

Nitrogenous and Phosphorus Soil Contents in Tierra del Fuego Forests: Relationships with Soil Organic Carbon, Climate, Vegetation and Landscape Metrics

 , , ,
, , ,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adhikari, K.; Hartemink, A. Linking soils to ecosystem services: A global review. Geoderma 2016, 262, 101–111. [Google Scholar] [CrossRef]
- Bouma, J. Soil science contributions towards sustainable development goals and their implementation: Linking soil functions with ecosystem services. J. Plant Nutr. Soil Sci. 2014, 177, 111–120. [Google Scholar] [CrossRef]
- Chen, S.; Arrouays, D.; Mulder, V.; Poggio, L.; Minasny, B.; Roudier, P.; Libohova, Z.; Lagacherie, P.; Shi, Z.; Hannam, J.; et al. Digital mapping of GlobalSoilMap soil properties at a broad scale: A review. Geoderma 2022, 409, e115567. [Google Scholar] [CrossRef]
- Forkuor, G.; Hounkpatin, O.; Welp, G.; Thiel, M. High resolution mapping of soil properties using remote sensing variables in South-Western Burkina Faso: A comparison of machine learning and multiple linear regression models. PLoS ONE 2017, 12, e0170478. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Prentice, S.; Tran, T.; Bingham, N.; King, J.; Chadwick, O. Modeling deep soil properties on California grassland hillslopes using LiDAR digital elevation models. Geoderma Reg. 2016, 7, 67–75. [Google Scholar] [CrossRef]
- Rodrigo-Comino, J.; Senciales, J.; Cerdà, A.; Brevik, E. The multidisciplinary origin of soil geography: A review. Earth Sci. Rev. 2018, 177, 114–123. [Google Scholar] [CrossRef]
- Zeraatpisheh, M.; Ayoubi, S.; Jafari, A.; Tajik, S.; Finke, P. Digital mapping of soil properties using multiple machine learning in a semiarid region, central Iran. Geoderma 2019, 338, 445–452. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, C.; He, N.; Wen, X.; Gao, Y.; Li, S.; Niu, S.; Butterbach-Bahl, K.; Luo, Y.; Yu, G. A global synthesis of the rate and temperature sensitivity of soil nitrogen mineralization: Latitudinal patterns and mechanisms. Glob. Chang. Biol. 2017, 23, 455–464. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Q.; Adhikari, K.; Jia, S.; Jin, X.; Liu, H. Spatial-temporal changes of soil organic carbon content in Wafangdian, China. Sustainability 2016, 8, 1154. [Google Scholar] [CrossRef]
- Wang, S.; Zhuang, Q.; Wang, Q.; Jin, X.; Han, C. Mapping stocks of soil organic carbon and soil total nitrogen in Liaoning Province of China. Geoderma 2017, 305, 250–263. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.; Schleucher, J.; Gröbner, G.; Vestergren, J.; Persson, P.; Jansson, M.; Giesler, R. Changes in organic phosphorus composition in boreal forest humus soils: The role of iron and aluminium. Biogeochemistry 2012, 108, 485–499. [Google Scholar] [CrossRef]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Hu, B.; Yang, B.; Pang, X.; Bao, W.; Tian, G. Responses of soil phosphorus fractions to gap size in a reforested spruce forest. Geoderma 2016, 279, 61–69. [Google Scholar] [CrossRef]
- McDowell, R.; Stewart, I. The phosphorus composition of contrasting soils in pastoral, native and forest management in Otago, New Zealand: Sequential extraction and 31P NMR. Geoderma 2006, 130, 176–189. [Google Scholar] [CrossRef]
- Hu, B.; Jia, Y.; Zhao, Z.; Li, F.; Siddique, H. Soil P availability, inorganic P fractions and yield effect in a calcareous soil with plastic-film-mulched spring wheat. Field Crop Res. 2012, 137, 221–229. [Google Scholar] [CrossRef]
- McBratney, A.; Santos, M.; Minasny, B. On digital soil mapping. Geoderma 2003, 117, 3–52. [Google Scholar] [CrossRef]
- Batjes, N. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Arrouays, D.; Deslais, W.; Badeau, V. The carbon content of topsoil and its geographical distribution in France. Soil Use Manag. 2001, 17, 7–11. [Google Scholar] [CrossRef]
- Adhikari, K.; Hartemink, A.; Minasny, B.; Kheir, R.; Greve, M.; Greve, M. Digital mapping of soil organic carbon contents and stocks in Denmark. PLoS ONE 2014, 9, e105519. [Google Scholar] [CrossRef]
- Yang, R.; Zhang, G.; Yang, F.; Zhi, J.; Yang, F.; Liu, F.; Zhao, Y.; Li, D. Precise estimation of soil organic carbon stocks in the northeast Tibetan Plateau. Sci. Rep. 2016, 6, e21842. [Google Scholar] [CrossRef]
- Khaledian, Y.; Brevik, E.; Pereira, P.; Cerdà, A.; Fattah, M.; Tazikeh, H. Modeling soil cation exchange capacity in multiple countries. Catena 2017, 158, 194–200. [Google Scholar] [CrossRef]
