
Estimating Fragmentation and Connectivity Patterns of the Temperate Forest in an Avocado-Dominated Landscape to Propose Conservation Strategies
 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
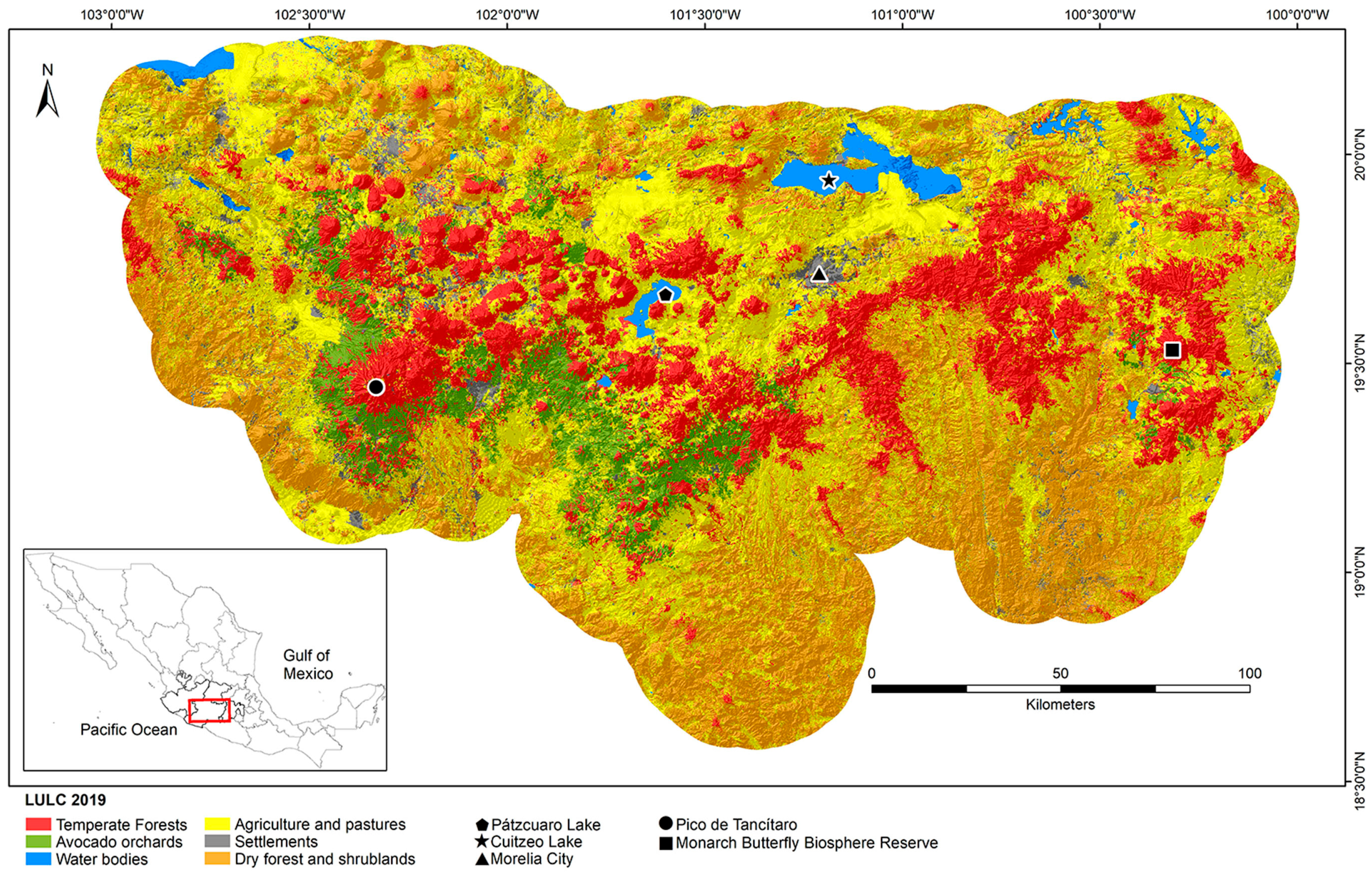
2.1. Land Use and Land Cover Classification and Accuracy
2.2. Fragmentation and Connectivity of the Temperate Forest
3. Results
3.1. Land Use and Land Cover Classification and Accuracy
3.2. Fragmentation and Connectivity of the Temperate Forest
4. Discussion
Implication for Conservation
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monson, R.K. Ecology of Temperate Forests. In Ecology and the Environment; Monson, R., Ed.; The Plant Sciences; Springer: New York, NY, USA, 2014. [Google Scholar]
- de Gouvenain, R.C.; Silander, J.A. Temperate Forests. Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2017; Available online: https://www.sciencedirect.com/science/article/pii/B9780128096338023104 (accessed on 27 July 2022).
