1. Introduction
Mental rotation is an abstract operation whereby a person imagines rotating an object or a body part so as to place it in a different position [
1]. It is a form of mental imagery [
2], a complex cognitive process having its roots in real perception, through which we recall from memory, generate and manipulate mental representations, in the absence of environmental input [
3]. Mental rotation is performed in absence of real movement, and it can take place either in an egocentric or in an object-reference frame [
4]. In an egocentric reference-frame the visuo-spatial analysis refers to locations of objects from the observer’s point of view (i.e., frames locating things up-down, front-back, and left-right with respect to the observer; [
5]); in the object-reference frame analysis refers to environmental coordinates (i.e., frames locating things up-down, front-back, and left-right respect to external coordinates; [
6]. In object-based mental rotation tasks, the objects must be judged in relation to each other while the relationship between the environment and the observer remains fixed. In egocentric mental rotation tasks, typically involving body-related stimuli, individuals have to imagine themselves rotating in order to complete the task.
Mental rotation was first revealed in behavioral experiments [
7,
8] by measuring the time to make a judgment about a rotated object: the time often increases in a near-linear fashion with the amount of rotation required to bring the object into alignment with a comparison object or with a previously learned template [
9]. This effect has been observed with different kinds of stimuli: both geometric and “abstract”, such as letters, lines, polygons and three-dimensional cubes, and embodied and concrete, such as hands, legs, and whole-body figures [
10,
11,
12,
13].
Several other studies analyzed the relationship of mental rotation ability with development and age. They have shown that the ability to transform mental images is present in very young children, reaches higher levels during adolescence and declines with ageing (see data and literature in [
2]). The severity of the decline in mental rotation performance often depends on the stimuli used and the kind of task (object-based vs. egocentric; [
14]). It has been shown that mental rotation is facilitated with human figures as stimuli and egocentric tasks, and that males are generally more accurate than females [
14]. As regards gender differences in mental rotation, although several studies indicate robust effects generally favoring males [
15,
16], and agreed that these differences emerge at the age of around 8 years, the question is still matter of debate.
Previous studies, trying to allocate this abstract function in the brain, suggested that mental rotation of objects can belong to the right hemisphere, that of body images to the left, although with less evidence ([
9,
17,
18]. The issue is not solved yet, since many other studies claim against the right hemisphere dominance for objects’ mental rotation [
1,
19]. However, very seldom the mental rotation of the whole body image in a front-back direction was directly assessed [
20,
21,
22].
Mental rotation is strictly concerned with anatomical imitation, as suggested in previous behavioral studies. The imitation is the earliest form of relationship between individuals and can be considered the simplest form of social behavior. Imitation plays a central role in development, motor learning, communicative competence and acquisition of social skills. Generally, when asked to imitate a model gesturing facing the imitator, one can choose between two imitating strategies: a mirror-mode (specular), i.e., using the right limb to copy a spatially matched left limb gesture of the facing model, or an anatomical-mode, i.e., using the right limb for imitating an anatomically matched right limb gesture by the model. For successful anatomical imitation (3PA and 1PC; [
23]), individuals have to make a comparison between the representations of the self and the other; this operation involves a transformation of the egocentric perspective to the allocentric one [
24], which requires a mental rotation activity. Therefore, imitation requiring mental rotation can be defined as “anatomical”, while “mirror” refers to imitation which occurs without mental rotation [
23].
