
The Contribution of Cultivated and Semi-Natural Patches to the Beta Diversity of Nocturnal Lepidoptera within an Organic Century-Old Olive Grove in a Fragmented Landscape

Abstract
1. Introduction
2. Materials and Methods
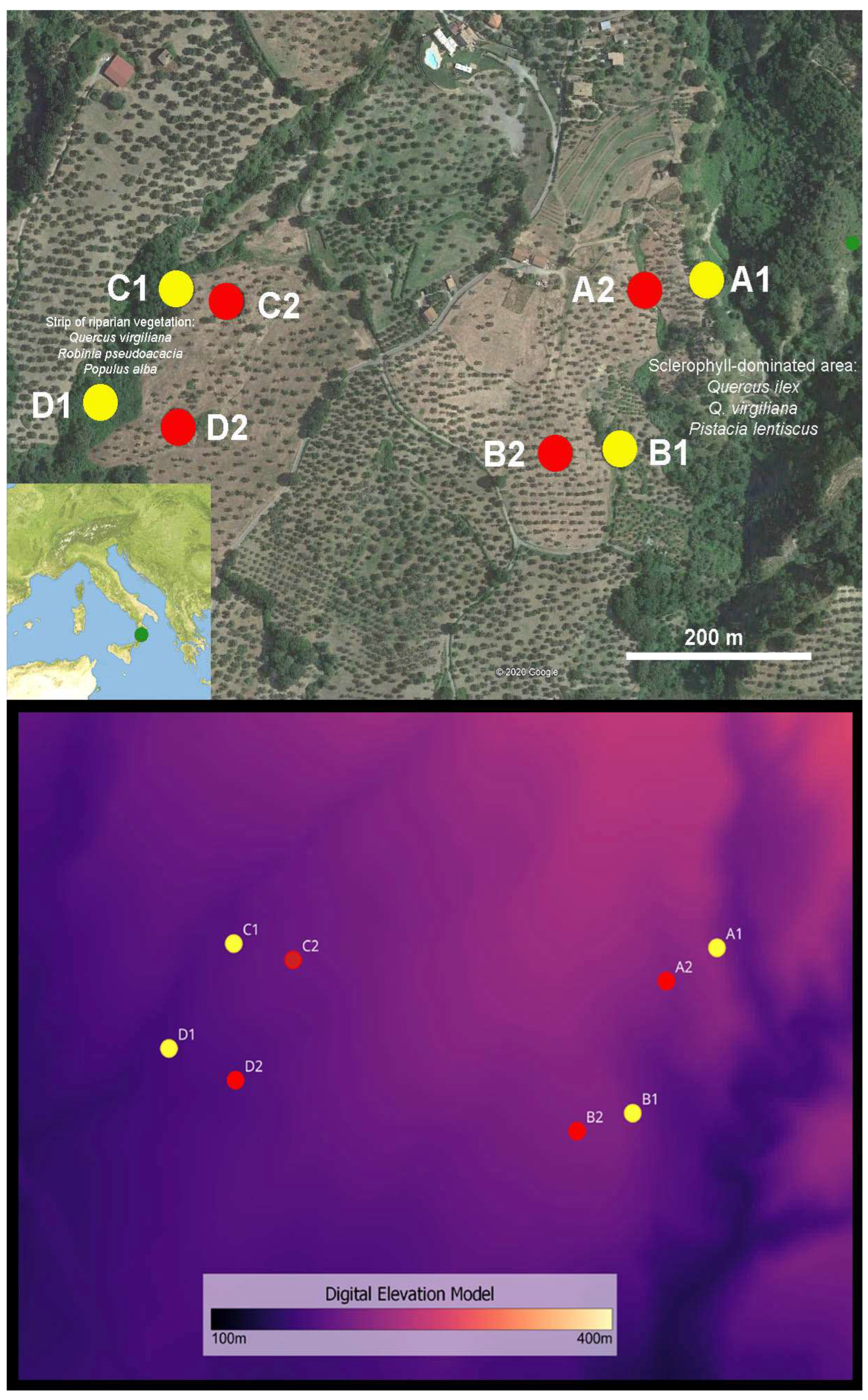
2.1. Study Area
2.2. Moth Dataset
2.3. Statistical Analysis
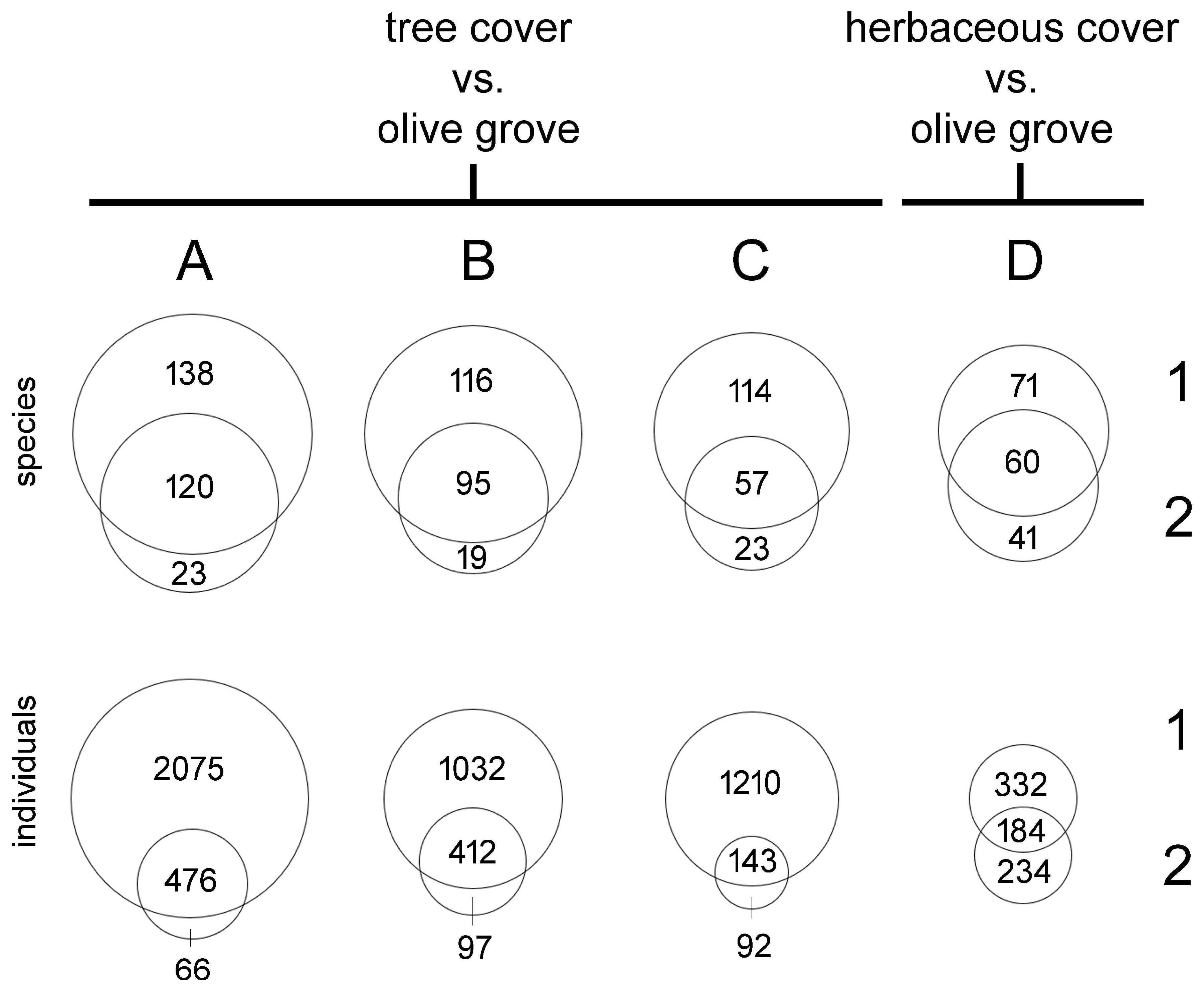
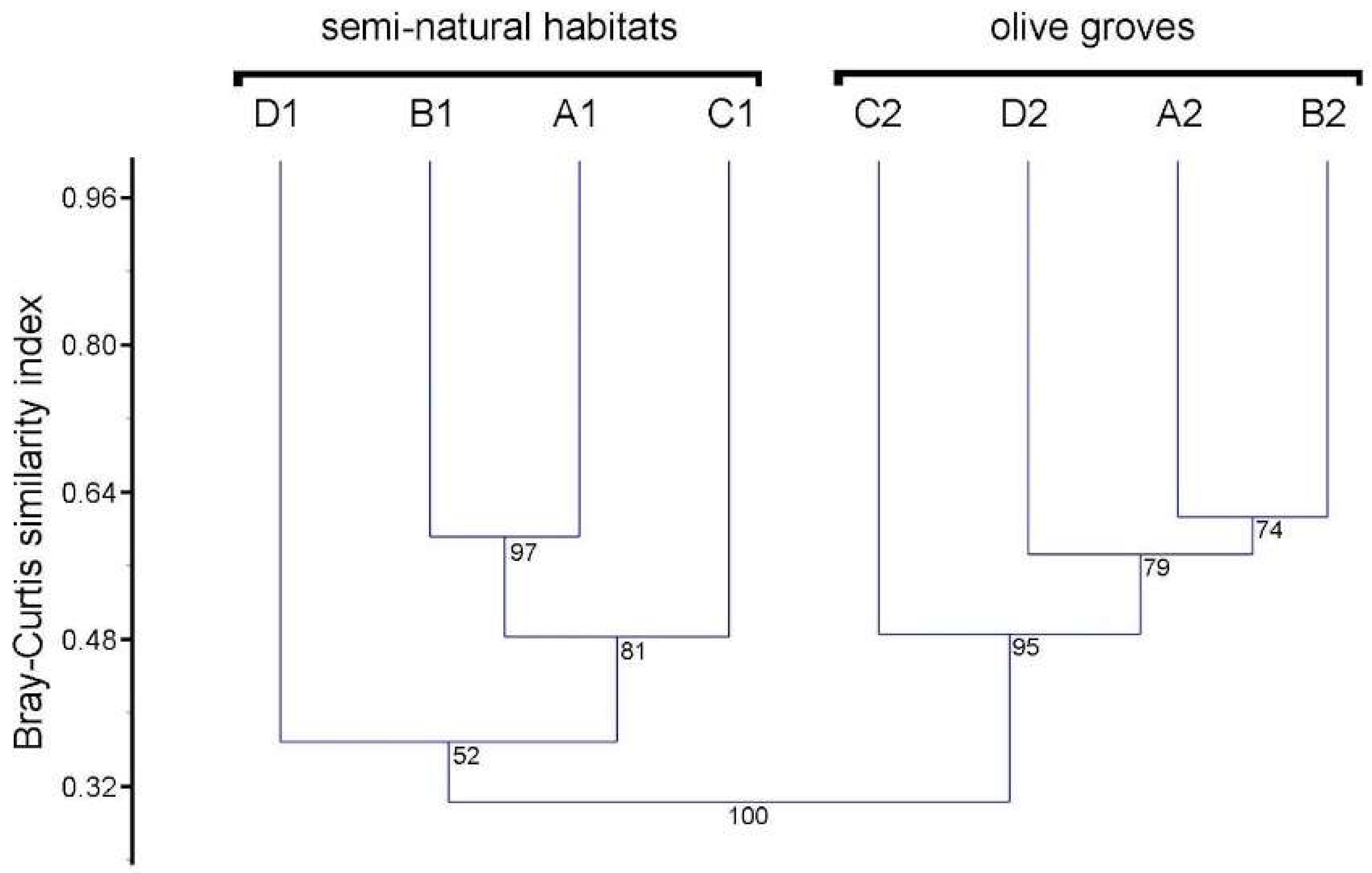
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kazemi, H.; Klug, H.; Kamkara, B. New services and roles of biodiversity in modern agroecosystems: A review. Ecol. Indic. 2018, 93, 1126–1135. [Google Scholar] [CrossRef]
- Swift, M.J. Biodiversity and ecosystem function in agroecosystems. In Biodiversity and Ecosystem Function; Schultze, E., Mooney, H.A., Eds.; Spinger: New York, NY, USA, 1993; pp. 57–83. [Google Scholar]
- Taylor, M.E.; Morecroft, M.D. Effects of agri-environment schemes in a long-term ecological time series. Agric. Ecosyst. Environ. 2009, 130, 9–15. [Google Scholar] [CrossRef]
- Jonason, D.; Franzén, M.; Pettersson, L. Transient peak in moth diversity as a response to organic farming. Basic Appl. Ecol. 2013, 14, 515–522. [Google Scholar] [CrossRef]
- Redon, M.; Berges, L.; Cordonnier, T.; Luque, S. Effects of increasing landscape heterogeneity on local plant species richness: How much is enough? Landsc. Ecol. 2014, 29, 773–787. [Google Scholar] [CrossRef]
- Weibull, A.C.; Bengtsson, J.; Nohlgren, E. Diversity of butterflies in the agricultural landscape: The role of farming system and landscape heterogeneity. Ecography 2000, 23, 743–750. [Google Scholar] [CrossRef]
- Steckel, J.; Westphal, C.; Peters, M.K.; Bellach, M.; Rothenwoehrer, C.; Erasmi, S.; Scherber, C.; Tscharntke, T.; Steffan-Dewenter, I. Landscape composition and configuration differently affect trap-nesting bees, wasps and their antagonists. Biol. Conserv. 2014, 172, 56–64. [Google Scholar] [CrossRef]
- Clough, Y.; Kruess, A.; Kleijn, D.; Tscharntke, T. Spider diversity in cereal fields: Comparing factors at local, landscape and regional scales. J. Biogeogr. 2005, 32, 2007–2014. [Google Scholar] [CrossRef]
