
The Effect of Climate on Strongly Disturbed Vegetation of Bait Sites in a Central European Lower Montane Zone, Hungary

Abstract
:1. Introduction
- How does the extent of weed coverage change over the years and whether seasonal changes are detected?
- How does precipitation and temperature affect the number and the coverage of weed species in each year?
2. Materials and Methods
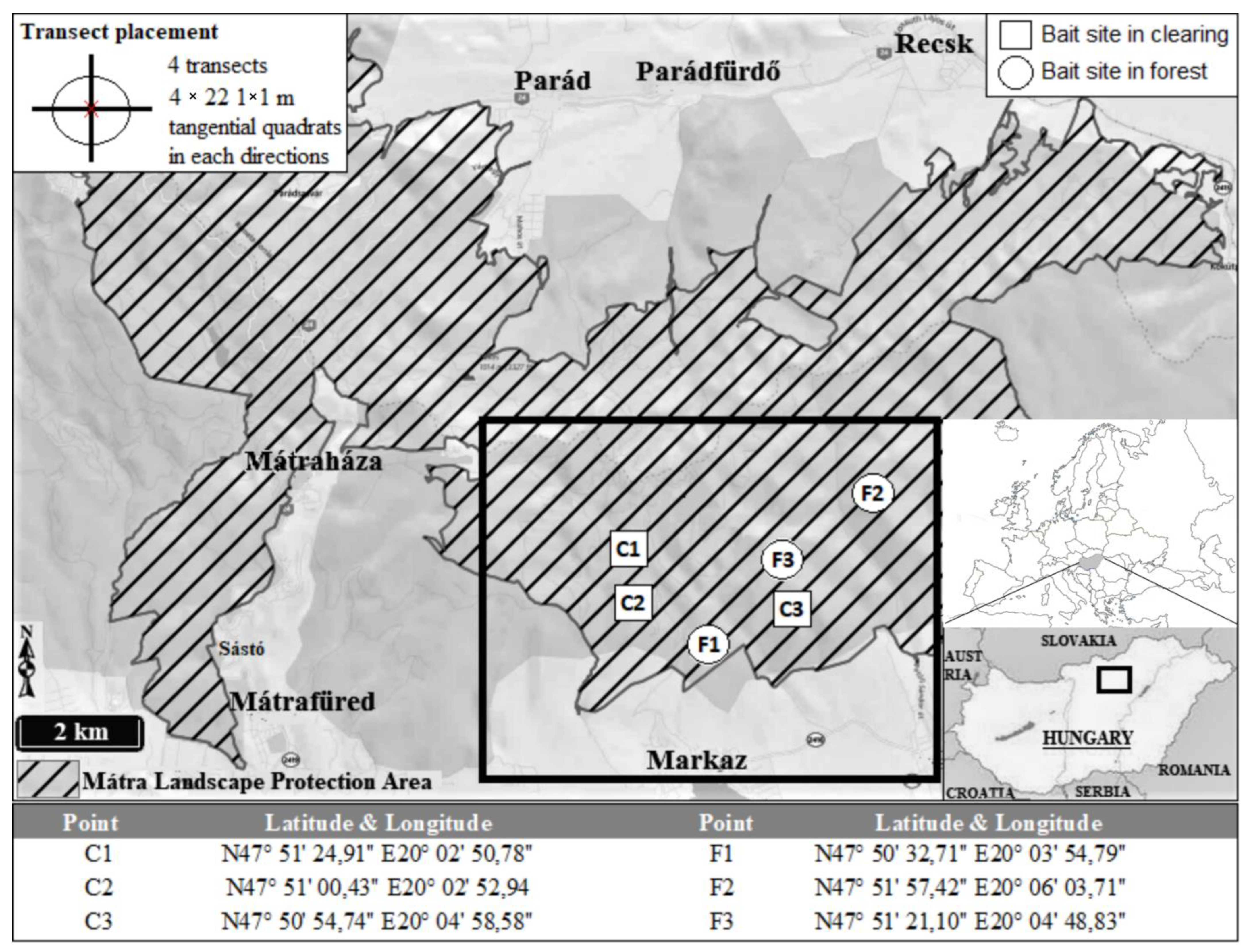
2.1. Description of the Study Area
2.2. Coenosystematic Classification
2.3. Statistical Analysis
3. Results
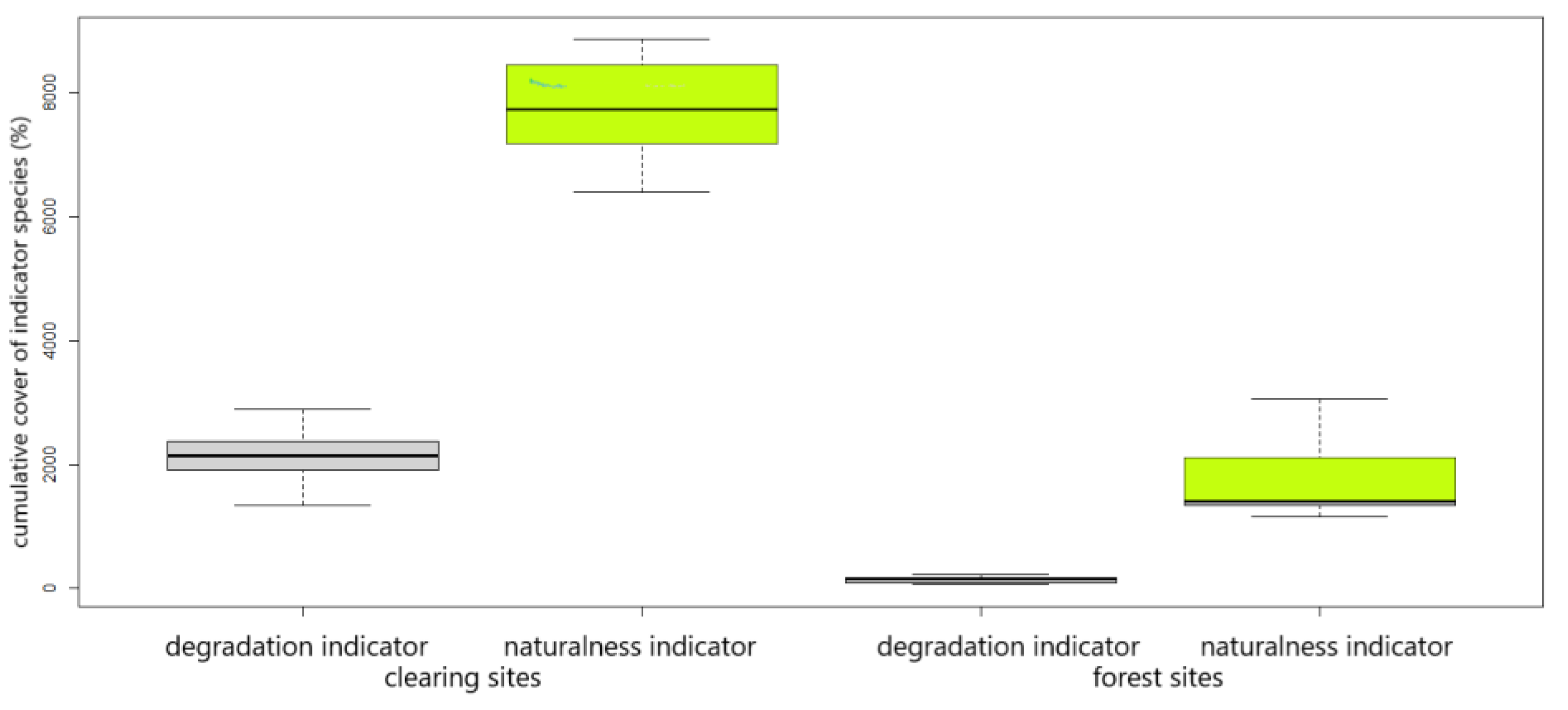
3.1. Species Pool and Abundance
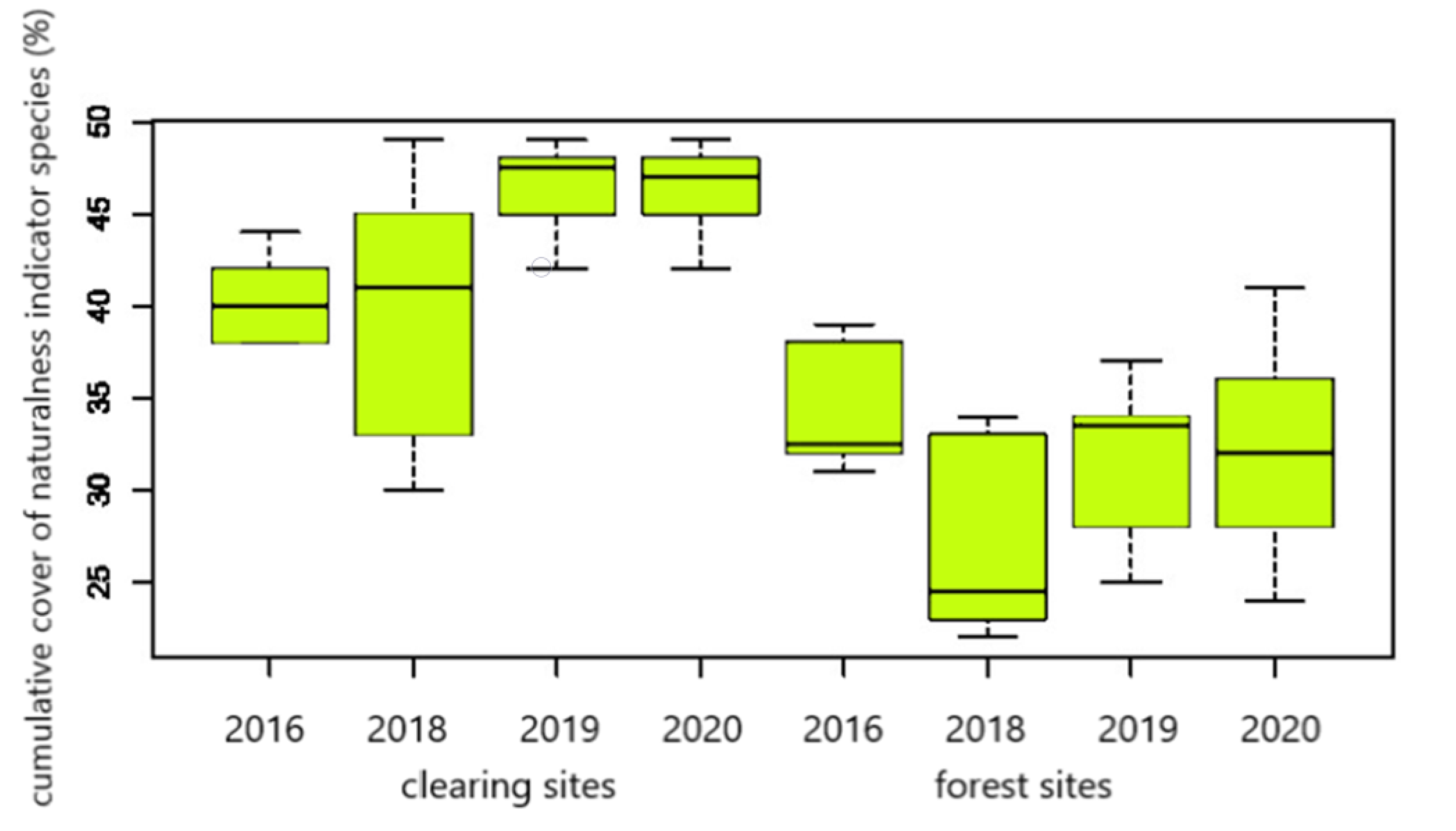
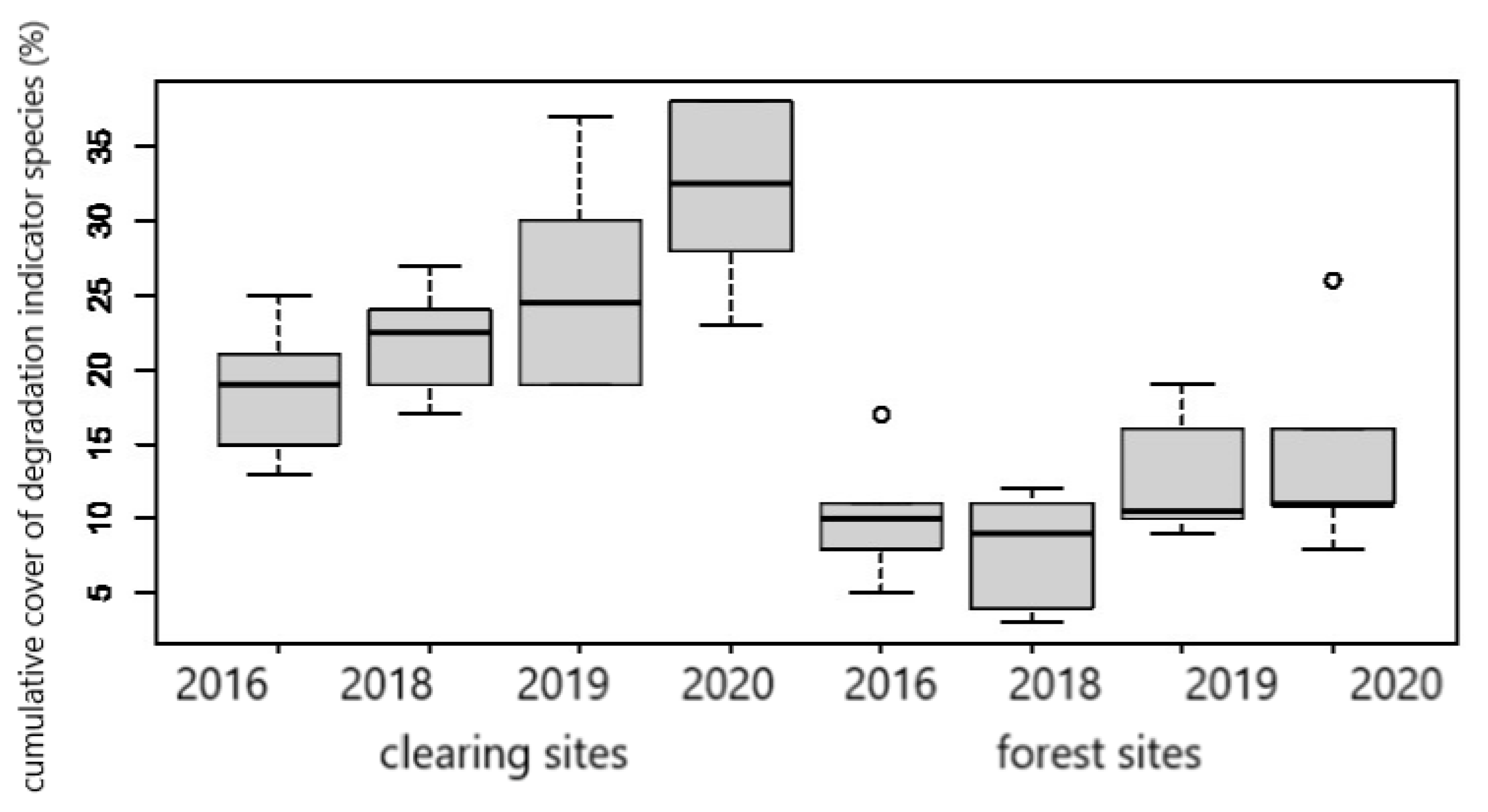
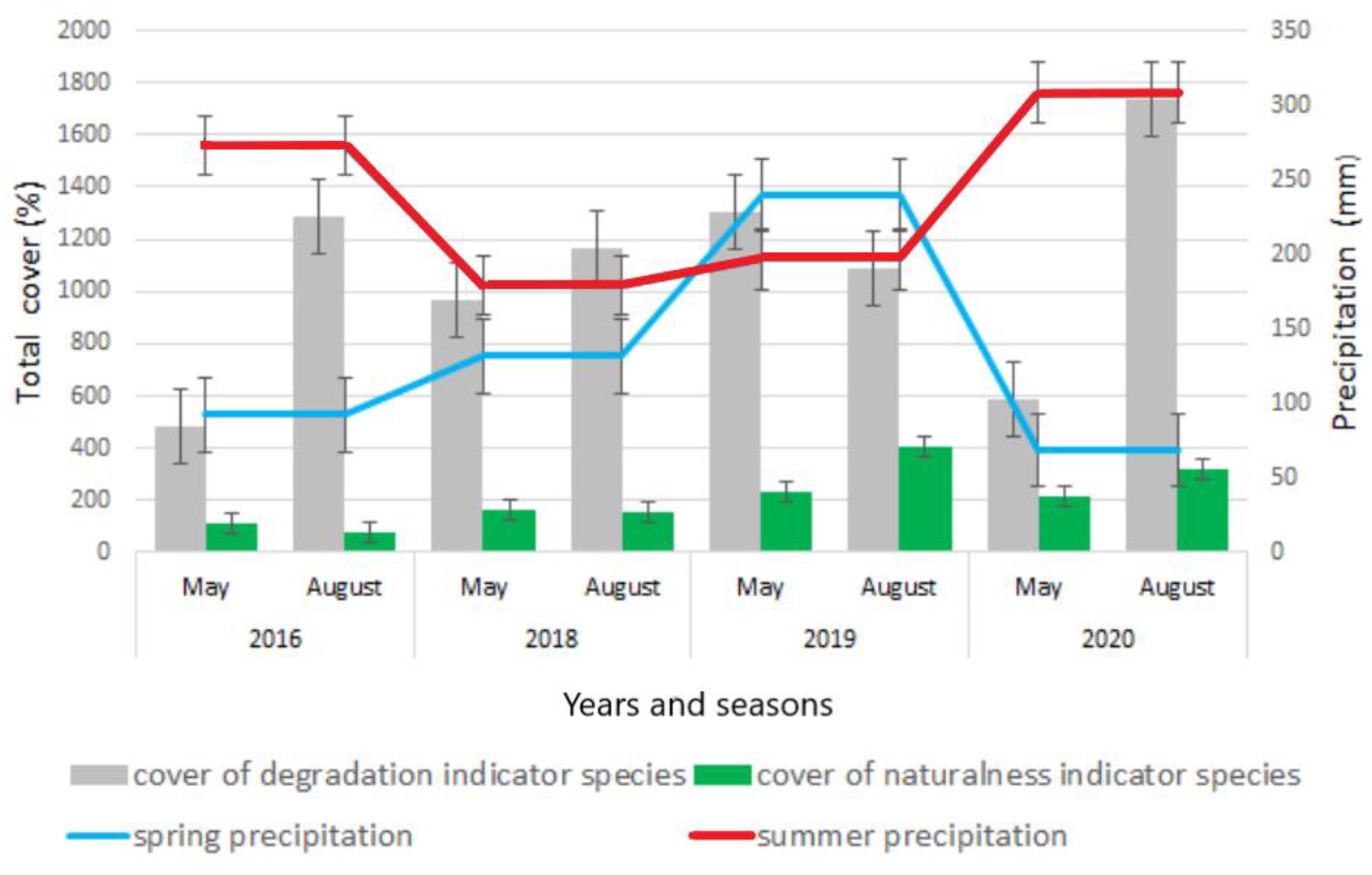
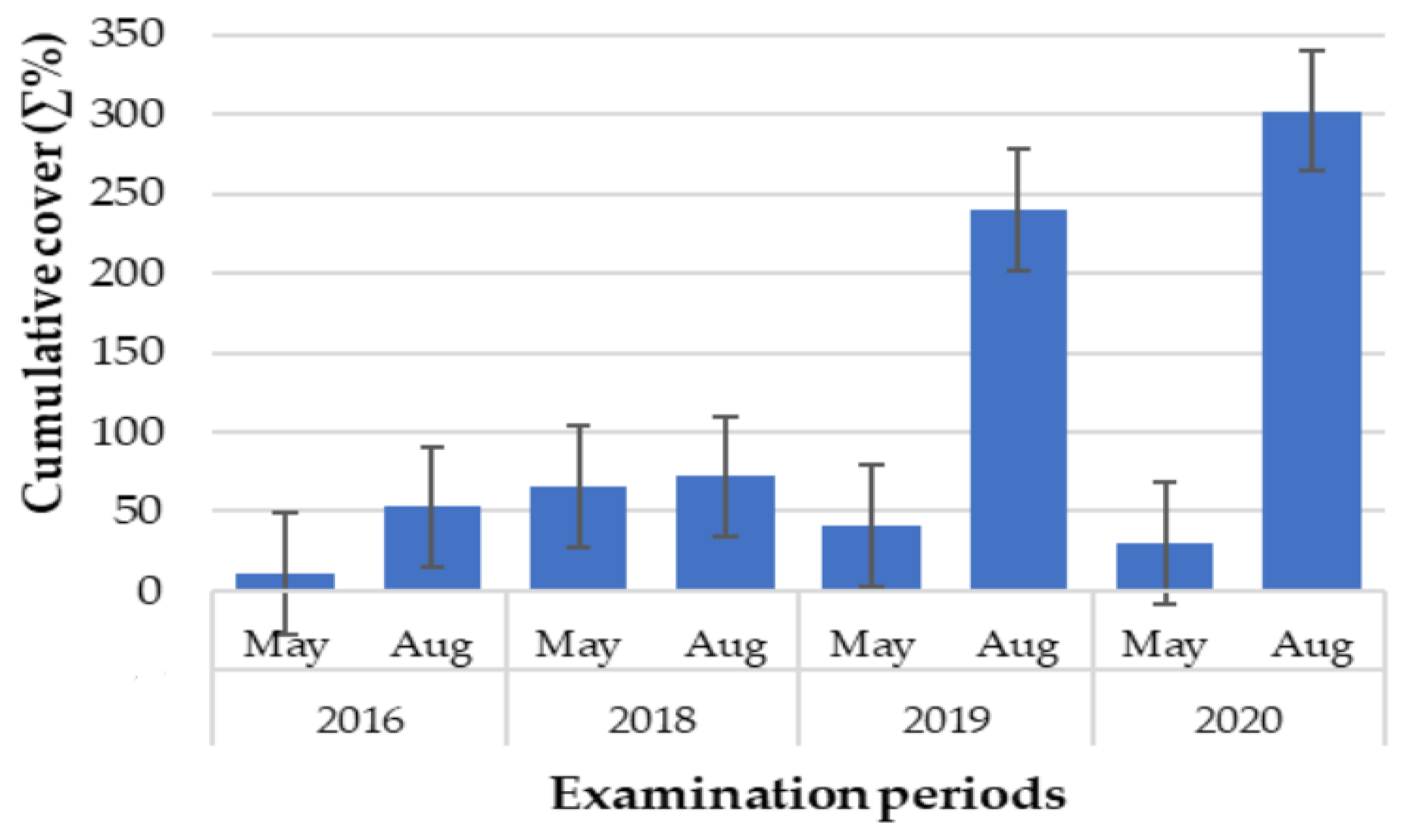