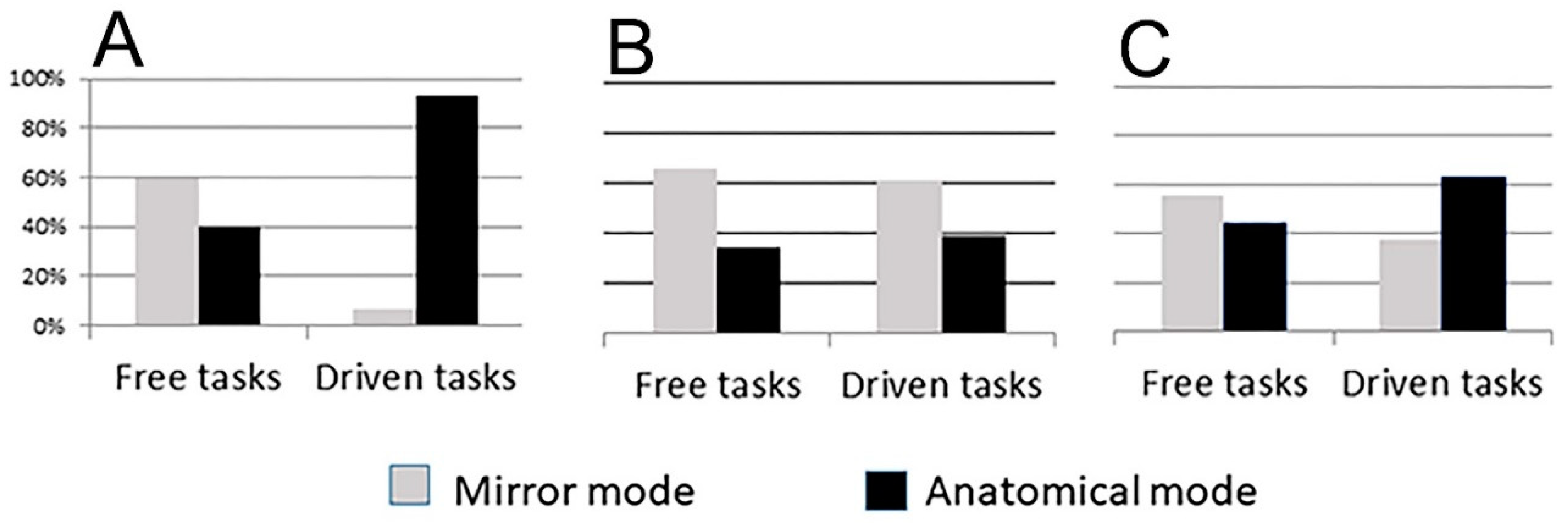
A recent research investigated the imitative behavior in patients with therapeutic corpus callosum (CC) resection and in healthy adults with an intact CC [
25,
26]. The study analyzed the perspective used by participants in imitating intransitive gestures. It has been shown that all subjects, when executing spontaneous imitation (free task), used the mirror mode (60% controls, 66% callosotomy patients;
Figure 1); when instructed to “use the same limb as the model” (driven task), the control group chose the anatomical mode (93%), but the callosotomy patients still preferred the mirror mode (i.e., 39% anatomical mode;
Figure 1; [
25,
26]). Later, an impairment of the anatomical imitative competence has been evidenced also in psychotic patients (41% anatomical imitation in free sessions, 64% in driven sessions;
Figure 1; [
27]), suggesting that individuals with surgical resection, microstructure alteration (as those reported in schizophrenic patients [
28,
29]), or incomplete maturation of the CC [
30], might be unable to execute the mental rotation. It has therefore been hypothesized that the different performances of the two patient groups with respect to the controls could possibly be due to an impaired capacity for mental rotation, in which the corpus callosum (CC) might have a role. The need of the CC seems thus to suggest that anatomical imitation, and therefore mental rotation ability on which anatomical imitation is based, are sustained by neural circuits including cortical areas of both hemispheres with different functions, according to an asymmetric distribution of competences.
Since a close correlation between anatomical imitation and mental rotation could be supposed, the present research investigated the ability to perform mental rotation in control subjects, in callosotomized patients and in psychotic patients. The purpose of this study was to test the hypotheses suggestive of a central role of the interhemispheric connections in mental rotation, since it is not clear yet whether this ability resides in the right or left hemisphere, or requires the cooperation of both. Two separate experimental sessions were provided; a verbal and a motor task. In the first, the verbal task, participants were asked to answer by voice whether the hand holding the cup in the displayed picture was the left or right hand. In this task it was possible to evaluate the mental rotation ability through a hand-laterality judgement. In the second, the motor task, participants were asked to indicate whether the hand holding the cup was the left or the right by lifting their own left or right hand. Preliminary results have been presented in abstract form [
31].