- Miyashita, T.; Chishiki, Y.; Takagi, S.R. Landscape heterogeneity at multiple spatial scales enhances spider species richness in an agricultural landscape. Popul. Ecol. 2012, 54, 573–581. [Google Scholar] [CrossRef]
- Ke, A.; Sibiya, M.D.; Reynolds, C.; McCleery, R.A.; Monadjem, A.; Fletcher, R.J. Landscape heterogeneity shapes taxonomic diversity of non-breeding birds across fragmented savanna landscapes. Biodivers. Conserv. 2018, 27, 2681–2698. [Google Scholar] [CrossRef]
- Serafini, V.N.; Priotto, J.W.; Gomez, M.D. Effects of agroecosystem landscape complexity on small mammals: A multi-species approach at different spatial scales. Landsc. Ecol. 2019, 34, 1117–1129. [Google Scholar] [CrossRef]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef] [PubMed]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C. A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Scalercio, S.; Iannotta, N.; Brandmayr, P. The role of semi-natural and abandoned vegetation patches in sustaining lepidopteran diversity in an organic olive orchard. Bull. Insectol. 2007, 60, 13–22. [Google Scholar]
- Öckinger, E.; Smith, H.G. Do corridors promote dispersal in grassland butterflies and other insects? Landsc. Ecol. 2008, 23, 27–40. [Google Scholar] [CrossRef]
- Riva, F.; Acorn, J.H.; Nielsen, S.E. Localized disturbances from oil sands developments increase butterfly diversity and abundance in Alberta’s boreal forests. Biol. Conserv. 2018, 217, 173–180. [Google Scholar] [CrossRef]
- Piccini, I.; Depetris, M.; Paradiso, F.; Cochis, F.; Audisio, M.; Artioli, P.; Smargiassi, S.; Bonifacino, M.; Giuliano, D.; La Cava, S.; et al. Macro-moth (Lepidoptera) Diversity of a Newly Shaped Ecological Corridor and the Surrounding Forest Area in the Western Italian Alps. Diversity 2023, 15, 95. [Google Scholar] [CrossRef]
- Hole, D.G.; Perkins, A.J.; Wilson, J.D.; Alexander, I.H.; Grice, P.V.; Evans, A.D. Does organic farming benefit biodiversity? Biol. Conserv. 2005, 122, 113–130. [Google Scholar] [CrossRef]
- Chiverton, P.A.; Sotherton, N.W. The effects on beneficial arthropods of the exclusion of herbicides from cereal crop edges. J. Appl. Ecol. 1991, 28, 10271039. [Google Scholar] [CrossRef]