3.2. Intra- and Interannual Changes in Vegetation
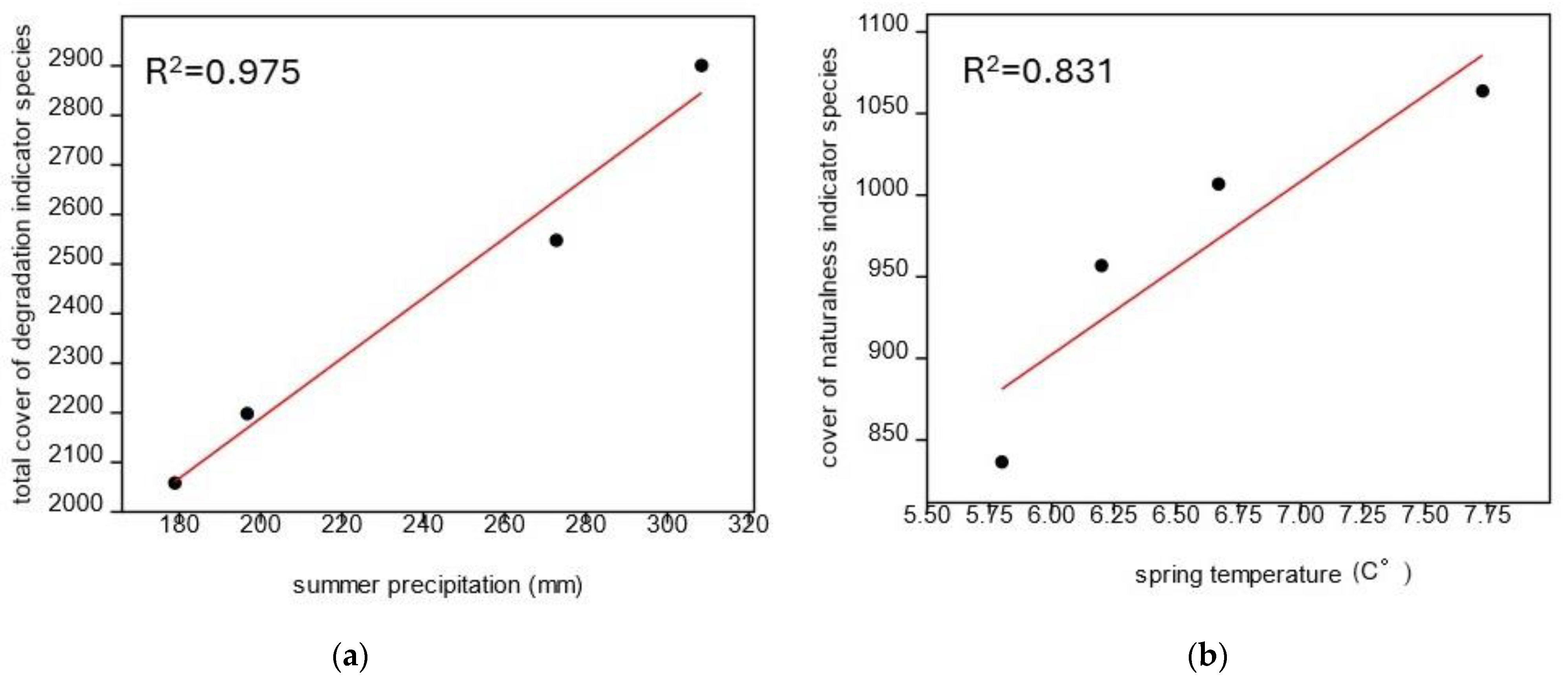
3.3. Influence of Meteorological Factors on Vegetation
4. Discussion
4.1. Species Pool and Abundance
4.2. Intra- and Interannual Changes in Vegetation
4.3. The Role of Meteorological Factors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selva, N.; Berezowska-Cnota, T.; Elguero-Claramunt, I. Unforeseen Effects of Supplementary Feeding: Ungulate Baiting Sites as Hotspots for Ground-Nest Predation. PLoS ONE 2014, 9, e90740. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C. Supplemental feeding of deer in west texas. Trans-pecos wildlife management series. Leaflet 2006, 9, 10. [Google Scholar]