4. Discussion
The present research investigated the ability to perform mental rotation of body images in control subjects, callosotomized and psychotic patients. The choice to study the above mentioned two groups of patient was guided by the following reasons: 1. as previously reported, both callosotomized [
26] and psychotic patients [
35] showed an impaired ability, more severe in the first group, for anatomical imitation, which likely requires the abstract operation of mental rotation; 2. recent papers reported a partially impaired performance in mental rotation of letters and hand pictures in schizophrenic patients [
36,
37]. Since both kinds of patients are likely to display a reduction (more severe in the first group) of interhemispheric transfer, due either to interruption of callosal fibers (callosotomized patients), or to microstructural alterations of them (psychotic patients [
28,
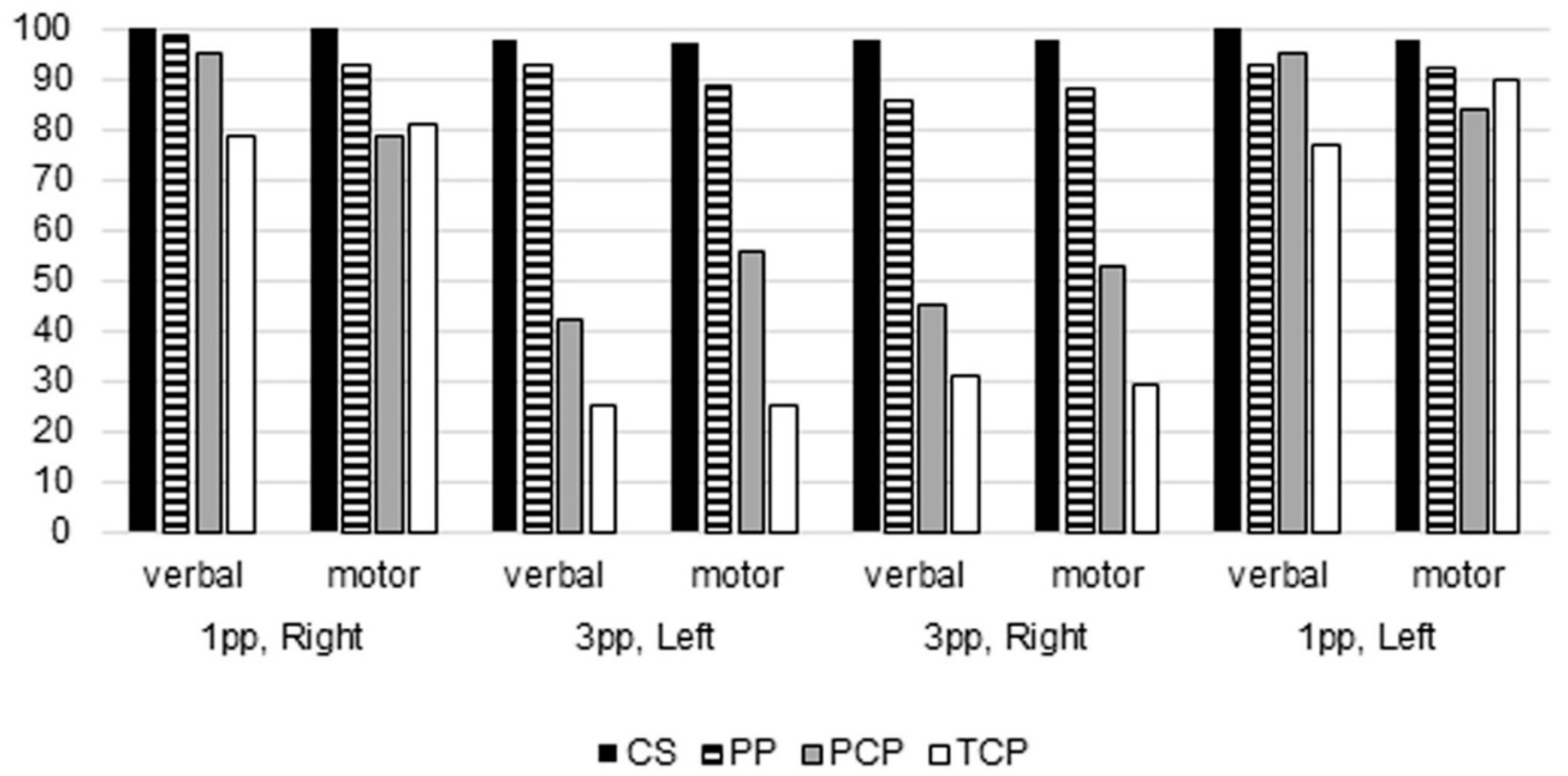
29]), the expected results were therefore to observe a reduced mental rotation ability of body images in both groups of patients, more pronounced in callosotomy ones. Actually, present data demonstrated that control subjects performed the task almost perfectly, in verbal and motor sessions, both when the model was in first-person perspective, and when she was in third-person perspective. Callosotomized patients showed some impairment, mainly in the verbal session and when the model was in third person perspective; psychotic patients too showed an impairment, although less pronounced than callosotomized ones. The present study indicated the central role of interhemispheric connections in mental rotation, and therefore, because of the need of the cooperation of both hemispheres to be performed, strongly suggest the value of considering mental rotation as an asymmetric function.