- Holzschuh, A.; Steffan-Dewenter, I.; Tscharntke, T. Agricultural landscapes with organic crops support higher pollinator diversity. Oikos 2008, 117, 354–361. [Google Scholar] [CrossRef]
- Bartual, A.M.; Sutter, L.; Bocci, G.; Moonen, A.C.; Cresswell, J.; Entling, M.; Giffard, B.; Jacot, K.; Jeanneret, P.; Hollans, J.; et al. The potential of different semi-natural habitats to sustain pollinators and natural enemies in European agricultural landscapes. Agric. Ecosyst. Environ. 2019, 279, 43–52. [Google Scholar] [CrossRef]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 280–338. [Google Scholar] [CrossRef]
- de Miranda, M.D.; Pereira, H.M.; Corley, M.F.; Merckx, T. Beta diversity patterns reveal positive effects of farmland abandonment on moth communities. Sci. Rep. 2019, 9, 1549. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; Tartaglini, N.; Ladisa, G. Study on Biodiversity in Century-Old Olive Groves; CIHEAM-Mediterranean Agronomic Institute: Bari, Italy, 2012; pp. 1–108. [Google Scholar]
- Morettini, A. Olivi memorabili. L’Italia Agric. 1963, 5, 423–435. [Google Scholar]
- Tartaglini, N.; Calabrese, G.; Servadei, L. Ancient olive orchards as high nature value farmland: A shared vision at Euro-Mediterranean level. In A Multi-Scale and Multi-Level Approach for Conservation of Ancient Olive Orchards in the Euro-Mediterranean Region; La Posta, A., Lacirignola, C., Mimiola, G., Eds.; IAMB: Bari, Italy, 2012; Volume 2, pp. 27–39. [Google Scholar]
- Rader, R.; Bartomeus, I.; Garibaldi, L.A.; Garratt, M.P.D.; Howlett, B.G.; Winfree, R.; Cunningham, S.A.; Mayfield, M.M.; Arthur, A.D.; Andersson, G.K.S.; et al. Non-bee insects are important contributors to global crop pollination. Proc. Natl. Acad. Sci. USA 2016, 113, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Summerville, K.S.; Crist, T.O. Contrasting effects of habitat quantity and quality on moth communities in fragmented landscapes. Ecography 2004, 27, 3–12. [Google Scholar] [CrossRef]
- Summerville, K.S.; Conoan, C.J.; Steichen, R.M. Species traits as predictors of lepidopteran composition in restored and remnant tallgrass prairies. Ecol. Appl. 2006, 16, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Ienco, A.; Dapporto, L.; Greco, S.; Infusino, M.; Scalercio, S. β-diversity partitioning of moth communities within and between different forest types. Sci. Nat. 2020, 107, 8. [Google Scholar] [CrossRef] [PubMed]