- Martin, M.; Orton, T.; Lacarce, E.; Meersmans, J.; Saby, N.; Paroissien, J.; Joliveta, C.; Boulonnea, L.; Arrouays, D. Evaluation of modelling approaches for predicting the spatial distribution of soil organic carbon stocks at the national scale. Geoderma 2014, 223, 97–107. [Google Scholar] [CrossRef]
- Minasny, B.; Malone, B.; McBratney, A.; Angers, D.; Arrouays, D.; Chambers, A.; Chaplot, V.; Chen, Z.; Cheng, K.; Das, B.; et al. Soil carbon 4 per mille. Geoderma 2017, 292, 59–86. [Google Scholar] [CrossRef]
- Peri, P.L.; Rosas, Y.M.; Ladd, B.; Toledo, S.; Lasagno, R.; Martínez Pastur, G. Modelling soil carbon content in South Patagonia and evaluating changes according to climate, vegetation, desertification and grazing. Sustainability 2018, 10, 438. [Google Scholar] [CrossRef]
- Martínez Pastur, G.; Aravena Acuña, M.C.; Silveira, E.; von Müller, A.; La Manna, L.; González-Polo, M.; Chaves, J.; Cellini, J.M.; Lencinas, M.V.; Radeloff, V.; et al. Mapping soil organic carbon in Patagonian forests based on climate, topography and vegetation metrics from satellite imagery. Remote Sens. 2022, 14, 5702. [Google Scholar] [CrossRef]
- Minasny, B.; Hartemink, A. Predicting soil properties in the tropics. Earth Sci. Rev. 2011, 106, 52–62. [Google Scholar] [CrossRef]
- Gonzalez Musso, R.; Oddi, F.; Goldenberg, M.; Garibaldi, L. Applying unmanned aerial vehicles (UAVs) to map shrubland structural attributes in northern Patagonia, Argentina. Can. J. For. Res. 2020, 50, 615–623. [Google Scholar] [CrossRef]
- Silveira, E.; Radeloff, V.; Martínez Pastur, G.; Martinuzzi, S.; Politi, N.; Lizarraga, L.; Rivera, L.; Gavier Pizarro, G.; Yin, H.; Rosas, Y.M.; et al. Forest phenoclusters for Argentina based on vegetation phenology and climate. Ecol. Appl. 2022, 32, e2526. [Google Scholar] [CrossRef]
- Silveira, E.; Radeloff, V.; Martinuzzi, S.; Martínez Pastur, G.; Bono, J.; Politi, N.; Lizarraga, L.; Rivera, L.; Ciuffoli, L.; Rosas, Y.M.; et al. Nationwide native forest structure maps for Argentina based on forest inventory data, SAR Sentinel-1 and vegetation metrics from Sentinel-2 imagery. Remote Sens. Environ. 2023, 285, e113391. [Google Scholar] [CrossRef]
- Gasparri, I.; Parmuchi, M.; Bono, J.; Karszenbaum, H.; Montenegro, C. Assessing multi-temporal Landsat 7 ETM + images for estimating above-ground biomass in subtropical dry forests of Argentina. J. Arid Environ. 2010, 74, 1262–1270. [Google Scholar] [CrossRef]
- Gasparri, I.; Baldi, G. Regional patterns and controls of biomass in semiarid woodlands: Lessons from the Northern Argentina Dry Chaco. Reg. Environ. Chang. 2013, 13, 1131–1144. [Google Scholar] [CrossRef]
- Bouvier, M.; Durrieu, S.; Fournier, R.; Renaud, J. Generalizing predictive models of forest inventory attributes using an area-based approach with airborne LiDAR data. Remote Sens. Environ. 2015, 156, 322–334. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Huang, C. Spatial variability of soil total nitrogen and soil total phosphorus under different land uses in a small watershed on the Loess Plateau, China. Geoderma 2009, 150, 141–149. [Google Scholar] [CrossRef]
- Wang, S.; Jin, X.; Adhikari, K.; Li, W.; Yu, M.; Bian, Z.; Wang, Q. Mapping total soil nitrogen from a site in northeastern China. Catena 2018, 166, 134–146. [Google Scholar] [CrossRef]
- Chi, Y.; Zhao, M.; Sun, J.; Xie, Z.; Wang, E. Mapping soil total nitrogen in an estuarine area with high landscape fragmentation using a multiple-scale approach. Geoderma 2019, 339, 70–84. [Google Scholar] [CrossRef]
- Diehl, P.; Mazzarino, M.; Funes, F.; Fontenla, S.; Gobbi, M.; Ferrari, J. Nutrient conservation strategies in native Andean-Patagonian forests. J. Veg. Sci. 2003, 14, 63–70. [Google Scholar] [CrossRef]
- Peri, P.L.; Gargaglione, V.; Martínez Pastur, G. Above and belowground nutrients storage and biomass accumulation in marginal Nothofagus antarctica forests in Southern Patagonia. For. Ecol. Manag. 2008, 255, 2502–2511. [Google Scholar] [CrossRef]
- Bahamonde, H.; Peri, P.L.; Alvarez, R.; Barneix, A.; Moretto, A.; Martínez Pastur, G. Litter decomposition and nutrients dynamics in Nothofagus antarctica forests under silvopastoral use in Southern Patagonia. Agrofor. Syst. 2012, 84, 345–360. [Google Scholar] [CrossRef]
- Bahamonde, H.; Peri, P.L.; Alvarez, R.; Barneix, A.; Moretto, A.; Martínez Pastur, G. Silvopastoral use of Nothofagus antarctica in Southern Patagonian forests, influence over net nitrogen soil mineralization. Agrofor. Syst. 2013, 87, 259–271. [Google Scholar] [CrossRef]