- Instituto Nacional de Estadística y Geografía—INEGI. Datos Vectoriales de uso del Suelo y Vegetación: Serie V. 2015. Available online: https://www.inegi.org.mx/temas/usosuelo/#Mapa (accessed on 15 March 2022).
- Rzedowski, J. Diversidad y orígenes de la flora fanerogámica de México. Acta Bot. Mex. 1991, 14, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Galicia, L.; Zarco-Arista, A.E. Multiple ecosystem services, possible trade-offs and synergies in a temperate forest ecosystem in Mexico: A review. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2014, 10, 275–288. [Google Scholar] [CrossRef]
- Galicia, L.; Potvin, C.; Messier, C. Maintaining the high diversity of pine and oak species in Mexican temperate forests: A new management approach combining functional zoning and ecosystem adaptability. Can. J. For. Res. 2015, 45, 1358–1368. [Google Scholar] [CrossRef] [Green Version]
- Instituto Nacional de Estadística y Geografía—INEGI. Anuario Estadístico de la Producción Forestal 2016. Secretaría de Medio Ambiente y Recursos Naturales, México. 2016. Available online: https://www.gob.mx/semarnat/documentos/anuarios-estadisticos-forestales (accessed on 1 February 2022).
- Torres-Rojo, J.M.; Guevara, S.A. El potencial de México para la producción de servicios ambientales: Captura de carbono y desempeño hidráulico. Gac. Ecológica 2002, 63, 40–59. [Google Scholar]
- Ordóñez, J.A.B.; de Jong, B.H.J.; García-Oliva, F.; Aviña, F.L.; Pérez, J.V.; Guerrero, G.; Martínez, R.; Masera, O. Carbon content in vegetation, litter, and soil under 10 different land-use and land-cover classes in the Central Highlands of Michoacán, Mexico For. Ecol. Manag. 2008, 255, 2074–2084. [Google Scholar] [CrossRef]
- Cavender-Bares, J. Diversity, distribution and ecosystem services of the North American oaks. Int. Oaks 2016, 27, 37–48. [Google Scholar]
- Farjon, A. Biodiversity of Pinus (Pinaceae) in Mexico: Speciation and palaeo-endemism. Bot. J. Linn. Soc. 1996, 121, 365–384. [Google Scholar] [CrossRef]
- Hipp, A.L.; Manos, P.S.; González-Rodríguez, A.; Hahn, M.; Kaproth, M.; McVay, J.D.; Cavender-Bares, J. Sympatric parallel diversification of major oak clades in the Americas and the origins of Mexican species diversity. New Phytol. 2018, 217, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Masera, O.; Ordoñez, M.J.; Dirzo, R. Carbon emissions from Mexican forest current situation and long term scenarios. Clim. Change 1997, 35, 265–295. [Google Scholar] [CrossRef]
- Valencia, A.S. Diversidad del género Quercus (Fagaceae) en México. Bol. Soc. Bot. México 2004, 75, 33–53. [Google Scholar] [CrossRef] [Green Version]
- Bocco, G.; Mendoza, M.; Masera, O.R. La dinámica del cambio del uso del suelo en Michoacán: Una propuesta metodológica para el estudio de los procesos de deforestación. Investig. Geográficas 2001, 44, 18–36. [Google Scholar] [CrossRef]