- Fayle, T.M.; Sharp, R.E.; Majerus, M.E. The effect of moth trap type on catch size and composition in British Lepidoptera. Br. J. Entomol. Nat. Hist. 2007, 20, 221–232. [Google Scholar]
- Infusino, M.; Brehm, G.; Di Marco, C.; Scalercio, S. Assessing the efficiency of UV LEDs as light sources for sampling the diversity of macro-moths (Lepidoptera). Eur. J. Entomol. 2017, 114, 25–33. [Google Scholar] [CrossRef]
- Wickramasinghe, L.P.; Harris, S.; Jones, G.; Vaughan Jennings, N. Abundance and species richness of nocturnal insects on organic and conventional farms: Effects of agricultural intensification on bat foraging. Conserv. Biol. 2004, 18, 1283–1292. [Google Scholar] [CrossRef]
- Sabatino, M.; Pecoraro, M.; Scalercio, S. I macrolepidotteri notturni di un oliveto a conduzione biologica della Stretta di Catanzaro, Italia meridionale (Insecta Lepidoptera). Mem. Soc. Entomol. Ital. 2021, 98, 3–21. [Google Scholar] [CrossRef]
- Scalercio, S.; Zucco, G. Moths in olive groves. Mendeley Data 2024. V1. [Google Scholar] [CrossRef]
- Prozorov, A.M.; Prozorova, T.A.; Volkova, J.S.; Yakovlev, R.V.; Nedoshivina, S.V.; Pinzari, M.; Scalercio, S.; Bianco, G.; Saldaitis, A.; Hausmann, A.; et al. Revision of the Lemonia taraxaci complex, with a description of a new species from Italy and clarification of the status of Lemonia strigata (Lepidoptera: Brahmaeidae: Lemoniinae). Zootaxa 2022, 5195, 337–360. [Google Scholar] [CrossRef] [PubMed]
- Govi, G.; Fiumi, G.; Barbut, J.; Scalercio, S.; Hausmann, A. An unexpected species complex unveiled in southern European populations of Phragmatiphila nexa (Hübner, 1808.) (Lepidoptera, Noctuidae, Noctuinae, Apameini). Zootaxa 2022, 5128, 355–383. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A. Past: Paleontological statistics software package for educaton and data anlysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science: Oxford, UK, 2004. [Google Scholar]