- Milner, J.M.; Van Beest, F.M.; Schmidt, K.T.; Brook, R.K.; Storaas, T. To feed or not to feed? Evidence of the intended and unintended effects of feeding wild ungulates. J. Wildl. Manag. 2014, 78, 1322–1334. [Google Scholar] [CrossRef]
- Sorensen, A.; van Beest, F.M.; Brook, R.K. Impacts of wildlife baiting and supplemental feeding on infectious disease transmission risk: A synthesis of knowledge. Prev. Vet. Med. 2014, 113, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Inslerman, R.A.; Baker, D.L.; Cumberland, R.; Doerr, P.; Miller, J.E.; Kennamer, J.E.; Stinson, E.R.; Williamson, S.J. Baiting and Supplemental Feeding of Game Wildlife Species; Technical Review 06-1; The Wildlife Society: Washington, DC, USA, 2006. [Google Scholar]
- Apollonio, M.; Andersen, R.; Putman, R. European Ungulates and Their Management in the 21st Century; Cambridge University Press: New York, NY, USA, 2010; 618p. [Google Scholar]
- Wilson, C.E.; Castro, K.L.; Thurston, G.B.; Sissons, A. Pathway risk analysis of weed seeds in imported grain: A Canadian perspective. NeoBiota 2016, 30, 49–74. [Google Scholar]
- Gervilla, C.; Rita, J.; Cursach, J. Contaminant seeds in imported crop seed lots: A non-negligible human-mediated pathway for introduction of plant species to islands. Weed Res. 2019, 59, 245–253. [Google Scholar] [CrossRef]
- Hobbs, R.; Huenneke, L. Disturbance, diversity, and invasion: Implications for conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef]
- OVA National Wildlife Management Database [Országos Vadgazdálkodási Adattár]. 2023. Available online: http://www.ova.info.hu/ (accessed on 3 June 2024).