- Mas, J.F.; Lemoine-Rodríguez, R.; González, R.; López-Sánchez, J.; Piña-Garduño, A.; Herrera-Flores, E. Evaluación de las tasas de deforestación en Michoacán a escala detallada mediante un método híbrido de clasificación de imágenes SPOT. Madera Bosques 2017, 23, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Loya-Carrillo, J.O.; Mass Caussel, J.F. Analysis of deforestation in Michoacán state: From the spatial to social. Rev. Cart. 2020, 101, 99–117. [Google Scholar]
- Molina Sánchez, A.; Delgado, P.; González-Rodríguez, A.; González, C.; Gómez-Tagle Rojas, A.; Lopez-Toledo, L. Spatio-temporal approach for identification of critical conservation areas: A case study with two pine species from a threatened temperate forest in Mexico. Biodivers. Conserv. 2019, 28, 1863–1883. [Google Scholar] [CrossRef]
- Marroquín-Páramo, J.A.; Suazo-Ortuño, I.; Mendoza, E.; Alvarado-Díaz, J.; Siliceo-Cantero, H.H. Herpetofaunal diversity in avocado orchards and in conserved habitats in Michoacán, Mexico. Rev. Mex. Biodivers. 2017, 88, 234–240. [Google Scholar] [CrossRef]
- Villamil, L.; Astier, M.; Merlín, Y.; Ayala-Barajas, R.; Ramírez-García, E.; Martínez-Cruz, J.; Gavito, M.E. Management practices and diversity of flower visitors and herbaceous plants in conventional and organic avocado orchards in Michoacán, Mexico. Agroecol. Sustain. Food Syst. 2018, 42, 530–551. [Google Scholar] [CrossRef]
- Sáenz-Ceja, J.E.; Pérez-Salicrup, D.R. Avocado Cover Expansion in the Monarch Butterfly Biosphere Reserve, Central Mexico. Conservation 2021, 1, 299–310. [Google Scholar] [CrossRef]
- Bravo-Espinosa, M.; Mendoza, M.E.; Carlón Allende, T.; Medina, L.; Sáenz-Reyes, J.T.; Páez, R. Effects of converting forest to avocado orchards on topsoil properties in the trans-Mexican volcanic system, Mexico. Land Degrad. Dev. 2014, 25, 452–467. [Google Scholar] [CrossRef]
- Cho, K.; Goldstein, B.; Gounaridis, D.; Newell, J.P. Where does your guacamole come from? Detecting deforestation associated with the export of avocados from Mexico to the United States. J. Environ. Manag. 2021, 278, 111482. [Google Scholar] [CrossRef]
- Denvir, A.; Arima, E.Y.; González-Rodríguez, A.; Young, K.R. Ecological and human dimensions of avocado expansion in México: Towards supply-chain sustainability. Ambio 2021, 51, 152–166. [Google Scholar] [CrossRef]
- Molina-Sánchez, A. Fragmentación del Hábitat y su Efecto en la Estructura Genética de dos Linajes del Género Pinus Distribuidos en la Meseta Purépecha. Master’s Thesis, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Mexico, 2017. Available online: http://bibliotecavirtual.dgb.umich.mx:8083/xmlui/handle/DGB_UMICH/1359 (accessed on 1 February 2022).
- Servicio de Información Agroalimentaria y Pesquera—SIAP. Avance de Siembras y Cosechas. Aguacate, Michoacán. 2022. Available online: https://nube.siap.gob.mx/avance_agricola/ (accessed on 1 October 2022).