- Bahamonde, H.; Peri, P.L.; Martínez Pastur, G.; Monelos, L. Litterfall and nutrients return in Nothofagus antarctica forests growing in a site quality gradient with different management uses in Southern Patagonia. Eur. J. For. Res. 2015, 134, 113–124. [Google Scholar] [CrossRef]
- Peri, P.L.; Lasagno, R.; Martínez Pastur, G.; Atkinson, R.; Thomas, E.; Ladd, B. Soil carbon is a useful surrogate for conservation planning in developing nations. Sci. Rep. 2019, 9, e3905. [Google Scholar] [CrossRef]
- Besteiro, S.; Descalzo, A. Contenidos de nitrógeno y fósforo del suelo ante un cambio de cobertura y condición topográfica. Rev. Inv. Agrop. 2021, 47, 285–292. [Google Scholar]
- Chaves, J.; Lencinas, M.V.; Cellini, J.M.; Peri, P.L.; Martínez Pastur, G. Changes in nutrients and fibre tissue contents in Nothofagus pumilio trees growing at site quality and crown class gradients. For. Ecol. Manag. 2022, 505, e119910. [Google Scholar] [CrossRef]
- Rosas, Y.M.; Peri, P.L.; Martínez Pastur, G. Assessment of provisioning ecosystem services in terrestrial ecosystems of Santa Cruz province, Argentina. In Ecosystem Services in Patagonia: A Multi-Criteria Approach for an Integrated Assessment; Peri, P.L., Nahuelhual, L., Martínez Pastur, G., Eds.; Natural and Social Sciences of Patagonia; Springer Nature: Cham, Switzerland, 2021; pp. 19–46. [Google Scholar]
- Rosas, Y.M.; Martínez Pastur, G.; Peri, P.L. Servicios Ecosistémicos y Biodiversidad de los Recursos Naturales de Santa Cruz; INTA: Buenos Aires, Argentina, 2022; 60p. [Google Scholar]
- Martínez Pastur, G.; Peri, P.L.; Soler, R.; Schindler, S.; Lencinas, M.V. Biodiversity potential of Nothofagus forests in Tierra del Fuego (Argentina): Tool proposal for regional conservation planning. Biodiv. Conserv. 2016, 25, 1843–1862. [Google Scholar] [CrossRef]
- Gutsch, M.; Lasch-Born, P.; Kollas, C.; Suckow, F.; Reyer, C. Balancing trade-offs between ecosystem services in Germany’s forests under climate change. Environ. Res. Let. 2018, 13, e045012. [Google Scholar] [CrossRef]
- Orsi, F.; Ciolli, M.; Primmer, E.; Varumo, L.; Geneletti, D. Mapping hotspots and bundles of forest ecosystem services across the European Union. Land Use Pol. 2020, 99, e104840. [Google Scholar] [CrossRef]
- Peri, P.L.; Ladd, B.; Lasagno, R.; Martínez Pastur, G. The effects of land management (grazing intensity) vs. the effects of topography, soil properties, vegetation type, and climate on soil carbon concentration in Southern Patagonia. J. Arid Environ. 2016, 134, 73–78. [Google Scholar] [CrossRef]
- Köhl, M.; Lasco, R.; Cifuentes, M.; Jonsson, O.; Korhonen, K.; Mundhenk, P.; de Jesus Navar, J.; Stinson, G. Changes in forest production, biomass and carbon: Results from the 2015 UN FAO Global Forest Resource Assessment. For. Ecol. Manage. 2015, 352, 21–34. [Google Scholar] [CrossRef]
- Moghaddas, E.; Stephens, S. Thinning, burning, and thin-burn fuel treatment effects on soil properties in a Sierra Nevada mixed-conifer forest. For. Ecol. Manag. 2007, 250, 156–166. [Google Scholar] [CrossRef]
- Nilsen, P.; Strand, L. Thinning intensity effects on carbon and nitrogen stores and fluxes in a Norway spruce (Picea abies (L.) Karst.) stand after 33 years. For. Ecol. Manag. 2008, 256, 201–208. [Google Scholar] [CrossRef]
- Nunery, J.; Keeton, W. Forest carbon storage in the north-eastern United States: Net effects of harvesting frequency, post-harvest retention, and wood products. For. Ecol. Manag. 2010, 259, 1363–1375. [Google Scholar] [CrossRef]
- Ontl, T.; Janowiak, M.; Swanston, C.; Daley, J.; Handler, S.; Cornett, M.; Hagenbuch, S.; Handrick, C.; Mccarthy, L.; Patch, N. Forest management for carbon sequestration and climate adaptation. J. For. 2019, 118, 86–101. [Google Scholar] [CrossRef]
- La Manna, L.; Tarabini, M.; Gomez, F.; Rostagno, C. Changes in soil organic matter associated with afforestation affect erosion processes: The case of erodible volcanic soils from Patagonia. Geoderma 2021, 403, e115265. [Google Scholar] [CrossRef]
- Gómez, F.; von Müller, A.; Tarabini, M.; La Manna, L. Resilient Andisols under silvopastoral systems. Geoderma 2022, 418, e115843. [Google Scholar] [CrossRef]
- Pang, X.; Bao, W.; Zhu, B.; Cheng, W. Response of soil respiration and its temperature sensitivity to thinning in a pine plantation. Agric. For. Meteorol. 2013, 171, 57–64. [Google Scholar] [CrossRef]