- Rusvai, K.; Saláta, D.; Falvai, D.; Czóbel, S. Assesment of weed invasion at bait sites in a Central European lower montane zone. Perspect. Plant Ecol. Evol. Syst. 2022, 55, 125667. [Google Scholar] [CrossRef]
- Rusvai, K.; Wichmann, B.; Saláta, D.; Grónás, V.; Skutai, J.; Czóbel, S. Changes in the Vegetation, Soil Seed Bank and Soil Properties at Bait Sites in a Protected Area of the Central European Lower Montane Zone. Sustainability 2022, 14, 13134. [Google Scholar] [CrossRef]
- Kochjarová, J.; Blanár, D.; Jarolímek, I.; Slezák, M. Wildlife supplementary feeding facilitates spread of alien plants in forested mountainous areas: A case study from the Western Carpathians. Biologia 2023, 2, 1381–1399. [Google Scholar] [CrossRef]
- Rusvai, K.; Czóbel, S. Changes in Soil Seed Bank and Vegetation at Abandoned Bait Sites in a Central European Hilly Area. Biol. Life Sci. Forum 2021, 2, 15. [Google Scholar] [CrossRef]
- Kochjarová, J.; Blanár, D. Anthropophytes in protected mountain area: Past, present and risk to diversity—A case study from the Muránska planina National park (Western Carpathians). Acta Oecol. Carp. 2018, 11, 75–90. [Google Scholar]
- Pinke, G.; Karácsony, P.; Czúcz, B.; Botta-Dukát, Z.; Lengyel, A. The influence of environment, management and site context on species composition of summer arable weed vegetation in Hungary. Appl. Veg. Sci. 2012, 15, 136–144. [Google Scholar] [CrossRef]
- HungaroMet. 2020. Available online: https://odp.met.hu/ (accessed on 20 June 2021).