- Arima, E.Y.; Denvir, A.; Young, K.R.; González-Rodríguez, A.; García-Oliva, F. Modelling avocado-driven deforestation in Michoacán, Mexico. Environ. Res. Lett. 2022, 17, 034015. [Google Scholar] [CrossRef]
- Auffret, A.G.; Plue, J. Scale-dependent diversity effects of seed dispersal by a wild herbivore in fragmented grasslands. Oecologia 2014, 175, 305–313. [Google Scholar] [CrossRef]
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity is a vital element of landscape structure. Oikos 1993, 68, 571–573. [Google Scholar] [CrossRef] [Green Version]
- With, K.A.; Gardner, R.H.; Turner, M.G. Landscape connectivity and population distributions in heterogeneous environments. Oikos 1997, 78, 151–169. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Uroy, L.; Ernoult, A.; Mony, C. Effect of landscape connectivity on plant communities: A review of response patterns. Landsc. Ecol. 2019, 34, 203–225. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Townshend, J.R. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.D.; Fahrig, L.; With, K.A. Landscape connectivity: A return to the basics. In Connectivity Conservation; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Keeley, A.T.; Beier, P.; Jenness, J.S. Connectivity metrics for conservation planning and monitoring. Biol. Conserv. 2021, 255, 109008. [Google Scholar] [CrossRef]
- Minor, E.S.; Urban, D.L. Graph theory as a proxy for spatially explicit population models in conservation planning. Ecol. Appl. 2007, 17, 1771–1782. [Google Scholar] [CrossRef]
- Pascual-Hortal, L.; Saura, S. Comparison and development of new graph-based landscape connectivity indices: Towards the priorization of habitat patches and corridors for conservation. Landsc. Ecol. 2006, 21, 959–967. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. 2012. Available online: https://www.fs.usda.gov/pnw/pubs/pnw_gtr351.pdf (accessed on 12 February 2023).
- Auffret, A.G.; Rico, Y.; Bullock, J.M.; Hooftman, D.A.; Pakeman, R.J.; Soons, M.B.; Cousins, S.A. Plant functional connectivity–integrating landscape structure and effective dispersal. J. Ecol. 2017, 105, 1648–1656. [Google Scholar] [CrossRef] [Green Version]
- Griffen, B.D.; Drake, J.M. Effects of habitat quality and size on extinction in experimental populations. Proc. R. Soc. B Biol. Sci. 2008, 275, 2251–2256. [Google Scholar] [CrossRef] [Green Version]
- Grace, S.L.; Hamrick, J.L.; Platt, W.J. Estimation of seed dispersal in an old-growth population of longleaf pine (Pinus palustris) using maternity exclusion analysis. Castanea 2004, 69, 207–215. [Google Scholar] [CrossRef]
- López-Barrera, F.; Manson, R.H.; González-Espinosa, M.; Newton, A.C. Effects of varying forest edge permeability on seed dispersal in a neotropical montane forest. Landsc. Ecol. 2007, 22, 189–203. [Google Scholar] [CrossRef]
- Axer, M.; Schlicht, R.; Wagner, S. Modelling potential density of natural regeneration of European oak species (Quercus robur L., Quercus petraea (Matt.) Liebl.) depending on the distance to the potential seed source: Methodological approach for modelling dispersal from inventory data at forest enterprise level. For. Ecol. Manag. 2021, 482, 118802. [Google Scholar]
- Ashley, M.V. Answers Blowing in the Wind: A Quarter Century of Genetic Studies of Pollination in Oaks. Forests 2021, 12, 575. [Google Scholar] [CrossRef]
- Deacon, N.J.; Cavender-Bares, J. Limited pollen dispersal contributes to population genetic structure but not local adaptation in Quercus oleoides forests of Costa Rica. PLoS ONE 2015, 10, e0138783. [Google Scholar] [CrossRef] [Green Version]
- Moracho, E.; Moreno, G.; Jordano, P.; Hampe, A. Unusually limited pollen dispersal and connectivity of Pedunculate oak (Quercus robur) refugial populations at the species’ southern range margin. Mol. Ecol. 2016, 25, 3319–3331. [Google Scholar] [CrossRef]
- Buschbom, J.; Yanbaev, Y.; Degen, B. Efficient long-distance gene flow into an isolated relict oak stand. J. Hered. 2011, 102, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Anguiano, C.J.; Alcántar, J.J.; Toledo, B.R.; Tapia, L.M.; Vidales-Fernández, J.A. Caracterización edafoclimática del área productora de aguacate de Michoacán, México. In Proceedings of the VI World Avocado Congress, Viña Del Mar, Chile, 12 November 2007; ISBN 978-956-17-0413-8. [Google Scholar]
- Dubrovina, I.A.; Bautista, F. Analysis of the suitability of various soil groups and types of climate for avocado growing in the state of Michoacán, Mexico. Eurasian Soil Sci. 2014, 47, 491–503. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Copernicus Land Monitoring Services. SENTINEL-2. 2019. Available online: https://sentinels.copernicus.eu/web/sentinel/thematic-areas/land-monitoring/land-cover-use-and-change-detection-mapping (accessed on 15 January 2022).