- Bolat, I. The effect of thinning on microbial biomass C, N and basal respiration in black pine forest soils in Mudurnu, Turkey. Eur. J. For. Res. 2014, 133, 131–139. [Google Scholar] [CrossRef]
- Dawes, M.; Schleppi, P.; Hattenschwiler, S.; Rixen, C.; Hagedorn, F. Soil warming opens the nitrogen cycle at the alpine treeline. Glob. Chang. Biol. 2017, 23, 421–434. [Google Scholar] [CrossRef]
- Burke, I.; Lauenroth, W.; Parton, W. Regional and temporal variation in net primary production and nitrogen mineralization in grasslands. Ecology 1997, 78, 1330–1340. [Google Scholar] [CrossRef]
- Li, Z.; Tian, D.; Wang, B.; Wang, J.; Wang, S.; Chen, H.; Xu, X.; Wang, C.; He, N.; Niu, S. Microbes drive global soil nitrogen mineralization and availability. Glob. Chang. Biol. 2019, 25, 107–1088. [Google Scholar] [CrossRef]
- Verhoef, H.; Brussaard, L. Decomposition and nitrogen mineralization in natural and agroecosystems: The contribution of soil animals. Biogeochemistry 1990, 11, 175–211. [Google Scholar] [CrossRef]
- Maaroufi, N.; De Long, J. Global change impacts on forest soils: Linkage between soil biota and carbon-nitrogen-phosphorus stoichiometry. Front. For. Glob. Chang. 2020, 3, e16. [Google Scholar] [CrossRef]
- Thornton, P.; Lamarque, J.; Rosenbloom, N.; Mahowald, N. Influence of carbon-nitrogen cycle coupling on land model response to CO2 fertilization and climate variability. Glob. Biogeochem. Cycles 2007, 21, GB4018. [Google Scholar] [CrossRef]
- Luque, S.; Martínez Pastur, G.; Echeverría, C.; Pacha, M. Overview of biodiversity loss in South America: A landscape perspective for sustainable forest management and conservation in temperate forests. In Landscape Ecology and Forest Management: Challenges and Solutions in a Changing Globe; Li, C., Lafortezza, R., Chen, J., Eds.; HEP-Springer: Amsterdam, The Netherlands, 2010; pp. 352–379. [Google Scholar]
- Angelstam, P.; Albulescu, C.; Andrianambinina, O.; Aszalós, R.; Borovichev, E.; Cano Cardona, W.; Dobrynin, D.; Fedoriak, M.; Firm, D.; Hunter, M.; et al. Frontiers of protected areas versus forest exploitation: Assessing habitat network functionality in 16 case study regions globally. AMBIO 2021, 50, 2286–2310. [Google Scholar] [CrossRef]
- Martínez Pastur, G.; Schlichter, T.; Matteucci, S.; Gowda, J.; Huertas Herrera, A.; Toro Manríquez, M.; Lencinas, M.V.; Cellini, J.M.; Peri, P.L. Synergies and trade-offs of national conservation policy and agro-Forestry management over forest loss in Argentina during the last decade. In Latin AMERICA in Times of Global Environmental Change; Lorenzo, C., Ed.; The Latin American Studies Book Series; Springer: Cham, Switzerland, 2020; pp. 135–155. [Google Scholar]
- Ministerio de Ambiente y Desarrollo Sustentable (MAyDS). Informe de Estado de Implementación 2010–2016 de la Ley N°26.331 de Presupuestos Mínimos de Protección Ambiental de los Bosques Nativos; Ministerio de Ambiente y Desarrollo Sostenible de la Nación: Buenos Aires, Argentina, 2017; 80p. [Google Scholar]
- Wang, S.; Huang, M.; Shao, X.; Mickler, R.; Li, K.; Ji, J. Vertical distribution of soil organic carbon in China. Environ. Manag. 2004, 33, 200–209. [Google Scholar] [CrossRef]
- Dirección Nacional de Bosques. Datos del Segundo Inventario Nacional de Bosques Nativos de la República Argentina; Ministerio de Ambiente y Desarrollo Sostenible de la Nación: Buenos Aires, Argentina, 2021; 67p. [Google Scholar]
- Hansen, M.; Potapov, P.; Moore, R.; Hancher, M.; Turubanova, S.; Tyukavina, A.; Thau, D.; Stehmans, S.; Goetzt, S.; Loveland, T.; et al. High-resolution global maps of 21st-Century forest cover change. Science 2013, 80, 850–853. [Google Scholar] [CrossRef]
- Peri, P.L.; Martínez Pastur, G.; Schlichter, T. Uso Sustentable del Bosque: Aportes Desde la Silvicultura Argentina; Ministerio de Ambiente y Desarrollo Sostenible de la Nación Argentina: Buenos Aires, Argentina, 2021; 888p. [Google Scholar]
- Carter, M.; Gregorich, E. Soil Sampling and Methods of Analysis, 2nd ed.; Canadian Society of Soil Science; Taylor and Francis: Boca Ratón, FL, USA, 2007; 1261p. [Google Scholar]
- Bray, R.; Kurtz, L. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute Inc.: Redlands, California, USA, 2011. [Google Scholar]