The statistical analysis demonstrated that the variable mostly influencing the subjects’ performance was the position of the model: when she was in third person perspective, both groups of patients displayed a higher proportion of incorrect responses, although psychotic patients performed slightly better than callosotomized patients.
The other variable influencing the performance was the presence of a callosal pathology: partial or total callosal resection, or microstructural callosal alteration likely present in psychotic patients. Actually, the performance obtained by psychotic patients was less poor than that of callosotomized patients, although it was still significantly lower than that of control subjects (
Table 3). A difference could be appreciated also between totally and partially callosotomized patients; however, the very low number of cases prevented an evaluation of the statistical significance.
No differences were observed in the present study depending on age, although the subjects were all adult, nor between women and men, in all groups. No differences were observed due to the position of the cup in the model’s hand (right or left), nor could any significant difference be appreciated due to the kind of session, whether verbal or motor.
The present results seem to indicate a role of the corpus callosum in the ability to perform the mental rotation, at least of the whole body image, in that the commissure is lacking in callosotomized patients, and likely impaired in psychotic patients. Further studies, verifying the functional integrity of the corpus callosum in psychotic patients could at least partially answer the question.
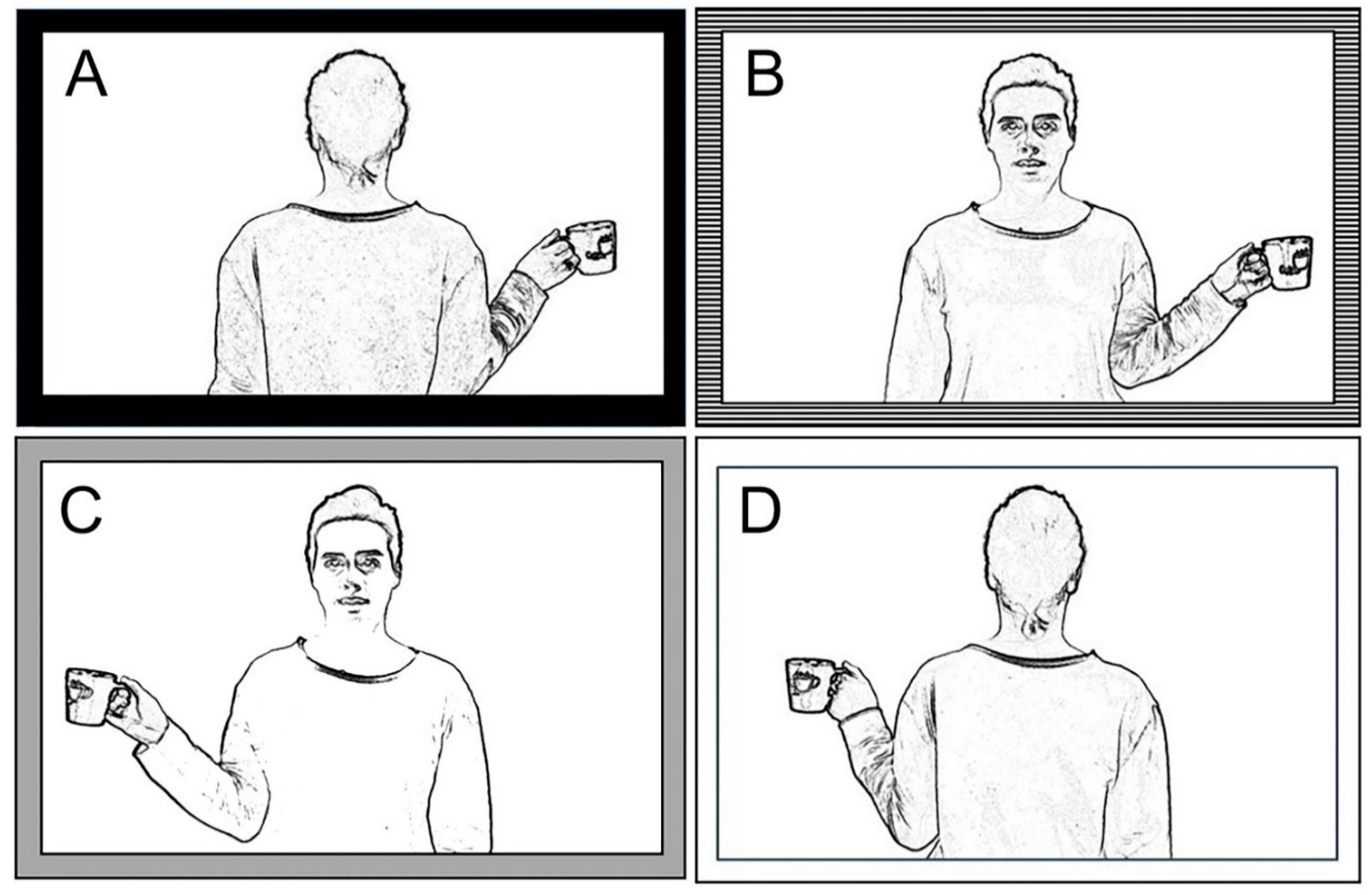
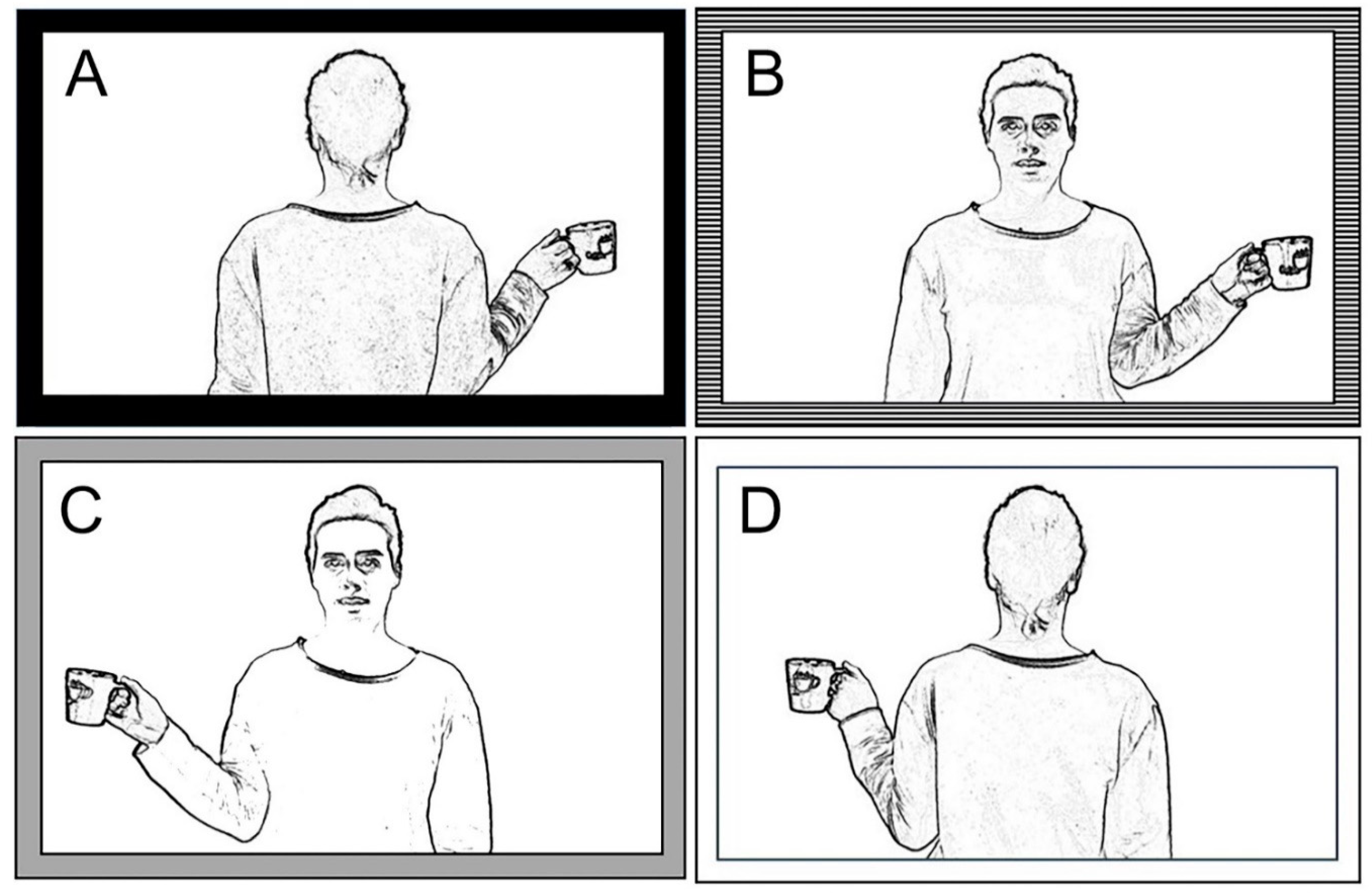
The present study analyzes an egocentric transformation task, requiring participants to indicate whether the model’s hand holding a cup is a right or a left hand; to answer, the participants have to look at a visual stimulus, a half body human picture presented in back or front perspective (0° or 180°). Behavioral results demonstrated that the orientation of the stimulus (front versus back position, i.e., third versus first person perspective) significantly influences the response correctness, mainly in callostomized patients.
