

Bird Diversity in Suburban Greenway Was Driven by Habitat Heterogeneity and Landscape Patterns in Autumn–Winter Seasons—Evidence from Hangzhou Qingshan Lake Greenway

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experiment Design
2.3. Bird Survey
2.4. Landscape Parameter Acquisition
2.5. Data Analysis
3. Results
3.1. Overall Distribution of Bird Communities
3.2. Bird Diversity in Green Corridor Distribution in Different Habitat Types
3.2.1. Alpha Diversity
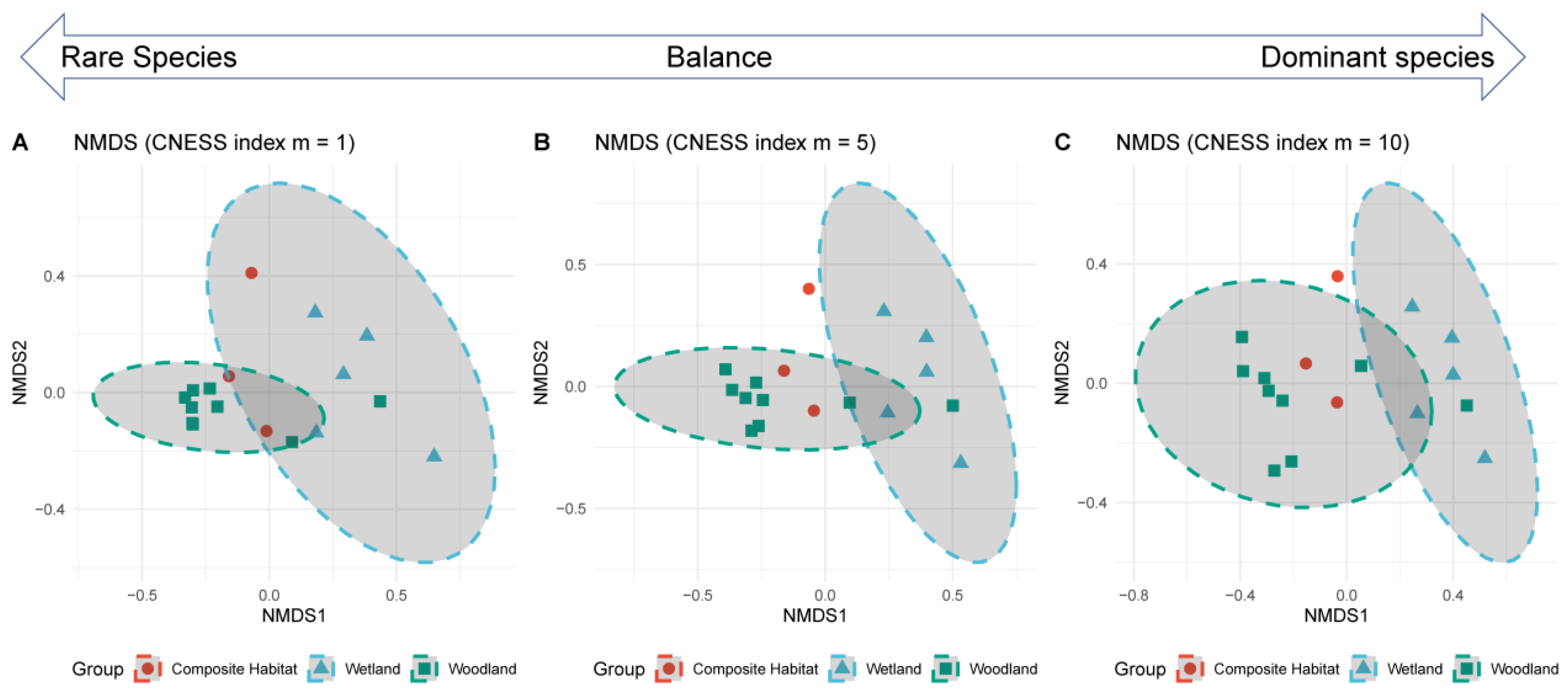
3.2.2. Beta Diversity
3.3. Bird Diversity in Green Corridor Response to Landscape Pattern
3.3.1. Diversity Response to Landscape Configuration
3.3.2. Diversity Response to Landscape Composition
4. Discussion
4.1. Bird Diversity Distribution in Urban Greenways
4.2. Impact of Landscape Patterns on Bird Diversity in Greenways
4.3. Implications for Future Urban Greenway Construction
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, X.; Xie, Y.; Qi, K.; Luo, Z.; Wang, X. Detecting the response of bird communities and biodiversity to habitat loss and fragmentation due to urbanization. Sci. Total Environ. 2018, 624, 1561–1576. [Google Scholar] [CrossRef] [PubMed]
- Reis, E.; López-Iborra, G.M.; Pinheiro, R.T. Changes in bird species richness through different levels of urbanization: Implications for biodiversity conservation and garden design in Central Brazil. Landsc. Urban Plan. 2012, 107, 31–42. [Google Scholar] [CrossRef]
- Chen, R.; Carruthers-Jones, J.; Carver, S.; Wu, J. Constructing urban ecological corridors to reflect local species diversity and conservation objectives. Sci. Total Environ. 2024, 907, 167987. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J.; Cox, D.T.; Canavelli, S.B.; García, D.; Hughes, B.; Maas, B.; Martínez, D.; Ogada, D.; Inger, R. Population abundance and ecosystem service provision: The case of birds. Bioscience 2018, 68, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- Wood, E.; Harsant, A.; Dallimer, M.; Cronin De Chavez, A.; McEachan, R.R.; Hassall, C. Not all green space is created equal: Biodiversity predicts psychological restorative benefits from urban green space. Front. Psychol. 2018, 9, 2320. [Google Scholar] [CrossRef]
- Kong, F.; Yin, H.; Nakagoshi, N.; Zong, Y. Urban green space network development for biodiversity conservation: Identification based on graph theory and gravity modeling. Landsc. Urban Plan. 2010, 95, 16–27. [Google Scholar] [CrossRef]
- Angold, P.G.; Sadler, J.P.; Hill, M.O.; Pullin, A.; Rushton, S.; Austin, K.; Small, E.; Wood, B.; Wadsworth, R.; Sanderson, R. Biodiversity in urban habitat patches. Sci. Total Environ. 2006, 360, 196–204. [Google Scholar] [CrossRef]
- Horte, O.S.; Eisenman, T.S. Urban greenways: A systematic review and typology. Land 2020, 9, 40. [Google Scholar] [CrossRef]
- Chin, E.Y.; Kupfer, J.A. Identification of environmental drivers in urban greenway communities. Urban For. Urban Green. 2020, 47, 126549. [Google Scholar] [CrossRef]