- Hijmans, R.; Cameron, S.; Parra, J.; Jones, P.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Climat. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Zomer, R.; Trabucco, A.; Bossio, D.; Van Straaten, O.; Verchot, L. Climate change mitigation: A spatial analysis of global land suitability for clean development mechanism afforestation and reforestation. Agric. Ecosyst. Environ. 2008, 126, 67–80. [Google Scholar] [CrossRef]
- Farr, T.; Rosen, P.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The shuttle radar topography mission. Rev. Geophys. 2007, 45, RG2004. [Google Scholar] [CrossRef]
- ORNL DAAC. MODIS Collection 5 Land Products Global Sub-Setting and Visualization Tool; ORNL DAAC: Oak Ridge, TN, USA, 2008. [Google Scholar]
- Zhao, M.; Running, S. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Peri, P.L.; Rosas, Y.M.; Ladd, B.; Toledo, S.; Lasagno, R.; Martínez Pastur, G. Modelling soil nitrogen content in South Patagonia across a climate gradient, vegetation type, and grazing. Sustainability 2019, 11, 2707. [Google Scholar] [CrossRef]
- Rosas, Y.M.; Peri, P.L.; Lencinas, M.V.; Lizarraga, L.; Martínez Pastur, G. Multi-taxon biodiversity assessment of Southern Patagonia: Supporting conservation strategies at different landscapes. J. Environ. Manag. 2022, 307, e114578. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, J.; Rosas, Y.M.; Lencinas, M.V.; Bortoluzzi, A.; Peri, P.L.; Martínez Pastur, G. Synergies and trade-offs among ecosystem services and biodiversity in different forest types inside and off-reserve in Tierra del Fuego, Argentina. In Ecosystem Services in Patagonia: A Multi-Criteria Approach for an Integrated Assessment; Peri, P.L., Nahuelhual, L., Martínez Pastur, G., Eds.; Natural and Social Sciences of Patagonia; Springer Nature: Cham, Switzerland, 2021; Chapter 4; pp. 75–97. [Google Scholar]
- Olivero, E.; Castro, L.; Scasso, R.; Fazio, A.; Miretzky, P. Fosfatos marinos del Paleógeno de la Isla Grande de Tierra del Fuego. Rev. Asoc. Geol. Argent. 1998, 53, 239–246. [Google Scholar]
- Selige, T.; Böhner, J.; Schmidhalter, U. High resolution topsoil mapping using hyperspectral image and field data in multivariate regression modeling procedures. Geoderma 2006, 136, 235–244. [Google Scholar] [CrossRef]
- Sumfleth, K.; Duttmann, R. Prediction of soil property distribution in paddy soil landscapes using terrain data and satellite information as indicators. Ecol. Indic. 2008, 8, 485–501. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, C.; Li, W. Predictive mapping of soil total nitrogen at a regional scale: A comparison between geographically weighted regression and cokriging. Appl. Geogr. 2013, 42, 73–85. [Google Scholar] [CrossRef]
- Hengl, T.; Heuvelink, G.; Kempen, B.; Leenaars, J.; Walsh, M.; Shepherd, K.; Tondoh, J. Mapping soil properties of Africa at 250m resolution: Random forests significantly improve current predictions. PLoS ONE 2015, 10, e0125814. [Google Scholar] [CrossRef]
- Bui, E.; Henderson, B.; Viergever, K. Knowledge discovery from models of soil properties developed through data mining. Ecol. Model. 2006, 191, 431–446. [Google Scholar] [CrossRef]
- Adhikari, K.; Kheir, R.; Greve, M.; Bøcher, P.; Malone, B.; Minasny, B.; Greve, M. High-resolution 3-D mapping of soil texture in Denmark. Soil Sci. Soc. Am. J. 2013, 77, 860–876. [Google Scholar] [CrossRef]
- Chang, C.; Laird, D.; Mausbach, M.; Hurburgh, C. Near infrared reflectance spectroscopy: Principal components regression analysis of soil properties. Soil Sci. Soc. Am. J. 2001, 65, 480–490. [Google Scholar] [CrossRef]
- Mosleh, Z.; Salehi, M.; Jafari, A.; Borujeni, I.; Mehnatkesh, A. The effectiveness of digital soil mapping to predict soil properties over low-relief areas. Environ. Monit. Assess. 2016, 188, e195. [Google Scholar] [CrossRef]
- Razakamanarivo, R.; Grinand, C.; Razafindrakoto, M.; Bernoux, M.; Albrecht, A. Mapping organic carbon stocks in eucalyptus plantations of the central highlands of Madagascar: A multiple regression approach. Geoderma 2011, 162, 335–346. [Google Scholar] [CrossRef]
- Jobbágy, E.; Jackson, R. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Bronson, K.; Zobeck, T.; Chua, T.; Acosta-Martínez, V.; van Pelt, R.; Booker, J. Carbon and nitrogen pools of southern high plains cropland and grassland soils. Soil Sci. Soc. Am. J. 2004, 68, 1695–1704. [Google Scholar] [CrossRef]
- Bardgett, R.; Streeter, T.; Bol, R. Soil microbes compete effectively with plants for organic-nitrogen inputs to temperate grasslands. Ecology 2003, 84, 1277–1287. [Google Scholar] [CrossRef]