- Congedo, L. Semi-Automatic Classification Plugin: A Python tool for the download and processing of remote sensing images in QGIS. J. Open Source Softw. 2021, 6, 3172. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2021. Available online: http://qgis.osgeo.org (accessed on 15 January 2022).
- Mas, J.F.; Pérez-Vega, A.; Ghilardi, A.; Martínez, S.; Loya-Carrillo, J.O.; Vega, E. A suite of tools for assessing thematic map accuracy. Geogr. J. 2014, 2014, 372349. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, P.; Foody, G.M.; Stehman, S.V.; Woodcock, C.E. Making better use of accuracy data in land change studies: Estimating accuracy and area and quantifying uncertainty using stratified estimation. Remote Sens. Environ. 2013, 129, 122–131. [Google Scholar] [CrossRef]
- Olofsson, P.; Foody, G.M.; Herold, M.; Stehman, S.V.; Woodcock, C.E.; Wulder, M.A. Good practices for estimating area and assessing accuracy of land change. Remote Sens. Environ. 2014, 148, 42–57. [Google Scholar] [CrossRef]
- Card, D.H. Using known map category marginal frequencies to improve estimates of thematic map accuracy. Photogramm. Eng. Remote Sens. 1982, 48, 431–439. [Google Scholar]
- Stehman, S.V. Model-assisted estimation as a unifying framework for estimating the area of landcover and land-cover change from remote sensing. Remote Sens. Environ. 2009, 113, 2455–2462. [Google Scholar] [CrossRef]
- Soares-Filho, B.S.; Rodrigues, H.O.; Costa, W.; Schlesinger, P. Modeling Environmental Dynamics with Dinamica EGO; Centro de Sensoriamento Remoto, Universidade Federal de Minas Gerais: Belo Horizonte, Brazil, 2009; p. 115. [Google Scholar]
- Granados, C.; Serrano Giné, D.; García-Romero, A. Efecto de borde en la composición y estructura de los bosques templados. Sierra de Monte-Alto, centro de México. Caldasia 2014, 36, 269–287. [Google Scholar] [CrossRef]
- Cázares, Y.; Vergara, P.M.; García-Romero, A. Regeneration of Quercus spp. along interactive forest boundaries in a fragmented peri-urban landscape of Mexico City. Environ. Conserv. 2020, 47, 39–45. [Google Scholar] [CrossRef]
- McGarigal, K.; Marks, B.J. Spatial Pattern Analysis Program for Quantifying Landscape Structure; General Technical Reports PNW-GTR-351; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Corvallis, OR, USA, 1995; Available online: https://andrewsforest.oregonstate.edu/sites/default/files/lter/pubs/pdf/pub1538.pdf (accessed on 15 January 2022s).
- Godínez-Gómez, O.; Correa Ayram, C.A. Makurhini: Analyzing landscape connectivity. Zeonodo 2020. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 15 January 2022).
- Saura, S.; Rubio, L. A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography 2010, 33, 523–537. [Google Scholar] [CrossRef]
- Neel, M.C. Patch connectivity and genetic diversity conservation in the federally endangered and narrowly endemic plant species Astragalus albens (Fabaceae). Biol. Conserv. 2008, 141, 938–955. [Google Scholar] [CrossRef]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). Índice de Impacto Humano en la Biodiversidad Terrestre. MEXBIO 2.0. 2014. Available online: https://www.conabio.gob.mx/informacion/gis/ (accessed on 1 October 2022).