- Saraçli, S.; Doğan, N.; Doğan, İ. Comparison of hierarchical cluster analysis methods by cophenetic correlation. J. Inequal. Appl. 2013, 2013, 203. [Google Scholar] [CrossRef]
- Strona, G.; Galli, P.; Seveso, D.; Montano, S.; Fattorini, S. Nestedness for Dummies (NeD): A user-friendly web interface for exploratory nestedness analysis. J. Stat. Softw. 2014, 59, 1–9. [Google Scholar] [CrossRef]
- Ulrich, W.; Gotelli, N.J. Null model analysis of species nestedness patterns. Ecology 2007, 88, 1824–1831. [Google Scholar] [CrossRef]
- Ulrich, W.; Almeida-Neto, M.; Gotelli, N.G. A consumer’s guide to nestedness analysis. Oikos 2009, 118, 3–17. [Google Scholar] [CrossRef]
- Almeida-Neto, M.; Guimaraes, P.; Guimaraes, P.R., Jr.; Loyola, R.D.; Ulrich, W. A consistent metric for nestedness analysis in ecological systems: Reconciling concept and measurement. Oikos 2008, 117, 1227–1239. [Google Scholar] [CrossRef]
- Rico-Gray, V.; Díaz-Castelazo, C.; Ramírez-Hernández, A.; Guimarães, P.R.; Nathaniel Holland, J. Abiotic factors shape temporal variation in the structure of an ant–plant network. Arthropod-Plant Interact. 2012, 6, 289–295. [Google Scholar] [CrossRef]
- Zucco, G.; Scalercio, S. Establishment of a long-term monitoring program of moths in a suburban area of south Italy: First results of the years 2019–2021 (Insecta Lepidoptera). Redia 2023, 106, 27–40. [Google Scholar] [CrossRef]
- Scalercio, S. Nocturnal macrolepidoptera associated to cork oak woodlands and neighbour forest remnants in a fragmented Mediterranean landscape (Lepidoptera). Redia 2022, 105, 131–139. [Google Scholar] [CrossRef]
- Lindsay, K.E. Farmland heterogeneity benefits birds in American mid-west watersheds. Am. Midl. Nat. 2013, 170, 121–143. [Google Scholar] [CrossRef]
- Regolin, A.L.; Ribeiro, M.C.; Martello, F.; Melo, G.L.; Sponchiado, J.; Campanha, L.F.D.C.; Sugai, L.S.M.; Silva, T.S.F.; Caceres, N.C. Spatial heterogeneity and habitat configuration overcome habitat composition influences on alpha and beta mammal diversity. Biotropica 2020, 52, 969–980. [Google Scholar] [CrossRef]