- Follett, R.; Stewart, C.; Pruessner, E.; Kimble, J. Effects of climate change on soil carbon and nitrogen storage in the US Great Plains. J. Soil Water Conserv. 2012, 67, 331–342. [Google Scholar] [CrossRef]
- Thornton, P.; Doney, S.; Lindsay, K.; Moore, J.; Mahowald, N.; Randerson, J.; Fung, I.; Lamarque, J.; Feddema, J.; Lee, Y. Carbon-nitrogen interactions regulate climate-carbon cycle feedbacks: Results from an atmosphere-ocean general circulation model. Biogeosciences 2009, 6, 2099–2120. [Google Scholar] [CrossRef]
- Bahamonde, H.; Gargaglione, V.; Peri, P.L. Sheep feces decomposition and nutrient release across an environmental gradient in Southern Patagonia. Ecol. Austral 2017, 27, 18–28. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, A.; Rossiter, D.; Du, F.; Burt, J. Trustworthiness indicator to select sample points for the individual predictive soil mapping method (iPSM). Geoderma 2020, 373, e114440. [Google Scholar] [CrossRef]
- Baxter, S.; Oliver, M. The spatial prediction of soil mineral N and potentially available N using elevation. Geoderma 2005, 128, 325–339. [Google Scholar] [CrossRef]
- Viscarra Rossel, R.; Adamchuk, V.; Sudduth, K.; McKenzie, N.; Lobsey, C. Proximal soil sensing: An effective approach for soil measurements in space and time. Adv. Agron. 2011, 113, 243–291. [Google Scholar]
- Toro-Manríquez, M.; Soler, R.; Lencinas, M.V.; Promis, A. Canopy composition and site are indicative of mineral soil conditions in Patagonian mixed Nothofagus forests. Ann. For. Sci. 2019, 76, e117. [Google Scholar] [CrossRef]
- Adhikari, K.; Mishra, U.; Owens, P.; Libohova, Z.; Wills, S.; Riley, W.; Hoffman, F.; Smith, D. Importance and strength of environmental controllers of soil organic carbon changes with scale. Geoderma 2020, 375, e114472. [Google Scholar] [CrossRef]
- Dreiss, L.; Volin, J. Influence of leaf phenology and site nitrogen on invasive species establishment in temperate deciduous forest understories. For. Ecol. Manag. 2013, 296, 1–8. [Google Scholar] [CrossRef]
- Henn, J.; Anderson, C.; Kreps, G.; Lencinas, M.V.; Soler, R.; Martínez Pastur, G. Determining abiotic and biotic drivers that limit active riparian forest restoration in abandoned beaver meadows in Tierra del Fuego. Ecol. Rest. 2014, 32, 369–378. [Google Scholar] [CrossRef]
- Bahamonde, H.; Martínez Pastur, G.; Lencinas, M.V.; Soler, R.; Rosas, Y.M.; Ladd, B.; Duarte Guardia, S.; Peri, P.L. The relative importance of soil properties and regional climate as drivers of productivity in southern Patagonia’s Nothofagus antarctica forests. Ann. For. Sci. 2018, 75, e45. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef]
- Binkley, D.; Valentine, D. Fifty-year biogeochemical effects of green ash, white pine and Norway spruce in a replicated experiment. For. Ecol. Manag. 1991, 40, 13–25. [Google Scholar] [CrossRef]
- Rothe, A.; Binkley, D. Nutritional interactions in mixed species forests: A synthesis. Can. J. For. Res. 2001, 31, 1855–1870. [Google Scholar] [CrossRef]
- Binkley, D.; Giardina, C. Why do tree species affect soils? The warp and woof of tree-soil interactions. Biogeochemistry 1998, 42, 89–106. [Google Scholar] [CrossRef]
- Gerding, V.; Thiers, O. Caracterización de suelos bajo bosques de Nothofagus betuloides (Mirb) Blume, en Tierra del Fuego, Chile. Rev. Chilena Hist. Nat. 2002, 75, 819–833. [Google Scholar] [CrossRef]
- Romanya, J.; Fons, J.; Sauras-Yera, T.; Gutiérrez, E.; Vallejo, R. Soil-plant relationships and tree distribution in old growth Nothofagus betuloides and N. pumilio forests of Tierra del Fuego. Geoderma 2005, 124, 169–180. [Google Scholar] [CrossRef]
- Huygens, D.; Rütting, T.; Boeckx, P.; Van Cleemput, O.; Godoy, R.; Müller, C. Soil nitrogen conservation mechanisms in a pristine south Chilean Nothofagus forest ecosystem. Soil Biol. Biochem. 2007, 39, 2448–2458. [Google Scholar] [CrossRef]
- Gargaglione, V.; Gonzalez Polo, M.; Birgi, J.; Toledo, S.; Peri, P.L. Silvopastoral use of Nothofagus antarctica forests in Patagonia: Impact on soil microorganisms. Agrofor. Syst. 2022, 96, 957–968. [Google Scholar] [CrossRef]
- Podwojewski, P.; Poulenard, J.; Nguyet, M.; de Rouw, A.; Nguyen, V.; Pham, Q.; Tran, D. Climate and vegetation determine soil organic matter status in an alpine inner-tropical soil catena in the Fan Si Pan Mountain, Vietnam. Catena 2011, 87, 226–239. [Google Scholar] [CrossRef]
- Garten, C.; Hanson, P. Measured forest soil C stocks and estimated turnover times along an elevation gradient. Geoderma 2006, 136, 342–352. [Google Scholar] [CrossRef]
