csathó AI, Campetella G, Canullo R, Chelli S, Tsakalos JL, et al. High-Resolution Transect Sampling and Multiple Scale Diversity Analyses for Evaluating Grassland Resilience to Climatic Extremes. Land. 2022; 11(3):378. https://doi.org/10.3390/land11030378
Chicago/Turabian StyleBartha, Sándor, Gábor Szabó, Sándor Csete, Dragica Purger, Judit Házi, András István Csathó, Giandiego Campetella, Roberto Canullo, Stefano Chelli, James Lee Tsakalos, and et al. 2022. "High-Resolution Transect Sampling and Multiple Scale Diversity Analyses for Evaluating Grassland Resilience to Climatic Extremes" Land 11, no. 3: 378. https://doi.org/10.3390/land11030378
APA StyleBartha, S., Szabó, G., Csete, S., Purger, D., Házi, J., Csathó, A. I., Campetella, G., Canullo, R., Chelli, S., Tsakalos, J. L., Ónodi, G., Kröel-Dulay, G., & Zimmermann, Z. (2022). High-Resolution Transect Sampling and Multiple Scale Diversity Analyses for Evaluating Grassland Resilience to Climatic Extremes. Land, 11(3), 378. https://doi.org/10.3390/land11030378