- Warzecha, D.; Diekötter, T.; Wolters, V.; Jauker, F. Spatial configuration and landscape context of wildflower areas determine their benefits to pollinator α-and β-diversity. Basic Appl. Ecol. 2021, 56, 335–344. [Google Scholar] [CrossRef]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How should beta-diversity inform biodiversity conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Lagerlöf, J.; Wallin, H. The abundance of arthropods along two field margins with different types of vegetation composition: An experimental study. Agric. Ecosyst. Environ. 1993, 43, 141–154. [Google Scholar] [CrossRef]
- Álvarez, H.A.; Morente, M.; Oi, F.S.; Rodríguez, E.; Campos, M.; Ruano, F. Semi-natural habitat complexity affects abundance and movement of natural enemies in organic olive orchards. Agric. Ecosyst. Environ. 2019, 285, 106618. [Google Scholar] [CrossRef]
- Scalercio, S.; Brandmayr, P.; Iannotta, N.; Petacchi, R.; Boccaccio, L. Correlations between landscape attributes and ecological traits of Lepidoptera communities in olive groves. Eur. J. Entomol. 2012, 109, 207–216. [Google Scholar] [CrossRef]
- Marasas, M.E.; Sarandón, S.J.; Cicchino, A. Semi-natural habitats and field margins in a typical agroecosystem of the Argentinean Pampas as a reservoir of carabid beetles. J. Sustain. Agric. 2010, 34, 153–168. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nicholls, C.I.; Ponti, L. Biodiversità e Controllo dei Fitofagi Negli Agroecosistemi, 1st ed.; Accademia Nazionale Italiana di Entomologia: Firenze, Italy, 2003. [Google Scholar]
- McLaughlin, A.; Mineau, P. The impact of agricultural practices on biodiversity. Agric. Ecosyst. Environ. 1995, 55, 201–212. [Google Scholar] [CrossRef]
- Chen, L.; Fu, B.; Zhao, W. Source-sink landscape theory and its ecological significance. Front. Biol. China 2008, 3, 131–136. [Google Scholar] [CrossRef]
- Barraquand, F.; Benhamou, S. Animal movements in heterogeneous landscapes: Identifying profitable places and homogeneous movement bouts. Ecology 2008, 89, 3336–3348. [Google Scholar] [CrossRef]
- Brühl, C.A.; Zaller, J.G. Indirect herbicide effects on biodiversity, ecosystem functions, and interactions with global changes. In Herbicides; Mesnage, R., Zaller, J.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 231–272. [Google Scholar]