- Sánchez, P.; Ahamed, S.; Carre, F.; Hartemink, A.; Hempel, J.; Huising, J.; Lagacherie, P.; McBratney, A.; McKenzie, N.; Mendonça-Santos, M.; et al. Digital soil map of the world. Science 2009, 325, 680–681. [Google Scholar] [CrossRef]
- Chenu, C.; Angers, D.; Barre, P.; Derrien, D.; Arrouays, D.; Balesdent, J. Increasing organic stocks in agricultural soils: Knowledge gaps and potential innovations. Soil Tillage Res. 2019, 188, 41–52. [Google Scholar] [CrossRef]
- Moreno, A.; Neumann, M.; Hasenauer, H. Optimal resolution for linking remotely sensed and forest inventory data in Europe. Remote Sens. Environ. 2016, 183, 109–119. [Google Scholar] [CrossRef]
- Zald, H.; Wulder, M.A.; White, J.C.; Hilker, T.; Hermosilla, T.; Hobart, G.W.; Coops, N.C. Integrating Landsat pixel composites and change metrics with lidar plots to predictively map forest structure and aboveground biomass in Saskatchewan, Canada. Remote Sens. Environ. 2016, 176, 188–201. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Terra, M.; ter Steege, H.; Maeda, E.; Acerbi Júnior, F.; Scolforo, J. Carbon-diversity hotspots and their owners in Brazilian southeastern savanna, Atlantic forest and semi-arid woodland domains. For. Ecol. Manag. 2019, 452, e117575. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Espírito Santo, F.; Wulder, M.A.; Acerbi Júnior, F.; Carvalho, M.C.; Mello, C.R.; Mello, J.M.; Shimabukuro, Y.E.; Terra, M.; Carvalho, L.; et al. Pre-stratified modelling plus residuals kriging reduces the uncertainty of aboveground biomass estimation and spatial distribution in heterogeneous savannas and forest environments. For. Ecol. Manag. 2019, 445, 96–109. [Google Scholar] [CrossRef]
- White, J.; Wulder, M.; Hobart, G.; Luther, J.; Hermosilla, T.; Griffiths, P.; Coops, N.; Hall, R.; Hostert, P.; Dyk, A.; et al. Pixel-based image compositing for large area dense time series applications and science. Can. J. Remote Sens. 2014, 40, 192–212. [Google Scholar] [CrossRef]
- Yuping, L.; Ramzan, M.; Xincheng, L.; Murshed, M.; Awosusi, A.; Ibrahim, S.; Adebayo, T. Determinants of carbon emissions in Argentina: The roles of renewable energy consumption and globalization. Energy Rep. 2021, 7, 4747–4760. [Google Scholar] [CrossRef]
- Murshed, M.; Rashid, S.; Ulucak, R.; Dagar, V.; Rehman, A.; Alvarado, R.; Nathaniel, S. Mitigating energy production-based carbon dioxide emissions in Argentina: The roles of renewable energy and economic globalization. Environ. Sci. Pollut. Res. 2022, 29, 16939–16958. [Google Scholar] [CrossRef]
- Baldassini, P.; Bagnato, C.; Paruelo, J. How may deforestation rates and political instruments affect land use patterns and carbon emissions in the semi-arid Chaco, Argentina? Land Use Policy 2020, 99, e104985. [Google Scholar] [CrossRef]
- Aagesen, D. Crisis and conservation at the end of the world: Sheep ranching in Argentine Patagonia. Environ. Conserv. 2000, 27, 208–215. [Google Scholar] [CrossRef]
- Mazzonia, E.; Vazquez, M. Desertification in Patagonia. Dev. Earth Surface Proc. 2009, 13, 351–377. [Google Scholar]
- Peri, P.L.; Lasagno, R.; Chartier, M.; Roig, F.; Rosas, Y.M.; Martínez Pastur, G. Soil erosion rates and nutrient loss in rangelands of Southern Patagonia. In Imperiled: The Encyclopedia of Conservation; Della Sala, D., Goldstein, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; Volume 1–3, Chapter 9; pp. 102–110. [Google Scholar]
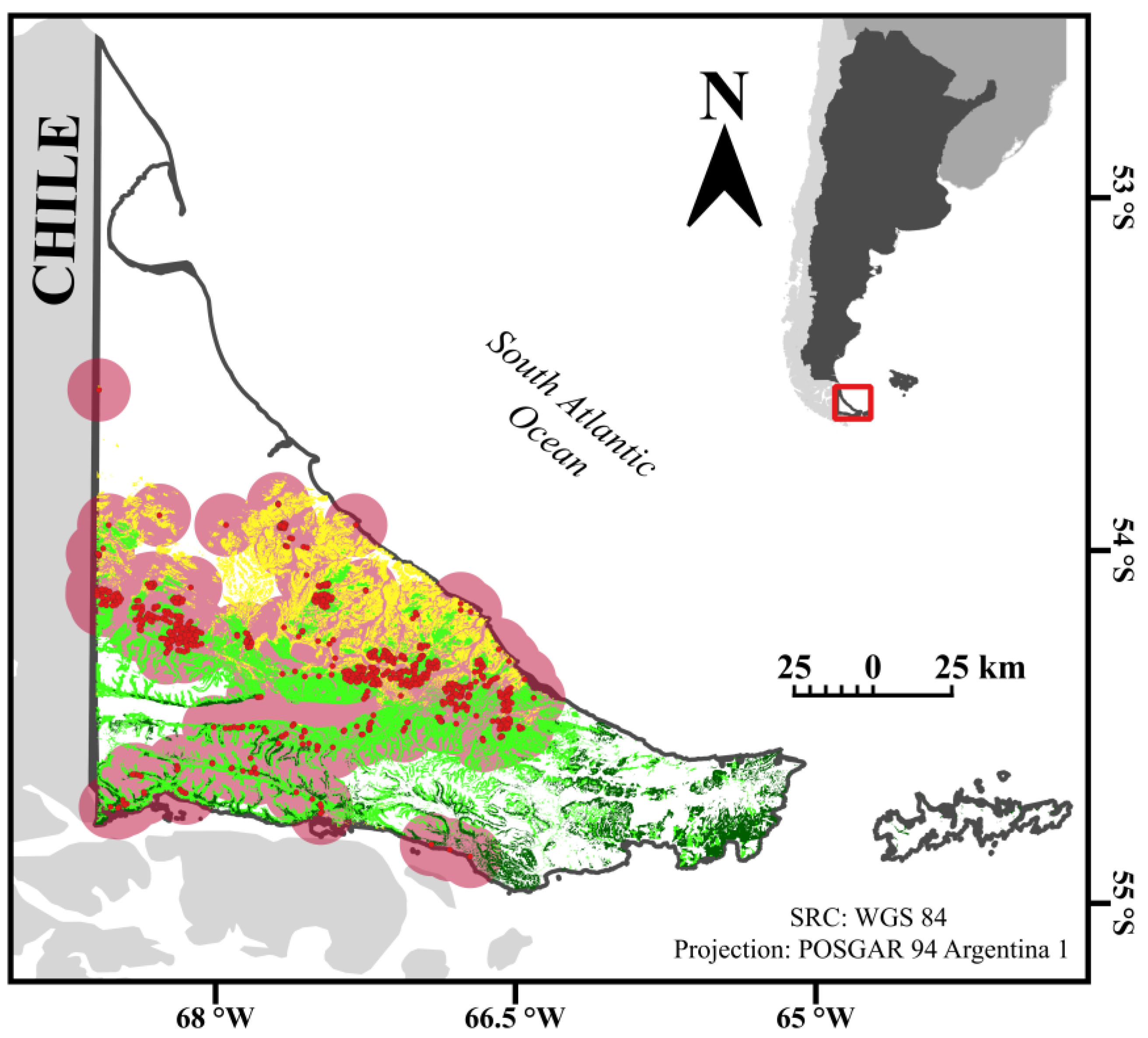



{kind=link}
{kind=link}
{kind=link}
| Forest Type | Area (km2) | Plots (n) | Sampling Effort (%) | ||
|---|---|---|---|---|---|
| NA | 2014.7 | 27.6% | 95 | 13.0% | −14.6% |
| NP | 4045.1 | 55.5% | 614 | 84.3% | +28.9% |
| MIX | 1232.6 | 16.9% | 19 | 2.6% | −14.3% |
| Total | 7292.4 | 728 | |||
| SN-GLOBAL | 0.0354235 × SOC + 0.224559 × BIO15 − 0.00281924 × NPP | |||
| R²-adj. = 92.1% | F(p) = 2824.9 (<0.01) | |||
| SEE = 2.8 | T(p) | SOC = 22.2 (<0.01) | BIO15 = 9.7 (<0.01) | |
| MAE = 2.1 | NPP = −4.6 (<0.01) | |||
| SN-NA | 0.0610033 × SOC + 3.3065 × BIO1 − 0.939747 × BIO15 | |||
| R²-adj. = 98.0% | F(p) = 1579.0 (<0.01) | |||
| SEE = 1.4 | T(p) | SOC = 15.0 (<0.01) | BIO1 = 7.4 (<0.01) | |
| MAE = 1.1 | BIO15 = −8.0 (<0.01) | |||
| SN-NP | 0.106466 × DH + 0.0312037 × BA + 0.0376064 × SOC − 0.00890734 × BIO12 + 2.40688 × NDVI | |||
| R²-adj. = 92.9% | F(p) = 1609.8 (<0.01) | |||
| SEE = 2.6 | T(p) | DH = 3.9 (<0.01) | BA = 4.2 (<0.01) | |