- Lynch, A.J. Creating effective urban greenways and stepping-stones: Four critical gaps in habitat connectivity planning research. J. Plan. Lit. 2019, 34, 131–155. [Google Scholar] [CrossRef]
- Klingbeil, B.T.; Willig, M.R. Community assembly in temperate forest birds: Habitat filtering, interspecific interactions and priority effects. Evol. Ecol. 2016, 30, 703–722. [Google Scholar]
- Carlier, J.; Moran, J. Landscape typology and ecological connectivity assessment to inform Greenway design. Sci. Total Environ. 2019, 651, 3241–3252. [Google Scholar] [CrossRef] [PubMed]
- Bueno, J.A.; Tsihrintzis, V.A.; Alvarez, L. South Florida greenways: A conceptual framework for the ecological reconnectivity of the region. Landsc. Urban Plan. 1995, 33, 247–266. [Google Scholar] [CrossRef]
- Buelow, C.; Sheaves, M. A birds-eye view of biological connectivity in mangrove systems. Estuar. Coast. Shelf Sci. 2015, 152, 33–43. [Google Scholar] [CrossRef]
- Von Haaren, C.; Reich, M. The German way to greenways and habitat networks. Landsc. Urban Plan. 2006, 76, 7–22. [Google Scholar]
- Thiele, T.; Jeltsch, F.; Blaum, N. Importance of woody vegetation for foraging site selection in the Southern Pied Babbler (Turdoides bicolor) under two different land use regimes. J. Arid. Environ. 2008, 72, 471–482. [Google Scholar] [CrossRef]
- Moudrý, V.; Moudrá, L.; Barták, V.; Bejček, V.; Gdulová, K.; Hendrychová, M.; Moravec, D.; Musil, P.; Rocchini, D.; Šťastný, K. The role of the vegetation structure, primary productivity and senescence derived from airborne LiDAR and hyperspectral data for birds diversity and rarity on a restored site. Landsc. Urban Plan. 2021, 210, 104064. [Google Scholar] [CrossRef]
- Xi, C.; Chi, Y.; Qian, T.; Zhang, W.; Wang, J. Simulation of Human Activity Intensity and Its Influence on Mammal Diversity in Sanjiangyuan National Park, China. Sustainability 2020, 12, 4601. [Google Scholar] [CrossRef]
- Semenchuk, P.; Plutzar, C.; Kastner, T.; Matej, S.; Bidoglio, G.; Erb, K.; Essl, F.; Haberl, H.; Wessely, J.; Krausmann, F. Relative effects of land conversion and land-use intensity on terrestrial vertebrate diversity. Nat. Commun. 2022, 13, 615. [Google Scholar] [CrossRef]
- Rich, A.C.; Dobkin, D.S.; Niles, L.J. Defining forest fragmentation by corridor width: The influence of narrow forest-dividing corridors on forest-nesting birds in southern New Jersey. Conserv. Biol. 1994, 8, 1109–1121. [Google Scholar] [CrossRef]
- Laiolo, P. Spatial and seasonal patterns of bird communities in Italian agroecosystems. Conserv. Biol. 2005, 19, 1547–1556. [Google Scholar] [CrossRef]
- Tzortzakaki, O.; Kati, V.; Kassara, C.; Tietze, D.T.; Giokas, S. Seasonal patterns of urban bird diversity in a Mediterranean coastal city: The positive role of open green spaces. Urban Ecosyst. 2018, 21, 27–39. [Google Scholar] [CrossRef]
- St, L.; Wold, S. Analysis of variance (ANOVA). Chemom. Intell. Lab. Syst. 1989, 6, 259–272. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Arsham, H.; Lovric, M. Bartlett’s Test. Int. Encycl. Stat. Sci. 2011, 2, 20–23. [Google Scholar]