- Nicholls, C.I.; Altieri, M.A. Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef]
- Reddy, N.V.; Dubey, N.; Avinashe, H. Destruction of Paddy-by-Paddy Swarming Caterpillar (Spodopteramauritia) in Assam. Int. J. Innov. Res. Tech. 2020, 7, 207–210. [Google Scholar]
- Johansen, L.; Westin, A.; Wehn, S.; Iuga, A.; Ivascu, C.M.; Kallioniemi, E.; Lennartsson, T. Traditional semi-natural grassland management with heterogeneous mowing times enhances flower resources for pollinators in agricultural landscapes. Glob. Ecol. Conserv. 2019, 18, e00619. [Google Scholar] [CrossRef]
- Zaller, J.G.; Kruse-Plaß, M.; Schlechtriemen, U.; Gruber, E.; Peer, M.; Nadeem, I.; Formayer, H.; Hutter, H.P.; Landler, L. Pesticides in ambient air, influenced by surrounding land use and weather, pose a potential threat to biodiversity and humans. Sci. Total Environ. 2022, 838, 156012. [Google Scholar] [CrossRef]
- Usher, M.B.; Keiller, S.W. The macrolepidoptera of farm woodlands: Determinants of diversity and community structure. Biodiver. Conserv. 1998, 7, 725–748. [Google Scholar] [CrossRef]
- Mohamad, R.S.; Bteich, M.R.; Cardone, G.; Marchini, A. Economic analysis in organic olive farms: The case of the ancient olive trees in the rural parkland in Apulia. New Medit 2013, 12, 55–61. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Individuals |
|---|---|---|
| Noctuidae | 135 | 2727 |
| Geometridae | 108 | 2719 |
| Erebidae | 52 | 1871 |
| Nolidae | 12 | 140 |
| Notodontidae | 8 | 25 |
| Lasiocampidae | 5 | 8 |
| Cossidae | 2 | 2 |
| Sphingidae | 1 | 8 |
| Limacodidae | 1 | 6 |
| Euteliidae | 1 | 5 |
| Brahmaeidae | 1 | 1 |
| Saturnidae | 1 | 1 |
| Individuals | Semi-Natural | Olive Grove | |
|---|---|---|---|
| (N) | (%) | (%) | |
| Acontia trabealis (Scopoli, 1763) | 180 | 17.2 | 82.8 |
| Tyta luctuosa (Denis & Schiffermüller, 1775) | 91 | 26.4 | 73.6 |
| Apochima flabellaria (Heeger, 1838) | 27 | 29.6 | 70.4 |
| Agrotis lata (Treitschke, 1835) | 13 | 15.4 | 84.6 |
| Heliothis peltigera (Denis & Schiffermüller, 1775) | 13 | 15.4 | 84.6 |
| Sesamia nonagrioides (Lefèbvre, 1827) | 5 | 0 | 100 |
| Samples | Statistics | Values | Z-Scores | Nested | p |