| MAE = 1.9 | SOC = 23.1 (<0.01) | BIO12 = −6.3 (<0.01) | ||
| NDVI = 3.7 (<0.01) | ||||
| SN-MIX | 0.0682924 × DH + 0.0174538 × SOC | |||
| R²-adj. = 95.8% | F(p) = 208.9 (<0.01) | |||
| SEE = 1.9 | T(p) | DH = 2.2 (0.04) | SOC = 7.1 (<0.01) | |
| MAE = 0.9 | ||||
| SP-GLOBAL | 0.00812562 × DH + 0.00833205 × SN − 0.000885286 × BIO16 | |||
| R²-adj. = 70.3% | F(p) = 574.58 (<0.01) | |||
| SEE = 0.08 | T(p) | DH = 12.8 (<0.01) | SN = 10.1 (<0.01) | |
| MAE = 0.06 | BIO16 = −8.4 (<0.01) | |||
| SP-NA | 0.0026018 × SN + 0.00692245 × BIO4 | |||
| R²-adj. = 83.5% | F(p) = 238.6 (<0.01) | |||
| SEE = 0.02 | T(p) | SN = 3.5 (<0.01) | BIO4 = 2.8 (<0.01) | |
| MAE = 0.01 | ||||
| SP-NP | 0.00840194 × SN + 0.094582 × BIO4 − 0.000500307 × BIO12 | |||
| R²-adj. = 71.4% | F(p) = 512.9 (<0.01) | |||
| SEE = 0.09 | T(p) | SN = 8.2 (<0.01) | BIO4 = 8.9 (<0.01) | |
| MAE = 0.06 | BIO12 = −7.8 (<0.01) | |||
| SP-MIX | −0.0014144 × DH + 0.000239921 × SOC + 0.000873535 × SLOPE | |||
| R²-adj. = 86.7% | F(p) = 39.9 (<0.01) | |||
| SEE = 0.02 | T(p) | DH = −2.4 (0.02) | SOC = 5.3 (<0.01) | |
| MAE = 0.01 | SLOPE = 2.1 (0.04) | |||
| Model | SN | Global | Individual | SP | Global | Individual | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| (ton ha−1) | SEE | MAE | SEE | MAE | (kg ha−1) | SEE | MAE | SEE | MAE | |
| NA | 10.25 | 0.72 | 1.97 | 0.01 | 1.14 | 48.84 | −13.14 | 27.19 | −0.04 | 16.53 |
| NP | 9.27 | 0.01 | 2.08 | <0.01 | 1.96 | 138.44 | 3.99 | 66.70 | −0.06 | 65.34 |
| MIX | 5.45 | −4.72 | 4.72 | −0.03 | 0.91 | 46.58 | −22.97 | 58.25 | −0.28 | 16.15 |
| Total | 9.30 | −0.02 | 2.14 | <0.01 | 1.82 | 124.35 | 1.05 | 61.32 | −0.07 | 59.38 |
| Type | Class | Area (km²) | SOC (ton ha−1) | Total SOC (mill ton) | SN (ton ha−1) | Total SN (mill ton) | SP (kg ha−1) | Total SP (thousand ton) |
|---|---|---|---|---|---|---|---|---|
| (A) | NA | 2014.7 | 141.3 (±22.3) | 28.5 | 7.7 (±1.7) | 1.56 | 45.0 (±20.6) | 9.1 |
| NP | 4045.1 | 158.7 (±22.8) | 64.2 | 6.9 (±2.1) | 2.81 | 129.8 (±41.6) | 52.5 | |
| MIX | 1232.6 | 184.5 (±30.6) | 22.7 | 4.9 (±1.1) | 0.60 | 77.2 (±25.0) | 9.5 | |
| (B) | Red | 2926.7 | 165.5 (±28.0) | 48.4 | 5.7 (±1.9) | 1.67 | 97.5 (±39.8) | 28.5 |
| Yellow | 3845.4 | 154.7 (±26.6) | 59.5 | 7.7 (±1.7) | 2.95 | 98.9 (±57.8) | 38.0 | |
| Green | 192.8 | 133.0 (±20.7) | 2.6 | 6.6 (±1.9) | 0.13 | 87.2 (±57.1) | 1.7 | |
| Unclassified | 327.5 | 151.6 (±27.2) | 5.0 | 6.5 (±2.1) | 0.21 | 86.6 (±46.5) | 2.8 | |
| (C) | National | 262.1 | 143.1 (±15.5) | 387 | 4.8 (±1.3) | 0.12 | 86.6 (±19.2) | 2.3 |
| Provincial | 643.7 | 153.4 (±19.9) | 9.9 | 7.3 (±1.8) | 0.47 | 126.7 (±43.3) | 8.2 | |
| Unprotected | 6386.6 | 159.4 (±28.7) | 101.8 | 6.8 (±2.1) | 4.37 | 94.9 (±51.5) | 60.6 | |
| Total | 7292.4 | 158.3(±27.9) | 115.4 | 6.8 (±2.1) | 4.97 | 97.4 (±50.9) | 71.1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez Pastur, G.; Aravena Acuña, M.-C.; Chaves, J.E.; Cellini, J.M.; Silveira, E.M.O.; Rodriguez-Souilla, J.; von Müller, A.; La Manna, L.; Lencinas, M.V.; Peri, P.L. Nitrogenous and Phosphorus Soil Contents in Tierra del Fuego Forests: Relationships with Soil Organic Carbon, Climate, Vegetation and Landscape Metrics. Land 2023, 12, 983. https://doi.org/10.3390/land12050983
Martínez Pastur G, Aravena Acuña M-C, Chaves JE, Cellini JM, Silveira EMO, Rodriguez-Souilla J, von Müller A, La Manna L, Lencinas MV, Peri PL. Nitrogenous and Phosphorus Soil Contents in Tierra del Fuego Forests: Relationships with Soil Organic Carbon, Climate, Vegetation and Landscape Metrics. Land. 2023; 12(5):983. https://doi.org/10.3390/land12050983
Chicago/Turabian StyleMartínez Pastur, Guillermo, Marie-Claire Aravena Acuña, Jimena E. Chaves, Juan M. Cellini, Eduarda M. O. Silveira, Julián Rodriguez-Souilla, Axel von Müller, Ludmila La Manna, María V. Lencinas, and Pablo L. Peri. 2023. "Nitrogenous and Phosphorus Soil Contents in Tierra del Fuego Forests: Relationships with Soil Organic Carbon, Climate, Vegetation and Landscape Metrics" Land 12, no. 5: 983. https://doi.org/10.3390/land12050983
APA StyleMartínez Pastur, G., Aravena Acuña, M.-C., Chaves, J. E., Cellini, J. M., Silveira, E. M. O., Rodriguez-Souilla, J., von Müller, A., La Manna, L., Lencinas, M. V., & Peri, P. L. (2023). Nitrogenous and Phosphorus Soil Contents in Tierra del Fuego Forests: Relationships with Soil Organic Carbon, Climate, Vegetation and Landscape Metrics. Land, 12(5), 983. https://doi.org/10.3390/land12050983