- Williams, L.J.; Abdi, H. Fisher’s least significant difference (LSD) test. Encycl. Res. Des. 2010, 218, 840–853. [Google Scholar]
- Zou, Y.; Axmacher, J.C. The Chord-Normalized Expected Species Shared (CNESS)-distance represents a superior measure of species turnover patterns. Methods Ecol. Evol. 2020, 11, 273–280. [Google Scholar] [CrossRef]
- Taguchi, Y.; Oono, Y. Relational patterns of gene expression via non-metric multidimensional scaling analysis. Bioinformatics 2005, 21, 730–740. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational multivariate analysis of variance (PERMANOVA). Wiley Statsref Stat. Ref. Online 2014, 1–15. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- O’Brien, R.M. A caution regarding rules of thumb for variance inflation factors. Qual. Quant. 2007, 41, 673–690. [Google Scholar] [CrossRef]
- Whittingham, M.J.; Stephens, P.A.; Bradbury, R.B.; Freckleton, R.P. Why do we still use stepwise modelling in ecology and behaviour? J. Anim. Ecol. 2006, 75, 1182–1189. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2023. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 321–326. [Google Scholar]
- de Mendiburu, F.; de Mendiburu, M.F. Package ‘agricolae’. R Package Version 2019, 1, 1143–1149. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82. [Google Scholar] [CrossRef]
- Lai, J.; Zou, Y.; Zhang, J.; Peres Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca. hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Peh, K.S.; De Jong, J.; Sodhi, N.S.; Lim, S.L.; Yap, C.A. Lowland rainforest avifauna and human disturbance: Persistence of primary forest birds in selectively logged forests and mixed-rural habitats of southern Peninsular Malaysia. Biol. Conserv. 2005, 123, 489–505. [Google Scholar] [CrossRef]
- Andren, H. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A review. Oikos 1994, 71, 355–366. [Google Scholar] [CrossRef]
- Kang, W.; Minor, E.S.; Park, C.; Lee, D. Effects of habitat structure, human disturbance, and habitat connectivity on urban forest bird communities. Urban Ecosyst. 2015, 18, 857–870. [Google Scholar] [CrossRef]
- Deppe, J.L.; Rotenberry, J.T. Scale-dependent habitat use by fall migratory birds: Vegetation structure, floristics, and geography. Ecol. Monogr. 2008, 78, 461–487. [Google Scholar] [CrossRef]
- Buron, R.; Hostetler, M.E.; Andreu, M. Urban forest fragments vs. residential neighborhoods: Urban habitat preference of migratory birds. Landsc. Urban Plan. 2022, 227, 104538. [Google Scholar] [CrossRef]
- Ramos, D.L.; Pizo, M.A.; Ribeiro, M.C.; Cruz, R.S.; Morales, J.M.; Ovaskainen, O. Forest and connectivity loss drive changes in movement behavior of bird species. Ecography 2020, 43, 1203–1214. [Google Scholar] [CrossRef]
- Eyster, H.N.; Srivastava, D.S.; Kreitzman, M.; Chan, K.M. Functional traits and metacommunity theory reveal that habitat filtering and competition maintain bird diversity in a human shared landscape. Ecography 2022, 2022, e6240. [Google Scholar] [CrossRef]