|---|---|---|---|---|---|
| Farm | NODF | 67.55 | 19.28 | yes | p < 0.001 |
| MT | 20.25 | −14.35 | yes | p < 0.001 | |
| pair A | NODF | 49.11 | 8.61 | yes | p < 0.001 |
| MT | 10.10 | −8.64 | yes | p < 0.001 | |
| pair B | NODF | 48.91 | 17.45 | yes | p < 0.001 |
| MT | 8.46 | −8.31 | yes | p < 0.001 | |
| pair C | NODF | 42.26 | 11.08 | yes | p < 0.001 |
| MT | 15.57 | −7.08 | yes | p < 0.001 | |
| pair D | NODF | 46.17 | 0.32 | no | N.S. |
| MT | 26.21 | −1.75 | yes | p < 0.05 |
| Samples | Statistics | Value | Z-Score | Nested | p |
|---|---|---|---|---|---|
| semi-natural patches | NODF | 66.81 | 9.77 | yes | p < 0.001 |
| MT | 17.02 | −11.24 | yes | p < 0.001 | |
| olive groves | NODF | 59.95 | 6.46 | yes | p < 0.001 |
| MT | 26.26 | −7.98 | yes | p < 0.001 | |
| dry farm | NODF | 67.69 | 11.60 | yes | p < 0.001 |
| MT | 14.31 | −11.17 | yes | p < 0.001 | |
| wet farm | NODF | 58.54 | 9.82 | yes | p < 0.001 |
| MT | 25.44 | −9.285 | yes | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zucco, G.; La Cava, S.; Rijllo, G.; Scalercio, S. The Contribution of Cultivated and Semi-Natural Patches to the Beta Diversity of Nocturnal Lepidoptera within an Organic Century-Old Olive Grove in a Fragmented Landscape. Land 2024, 13, 532. https://doi.org/10.3390/land13040532
Zucco G, La Cava S, Rijllo G, Scalercio S. The Contribution of Cultivated and Semi-Natural Patches to the Beta Diversity of Nocturnal Lepidoptera within an Organic Century-Old Olive Grove in a Fragmented Landscape. Land. 2024; 13(4):532. https://doi.org/10.3390/land13040532
Chicago/Turabian StyleZucco, Giada, Sara La Cava, Giuseppe Rijllo, and Stefano Scalercio. 2024. "The Contribution of Cultivated and Semi-Natural Patches to the Beta Diversity of Nocturnal Lepidoptera within an Organic Century-Old Olive Grove in a Fragmented Landscape" Land 13, no. 4: 532. https://doi.org/10.3390/land13040532
APA StyleZucco, G., La Cava, S., Rijllo, G., & Scalercio, S. (2024). The Contribution of Cultivated and Semi-Natural Patches to the Beta Diversity of Nocturnal Lepidoptera within an Organic Century-Old Olive Grove in a Fragmented Landscape. Land, 13(4), 532. https://doi.org/10.3390/land13040532