- Borhidi, A. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian Flora. Acta Bot. Hung. 1995, 39, 97–181. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Zeigerwerte von Pflanzen in Mitteleuropa. Scr. Geobot. 1991, 18, 1–248. [Google Scholar]
- Grime, J.P. Plant Strategies and Vegetation Processes; John Wiley: Chichester, NY, USA, 1979. [Google Scholar]
- Király, G. (Ed.) Új Magyar Füvészkönyv. Magyarország Hajtásos Növényei. Határozókulcsok [New Hungarian Herbal. The Vascular Plants of Hungary. Identification Key]; ANP Igazgatóság: Jósvafő, Hungary, 2009; pp. 3–456.
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 20 October 2023).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1. Available online: https://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 20 February 2023).
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Hejda, M.; Arianoutsou, M.; Essl, F.; Jarošík, V.; Pergl, J.; Winter, M.; Anastasiu, P.; et al. Alien flora of Europe: Species diversity, temporal trends, geographical patterns and research needs. Preslia 2008, 80, 101–149. [Google Scholar]
- Pauchard, A.; Kueffer, C.; Dietz, H.; Daehler, C.C.; Alexander, J.; Edwards, P.J.; Arévalo, J.R.; Cavieres, L.; Guisan, A.; Haider, S.; et al. Ain’t no mountain high enough: Plant invasions reaching new elevations. Front. Ecol. Environ. 2009, 7, 479–486. [Google Scholar] [CrossRef]
- Pinke, G.; Pál, R. The Origin, Growing Place and Protection of Our Weeds; Alexandra Kiadó: Pécs, Hungary, 2005; 232p. (In Hungarian) [Google Scholar]
- Von Holle, B. Environmental stress alters native-nonnative relationships at the community scale. Biol. Invasions 2013, 15, 417–427. [Google Scholar] [CrossRef]
- Bíró, I. Nature conservation aspects of hunting and wildlife management in Békés County. Puszta 1998, 1, 73–96. (In Hungarian) [Google Scholar]
- Molnár, V.A. Conservation Botany; Teaching aids for the University of Debrecen course; Debreceni Egyetem TTK Növénytani Tanszék: Debrecen, Hungary, 2014; 65p. (In Hungarian) [Google Scholar]
- Berzsenyi, Z. Weeds, Weed Control, Weed Biology; Mezőgazda Kiadó: Budapest, Hungary, 2000; 347p. (In Hungarian) [Google Scholar]
- Pinke, G.; Pál, R.; Botta-Dukát, Z. Effects of environmental factors on weed species composition of cereal and stubble fields in western Hungary. Cent. Eur. J. Biol. 2010, 5, 283–292. [Google Scholar] [CrossRef]
- Sauter-Louis, C.; Conraths, F.J.; Probst, C.; Blohm, U.; Schulz, K.; Sehl, J.; Fischer, M.; Forth, J.H.; Zani, L.; Depner, K.; et al. African Swine Fever in Wild Boar in Europe—A Review. Viruses 2021, 13, 1717. [Google Scholar] [CrossRef] [PubMed]
- Szabó, G. (Ed.) “Subjective Experience Geography” Study Volume in Honor of Professor Antal Aubert; Pécsi Tudományegyetem, Természettudományi Kar, Földrajzi és Földtudományi Intézet: Pécs, Hungary, 2021; 247p. (In Hungarian) [Google Scholar]
- Jánoska, F. Environmental impact assessments in wild boar gardens. Gyepgazdálkodási Közlemények 2006, 4, 82–85. (In Hungarian) [Google Scholar] [CrossRef]
- Lehoczky, É.; Kamuti, M.; Mazsu, N.; Sándor, R. Changes to soil water content and biomass yield under combined maize and maize-weed vegetation with different fertilization treatments in loam soil. J. Hydrol. Hydromech. 2016, 64, 150–159. [Google Scholar] [CrossRef]
- Wainwright, C.E.; Cleland, E.E. Exotic species display greater germination plasticity and higher germination rates than native species across multiple cues. Biol. Invasions 2013, 15, 2253–2264. [Google Scholar] [CrossRef]
- Hulme, P.E. Climate change and biological invasions: Evidence, expectations, and response options. Biol. Rev. 2016, 92, 1297–1313. [Google Scholar] [CrossRef] [PubMed]
- Hirka, A.; Csóka, G. Annual ragweed (Ambrosia artemisiifolia L.) in Hungarian forests. Növényvédelem 2009, 45, 438–439. (In Hungarian) [Google Scholar]
- Rusvai, K. The Effects of Feeding Places for Hunting Purposes on Vegetation, Seed Bank And Soil in the Mátra Mountains. Doctoral Dissertation, Hungarian University of Agriculture and Life Sciences, Ph.D. School of Environmental Sciences, Gödöllő, Hungary, 2023; 189p. [Google Scholar]