- Alkemade, R.; van Oorschot, M.; Miles, L. GLOBIO3: A Framework to Investigate Options for Reducing Global Terrestrial Biodiversity Loss. Ecosystems 2009, 12, 374–390. [Google Scholar] [CrossRef] [Green Version]
- Urban, D.; Keitt, T. Landscape connectivity: A graph-theoretic perspective. Ecology 2001, 82, 1205–1218. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Saura, S.; Estreguil, C.; Mouton, C.; Rodríguez-Freire, M. Network analysis to assess landscape connectivity trends: Application to European forests (1990–2000). Ecol. Indic. 2011, 11, 407–416. [Google Scholar] [CrossRef]
- Gómez-Tagle, A.F.; Gómez-Tagle, A.; Fuerte-Velázquez, D.J.; Barajas-Alcalá, A.G.; Quiroz-Rivera, F.; Alarcón-Chaires, P.E.; Guerrero-García-Rojas, H. Blue and Green Water Footprint of Agro-Industrial Avocado Production in Central Mexico. Sustainability 2022, 14, 9664. [Google Scholar] [CrossRef]
- Zamora-Torres, A.I.; Baez-Figueroa, I. Profitability and comparative advantage: Analysis of strawberry production in Michoacán, Mexico. Agro Product. 2022, 15, 89–97. [Google Scholar] [CrossRef]
- López-Mendoza, A.; Oyama, K.; Pineda-García, F.; Aguilar-Romero, R. Defining Conservation Priorities for Oak Forests in Central Mexico Based on Networks of Connectivity. Forests 2022, 13, 1085. [Google Scholar] [CrossRef]
- Correa-Ayram, C.A.; Mendoza, M.E.; Salicrup, D.R.P.; Granados, E.L. Identifying potential conservation areas in the Cuitzeo Lake basin, Mexico by multitemporal analysis of landscape connectivity. J. Nat. Conserv. 2014, 22, 424–435. [Google Scholar] [CrossRef]
- Fuentes-Montemayor, E.; Watts, K.; Macgregor, N.A.; Lopez-Gallego, Z.; Park, K. Species mobility and landscape context determine the importance of local and landscape-level attributes. Ecol. Appl. 2017, 27, 1541–1554. [Google Scholar] [CrossRef] [Green Version]
- Szangolies, L.; Rohwäder, M.S.; Jeltsch, F. Single large AND several small habitat patches: A community perspective on their importance for biodiversity. Basic Appl. Ecol. 2022, 65, 16–27. [Google Scholar] [CrossRef]
- Herrera-Arroyo, M.L.; Sork, V.L.; González-Rodríguez, A.; Rocha-Ramírez, V.; Vega, E.; Oyama, K. Seed-mediated connectivity among fragmented populations of Quercus castanea (Fagaceae) in a Mexican landscape. Am. J. Bot. 2013, 100, 1663–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyama, K.; Herrera-Arroyo, M.L.; Rocha-Ramírez, V.; Benítez-Malvido, J.; Ruiz-Sánchez, E.; González-Rodríguez, A. Gene flow interruption in a recently human-modified landscape: The value of isolated trees for the maintenance of genetic diversity in a Mexican endemic red oak. For. Ecol. Manag. 2017, 390, 27–35. [Google Scholar] [CrossRef]
- Ramírez Enríquez, E.; Delgado Valerio, P.; García Magaña, J.J.; Molina Sánchez, A. Diversidad genética y conservación de pinos nativos de la cuenca del río Cupatitzio, en Michoacán. Rev. Mex. Cienc. For. 2019, 10, 4–32. [Google Scholar] [CrossRef] [Green Version]
- Méndez Solórzano, M.I. Efecto de la Fragmentación de Bosque en la Franja Aguacatera Sobre Caracteres Funcionales Foliares y la Diversidad y Estructura Genética de Quercus castanea. Master Thesis, Universidad Nacional Autónoma de México, Mexico City, Mexico, 2021. [Google Scholar]