- Bryant, M.M. Urban landscape conservation and the role of ecological greenways at local and metropolitan scales. Landsc. Urban Plan. 2006, 76, 23–44. [Google Scholar] [CrossRef]
- Vergnes, A.; Le Viol, I.; Clergeau, P. Green corridors in urban landscapes affect the arthropod communities of domestic gardens. Biol. Conserv. 2012, 145, 171–178. [Google Scholar] [CrossRef]
- Beaugeard, E.; Brischoux, F.; Angelier, F. Green infrastructures and ecological corridors shape avian biodiversity in a small French city. Urban Ecosyst. 2021, 24, 549–560. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Fletcher Jr, R.J.; Didham, R.K.; Banks-Leite, C.; Barlow, J.; Ewers, R.M.; Rosindell, J.; Holt, R.D.; Gonzalez, A.; Pardini, R.; Damschen, E.I. Is habitat fragmentation good for biodiversity? Biol. Conserv. 2018, 226, 9–15. [Google Scholar] [CrossRef]
- Nielsen, A.B.; Van Den Bosch, M.; Maruthaveeran, S.; van den Bosch, C.K. Species richness in urban parks and its drivers: A review of empirical evidence. Urban Ecosyst. 2014, 17, 305–327. [Google Scholar] [CrossRef]
- Damschen, E.I.; Baker, D.V.; Bohrer, G.; Nathan, R.; Orrock, J.L.; Turner, J.R.; Brudvig, L.A.; Haddad, N.M.; Levey, D.J.; Tewksbury, J.J. How fragmentation and corridors affect wind dynamics and seed dispersal in open habitats. Proc. Natl. Acad. Sci. USA 2014, 111, 3484–3489. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.F. Habitat corridors and the conservation of small mammals in a fragmented forest environment. Landsc. Ecol. 1990, 4, 109–122. [Google Scholar] [CrossRef]
- Gehring, T.M.; Swihart, R.K. Body size, niche breadth, and ecologically scaled responses to habitat fragmentation: Mammalian predators in an agricultural landscape. Biol. Conserv. 2003, 109, 283–295. [Google Scholar] [CrossRef]
- Rybicki, J.; Abrego, N.; Ovaskainen, O. Habitat fragmentation and species diversity in competitive communities. Ecol. Lett. 2020, 23, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Schütz, C.; Schulze, C.H. Functional diversity of urban bird communities: Effects of landscape composition, green space area and vegetation cover. Ecol. Evol. 2015, 5, 5230–5239. [Google Scholar] [CrossRef] [PubMed]
- Tryjanowski, P.; Morelli, F.; Mikula, P.; Krištín, A.; Indykiewicz, P.; Grzywaczewski, G.; Kronenberg, J.; Jerzak, L. Bird diversity in urban green space: A large-scale analysis of differences between parks and cemeteries in Central Europe. Urban For. Urban Green. 2017, 27, 264–271. [Google Scholar] [CrossRef]
- May, F.; Giladi, I.; Ziv, Y.; Jeltsch, F. Dispersal and diversity–unifying scale-dependent relationships within the neutral theory. Oikos 2012, 121, 942–951. [Google Scholar]
- Zhang, Z.; Meerow, S.; Newell, J.P.; Lindquist, M. Enhancing landscape connectivity through multifunctional green infrastructure corridor modeling and design. Urban For. Urban Green. 2019, 38, 305–317. [Google Scholar]
- Yang, C.; Guo, H.; Huang, X.; Wang, Y.; Li, X.; Cui, X. Ecological network construction of a national park based on MSPA and MCR models: An example of the proposed national parks of “Ailaoshan-Wuliangshan” in China. Land 2022, 11, 1913. [Google Scholar] [CrossRef]