- Milad, M.; Schaich, H.; Konold, W. How is adaptation to climate change reflected in current practice of forest management and conservation? A case study from Germany. Biodivers. Conserv. 2011, 22, 1181–1202. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flanningan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate Change and Forest Disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Kueffer, C.; McDougall, K.; Alexander, J.; Daehler, C.; Edwards, P.; Haider, S.; Milbau, A.; Parks, C.; Pauchard, A.; Reshi, Z.A.; et al. Plant Invasions into Mountain Protected Areas: Assessment, Prevention and Control at Multiple Spatial Scales. Plant Invasions in Protected Areas. In Plant Invasions in Protected Areas. Patterns, Problems and Challenges; Foxcroft, L.C., Pyšek, P., Richardson, D.M., Genovesi, P., Eds.; Invading Nature–Springer Series in Invasion Ecology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 7, pp. 89–113. [Google Scholar]
- Laurence, W.F.; Yensen, E. Predicting the impacts of edge effects in fragmented habitats. Biol. Conserv. 1991, 55, 77–92. [Google Scholar] [CrossRef]
- Martin, P.H.; Canham, C.D.; Marks, P.L. Why forests appear resistant to exotic plant invasions: Intentional introductions, stand dynamics, and the role of shade tolerance. Front. Ecol. Environ. 2009, 7, 142–149. [Google Scholar] [CrossRef]
- Rejmánek, M.; Richardson, D.M.; Pyšek, P. Plant Invasions and Invasibility of Plant Communities. In Vegetation Ecology, 2nd ed.; Van der Maarel, E., Franklin, J., Eds.; Wiley-Blackwell: Oxford, UK, 2013; pp. 387–424. [Google Scholar]
- Blossey, B.; Gorchov, D.L. Introduction to the special issue: Ungulates and invasive species: Quantifying impacts and understanding interactions. AoB Plants 2017, 9, plx063. [Google Scholar] [CrossRef] [PubMed]
- Burst, M.; Chauchard, S.; Dupouey, J.-L.; Amiaud, B. Interactive effects of land-use change and distance-to-edge on the distribution of species in plant communities at the forest-grassland interface. J. Veg. Sci. 2017, 28, 515–526. [Google Scholar] [CrossRef]
- Kleijn, D.; Sutherland, W.J. How effective are European agrienvironment schemes in conserving and promoting biodiversity? J. Appl. Ecol. 2003, 40, 947–969. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Naturalness Indicator Species | Indicator | Degradation Indicator Species | Indicator |
|---|---|---|---|
| Urtica dioica L. | DT | Rumex crispus L. | W |
| Carex divulsa Stokes. | DT | Fallopia convolvulus (L.) Á. Löve | W |
| Poa nemoralis L. | C | Chenopodium album L. | RC |
| Geum urbanum L. | DT | Plantago major L. | W |
| Moehringia trinervia (L.) Clairv. | DT | Polygonum aviculare L. | RC |
| Poa angustifolia L. | DT | Galium aparine L. | W |
| Festuca heterophylla Lam. | C | Ambrosia artemisiifolia L. | AC |
| Melica uniflora Retz. | C | Veronica hederifolia L. | W |
| Lysimachia nummularia L. | DT | Datura stramonium L. | W |
| Dactylis glomerata L. | DT | Convolvulus arvensis L. | RC |
| Naturalness Indicator Species | Indicator | Degradation Indicator Species | Indicator |
|---|---|---|---|
| Poa angustifolia L. | DT | Polygonum aviculare L. | RC |
| Carex praecox Schreb. | G | Xanthium spinosum L. | W |
| Fragaria viridis Duch. | G | Tripleurospermum inodorum (L.) Sch.Bip | W |
| Festuca rubra L. | C | Bromus sterilis L. | RC |
| Achillea collina J. Beck | DT | Datura stramonium L. | W |
| Bromus hordeaceus L. | DT | Capsella bursa-pastoris (L.) Medik | W |
| Euphorbia cyparissias L. | DT | Convolvulus arvensis L. | RC |
| Festuca pseudovina Hack. ex Wiesb. | C | Ballota nigra L. | W |
| Urtica dioica L. | DT | Chenopodium album L. | RC |
| Galium verum L. | DT | Rumex crispus L. | W |
| 2016 | 2018 | 2019 | 2020 | |
|---|---|---|---|---|
| Alien species | +790% | +46% | +514% | +1763% |
| Native weeds | +3% | +1% | −10% | +78% |
| Total weed coverage | +21% | +18% | +0.4% | +116% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusvai, K.; Házi, J.; Czóbel, S. The Effect of Climate on Strongly Disturbed Vegetation of Bait Sites in a Central European Lower Montane Zone, Hungary. Land 2024, 13, 1108. https://doi.org/10.3390/land13071108
Rusvai K, Házi J, Czóbel S. The Effect of Climate on Strongly Disturbed Vegetation of Bait Sites in a Central European Lower Montane Zone, Hungary. Land. 2024; 13(7):1108. https://doi.org/10.3390/land13071108
Chicago/Turabian StyleRusvai, Katalin, Judit Házi, and Szilárd Czóbel. 2024. "The Effect of Climate on Strongly Disturbed Vegetation of Bait Sites in a Central European Lower Montane Zone, Hungary" Land 13, no. 7: 1108. https://doi.org/10.3390/land13071108