- Correa Ayram, C.A.; Mendoza, M.E.; Etter, A.; Pérez-Salicrup, D.R. Effect of the landscape matrix condition for prioritizing multispecies connectivity conservation in a highly biodiverse landscape of Central Mexico. Reg. Environ. Chang. 2019, 19, 149–163. [Google Scholar] [CrossRef]
- Visconti, P.; Elkin, C. Using connectivity metrics in conservation planning—When does habitat quality matter? Divers. Distrib. 2009, 15, 602–612. [Google Scholar] [CrossRef]
- Pérez-López, S.P. Regeneración natural de Quercus castanea Neé (Fagaceae) en Bosques fragmentados de la cuenca de Cuitzeo, Michoacán. Ph.D. Thesis, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Mexico, 2014. [Google Scholar]
- Kim, M.; Lee, S.; Lee, S.; Yi, K.; Kim, H.S.; Chung, S.; Yoon, T.K. Seed Dispersal Models for Natural Regeneration: A Review and Prospects. Forests 2002, 13, 659. [Google Scholar] [CrossRef]
- Hsiao, L.-H.; Cheng, K.-S. Assessing Uncertainty in LULC Classification Accuracy by Using Bootstrap Resampling. Remote Sens. 2016, 8, 705. [Google Scholar] [CrossRef] [Green Version]
- Sáenz-Romero, C.; Mendoza-Maya, E.; Gómez-Pineda, E.; Blanco-García, A.; Endara-Agramont, A.R.; Lindig-Cisneros, R.; Vargas-Hernández, J.J. Recent evidence of Mexican temperate forest decline and the need for ex situ conservation, assisted migration, and translocation of species ensembles as adaptive management to face projected climatic change impacts in a megadiverse country. Can. J. For. Res. 2020, 50, 843–854. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Area (ha) | % Land | UA (%) | CI (%) | PA (%) | CI (%) |
|---|---|---|---|---|---|---|
| Temperate forests | 786,812 | 19.8 | 91.11 | 4.16 | 0.59 | 0.04 |
| Avocado orchards | 244,705 | 6.1 | 65.61 | 7.43 | 0.64 | 0.11 |
| Water bodies | 79,621 | 2.0 | 91.45 | 5.07 | 0.85 | 0.13 |
| Agriculture and pastures | 1,590,748 | 40.0 | 55.63 | 5.78 | 0.78 | 0.04 |
| Settlements | 180,329 | 4.5 | 31.29 | 7.50 | 0.77 | 0.20 |
| Dry forest and shrublands | 1,097,680 | 27.6 | 75.96 | 5.81 | 0.68 | 0.05 |
| Overall | 3,979,896 | 100 | 68.40 | 2.98 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latorre-Cárdenas, M.C.; González-Rodríguez, A.; Godínez-Gómez, O.; Arima, E.Y.; Young, K.R.; Denvir, A.; García-Oliva, F.; Ghilardi, A. Estimating Fragmentation and Connectivity Patterns of the Temperate Forest in an Avocado-Dominated Landscape to Propose Conservation Strategies. Land 2023, 12, 631. https://doi.org/10.3390/land12030631
Latorre-Cárdenas MC, González-Rodríguez A, Godínez-Gómez O, Arima EY, Young KR, Denvir A, García-Oliva F, Ghilardi A. Estimating Fragmentation and Connectivity Patterns of the Temperate Forest in an Avocado-Dominated Landscape to Propose Conservation Strategies. Land. 2023; 12(3):631. https://doi.org/10.3390/land12030631
Chicago/Turabian StyleLatorre-Cárdenas, María Camila, Antonio González-Rodríguez, Oscar Godínez-Gómez, Eugenio Y. Arima, Kenneth R. Young, Audrey Denvir, Felipe García-Oliva, and Adrián Ghilardi. 2023. "Estimating Fragmentation and Connectivity Patterns of the Temperate Forest in an Avocado-Dominated Landscape to Propose Conservation Strategies" Land 12, no. 3: 631. https://doi.org/10.3390/land12030631