- Beier, P.; Noss, R.F. Do habitat corridors provide connectivity? Conserv. Biol. 1998, 12, 1241–1252. [Google Scholar]
- Haddad, N.M.; Brudvig, L.A.; Damschen, E.I.; Evans, D.M.; Johnson, B.L.; Levey, D.J.; Orrock, J.L.; Resasco, J.; Sullivan, L.L.; Tewksbury, J.J. Potential negative ecological effects of corridors. Conserv. Biol. 2014, 28, 1178–1187. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, M. When corridors work: Insights from a microecosystem. Ecol. Model. 2007, 202, 441–453. [Google Scholar]
- Oliver, T.; Roy, D.B.; Hill, J.K.; Brereton, T.; Thomas, C.D. Heterogeneous landscapes promote population stability. Ecol. Lett. 2010, 13, 473–484. [Google Scholar] [CrossRef]
- Salviano, I.R.; Gardon, F.R.; Dos Santos, R.F. Ecological corridors and landscape planning: A model to select priority areas for connectivity maintenance. Landsc. Ecol. 2021, 36, 3311–3328. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Df | Sum Sq | Mean Sq | F | p | Sig |
|---|---|---|---|---|---|---|
| Species richness | 2 | 1016.1 | 508.1 | 15 | <0.001 | *** |
| Abundance (log-transformed) | 2 | 11.34 | 5.67 | 15.03 | <0.001 | *** |
| Shannon diversity | 2 | 0.596 | 0.2979 | 1.259 | 0.314 | |
| Simpson diversity | 2 | 0.01223 | 0.006117 | 0.289 | 0.753 |
| Model | Variable | Estimate | Std. Error | t/z Value | p |
|---|---|---|---|---|---|
| Species Richness 1 | (Intercept) | 59.69 | 10.59 | 5.636 | <0.001 |
| PARA_SD | −227.4 | 63.26 | −3.595 | 0.0032 | |
| Abundance 1 | (Intercept) | 8.812 | 0.501 | 17.604 | <0.001 |
| DIVISION | −5.283 | 0.278 | −19.024 | <0.001 | |
| PD | 0.008 | 0.001 | 8.182 | <0.001 | |
| PR | 0.041 | 0.006 | 6.606 | <0.001 | |
| SHDI | 1.369 | 0.11 | 12.491 | <0.001 | |
| PARA_SD | −18.56 | 1.454 | −12.763 | <0.001 | |
| Shannon Diversity | (Intercept) | 3.714 | 0.613 | 6.058 | <0.001 |
| PR | −0.065 | 0.026 | −2.521 | 0.0249 | |
| Simpson Diversity | (Intercept) | 1.184 | 0.183 | 6.451 | <0.001 |
| PR | −0.017 | 0.008 | −2.179 | 0.0457 |
| Landscape Configuration Index | Bird Community Composition with CNESS Index m = 1 | Bird Community Composition with CNESS Index m = 5 | Bird Community Composition with CNESS Index m = 10 | |||
|---|---|---|---|---|---|---|
| Explanation Rate (m = 1) | I.perc (%) (m = 1) | Explanation Rate (m = 5) | I.perc (%) (m = 5) | Explanation Rate (m = 10) | I.perc (%) (m = 10) | |
| AI | −0.0081 | −6.33 | −0.0046 | −3.51 | −0.0008 | −0.65 |
| DIVISION | 0.0113 | 8.83 | 0.0079 | 6.03 | 0.0028 | 2.28 |
| ED | −0.0086 | −6.72 | −0.0056 | −4.27 | −0.0022 | −1.79 |
| PARA_SD | 0.0616 | 48.12 | 0.0585 | 44.66 | 0.0478 | 38.86 |
| PD | 0.0134 | 10.47 | 0.0166 | 12.67 | 0.0184 | 14.96 |
| PR | 0.0279 | 21.8 | 0.0235 | 17.94 | 0.0193 | 15.69 |
| SHDI | 0.0171 | 13.36 | 0.0187 | 14.27 | 0.0195 | 15.85 |
| SHEI | 0.0129 | 10.08 | 0.0162 | 12.37 | 0.0179 | 14.55 |
| Model | Variable | Estimate | Std. Error | t/z Value | p |
|---|---|---|---|---|---|
| Species Richness 1 | (Intercept) | 14.744 | 5.481 | 2.69 | 0.041 |
| Park and Green Space | 60.187 | 21.309 | 2.824 | 0.0156 | |
| Public Facility Land | 434.045 | 197.718 | 2.195 | 0.0494 | |
| Facility Agricultural Land | −152.728 | 100.708 | −1.517 | 0.1561 | |
| Abundance 1 | (Intercept) | −28 | 130.3 | −0.215 | 0.835323 |
| Rural Road | −32,449.6 | 3644.7 | −8.903 | <0.001 | |
| Park and Green Space | 5765.7 | 806.6 | 7.148 | <0.001 | |
| Canal | 10,903.1 | 1919.1 | 5.681 | <0.001 | |
| Public Facility Land | 18,359.3 | 7515.5 | 2.443 | 0.040384 | |
| Grassland | −18,408.1 | 4574.8 | −4.024 | 0.004 | |
| Facility Agricultural Land | −10,102 | 2608.8 | −3.872 | 0.004726 | |
| Orchard | −4796 | 2047.5 | −2.342 | 0.04724 | |
| Bare Rock | 7402.4 | 1727.3 | 4.286 | 0.002667 | |
| Shannon Diversity 2 | - | ||||
| Simpson Diversity 2 | - | ||||
| Landscape Configuration Index | Bird Community Composition with CNESS Index m = 1 | Bird Community Composition with CNESS Index m = 5 | Bird Community Composition with CNESS Index m = 10 | |||
|---|---|---|---|---|---|---|
| Explanation Rate (m = 1) | I.perc (%) (m = 1) | Explanation Rate (m = 5) | I.perc (%) (m = 5) | Explanation Rate (m = 10) | I.perc (%) (m = 10) | |
| Rural roads | 0.0172 | 4.62 | 0.0137 | 4.58 | 0.0113 | 5.18 |
| Parks and green spaces | 0.0391 | 10.51 | 0.0256 | 8.56 | 0.0137 | 6.28 |
| Ditches | 0.0137 | 3.68 | 0.0048 | 1.61 | −0.0036 | −1.65 |
| Urban residential land | 0.0056 | 1.51 | −0.0036 | −1.2 | −0.0134 | −6.15 |
| Tea plantations | 0.0335 | 9.01 | 0.0234 | 7.83 | 0.0084 | 3.85 |
| Public facilities land | 0.0078 | 2.1 | 0.0026 | 0.87 | −0.0004 | −0.18 |
| Other grasslands | 0.0498 | 13.39 | 0.0346 | 11.57 | 0.022 | 10.09 |
| Facility agricultural land | 0.0787 | 21.16 | 0.0794 | 26.56 | 0.072 | 33.03 |
| Orchards | −0.0003 | −0.08 | −0.012 | −4.01 | −0.0185 | −8.49 |
| Arbor forests | 0.024 | 6.45 | 0.0191 | 6.39 | 0.0127 | 5.83 |
| Reservoir water surface | 0.0022 | 0.59 | −0.0026 | −0.87 | −0.0065 | −2.98 |
| Bare rocky gravel land | −0.0019 | −0.51 | −0.0002 | −0.07 | −0.0007 | −0.32 |
| Adjustable arbor forests | 0.0883 | 23.74 | 0.1047 | 35.02 | 0.1158 | 53.12 |
| Total building land | 0.0145 | 3.9 | 0.0093 | 3.11 | 0.0049 | 2.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, Y.; Hu, W.; Wang, W.; He, L.; Xu, W. Bird Diversity in Suburban Greenway Was Driven by Habitat Heterogeneity and Landscape Patterns in Autumn–Winter Seasons—Evidence from Hangzhou Qingshan Lake Greenway. Land 2024, 13, 1192. https://doi.org/10.3390/land13081192
Tao Y, Hu W, Wang W, He L, Xu W. Bird Diversity in Suburban Greenway Was Driven by Habitat Heterogeneity and Landscape Patterns in Autumn–Winter Seasons—Evidence from Hangzhou Qingshan Lake Greenway. Land. 2024; 13(8):1192. https://doi.org/10.3390/land13081192
Chicago/Turabian StyleTao, Yizhou, Wenhao Hu, Wenjing Wang, Lan He, and Wenhui Xu. 2024. "Bird Diversity in Suburban Greenway Was Driven by Habitat Heterogeneity and Landscape Patterns in Autumn–Winter Seasons—Evidence from Hangzhou Qingshan Lake Greenway" Land 13, no. 8: 1192. https://doi.org/10.3390/land13081192