The hypothesis advanced in the present work stays for the close correlation between stimulus orientation, mechanisms involved in processing the stimulus (i.e., visuo-spatial match versus mental rotation) and effect that an altered interhemispheric communication could exert onto the performance when the stimulus to be judged is placed in front of the observer, i.e., in third person perspective. As such, because of deficiency in interemispheric communication, callosotomized patients were unable to correctly respond to the task when the stimulus was in front-orientation, because of a failure of the neural circuit involved in mental rotation. It can be suggested, therefore, that mental rotation neural circuit is distributed in both hemispheres.
Many studies have been produced which analyze the egocentric transformation of the whole body. Some of them could be compared with ours, which is also based on an egocentric transformation of the whole body, in particular those describing experimental protocols similar to ours [
21,
22,
38,
39,
40,
41]. All of them reported as easier the task of mental rotation of body images, with the performance depending on the disparity angle, i.e., better responses for back (first person) presentation, worst for front (third person).
Parsons [
21] examined two related phenomena: the judgment of whether a human body part belongs to the left or right half of the body and the imagined spatial transformation of one’s body: observers made left-right judgments of a part of a body whose orientation differed from their own by a rotation about one of 13 axes. To do so, they imagined themselves passing to the orientation of the stimulus. Time for left-right judgments and accompanying imagined spatial transformations depended on the extent of the orientation difference between the observer and stimulus.
Thirioux and collegues [
38] investigated the behavioral and neural mechanisms of embodied and disembodied self-location in spontaneous interaction of healthy subjects with an avatar, where participants were instructed to observe the tightrope walker in front and back positions, and to lean when she was leaning (i.e., an imitation task-related). Embodied self-location was the condition in which the subject imagined their own body at the actual body position, and the reproduction of the avatar tilting occurred as in front of a mirror, adopting a visuo-spatial match. Disembodied self-location was the condition in which the imitator was imagining themself at the target body position, by performing a mental rotation of his/her body. The paradigm of that research [
38], in which the reproduction of a model’s movements occurred in two conditions, first (back position) and third (front position) perspective, could be considered similar to that used in the present study, and the results are comparable. Behavioral results reported correct performances in both tasks requiring embodied or disembodied self-location, and a significant tendency to use disembodied self-location in the spontaneous task. Interestingly, the higher percentage of correct performances was correlated with spontaneous and disembodied self-location performances in response to the stimulus in the back orientation, and reaction times increased when the rotation angle between the participant and the avatar increased.
In the study of Crescentini and coworkers [
22], an “own body transformation” paradigm and a letter transformation task were employed in a group of children ranging from 7 to 11 years of age to investigate the development of egocentric perspective transformations and object-related transformations. A control group of young adults was also administered the two experimental tasks. The results from this latter group for egocentric perspective transformations was similar to those of the present study, by showing that the reaction time was lower and the accuracy was higher for back (first person) presentation. In addition, they found that egocentric perspective transformations develop later than object-based transformations, namely, from 8 rather than 7 years of age. These findings were held to support the view that the two mental spatial transformation systems are separated in that they follow two different developmental trajectories.
Other studies [
40] investigated developmental changes in mental rotation performance with a focus on two types of strategies: object-based and egocentric transformations. This study revealed two important findings: 1. an age-related decline in processing speed and the possible role of white matter functionality in mental rotation performance; 2. the observation that children and older adults seem to show deficits in perspective-taking compared to adults. Therefore, they propose that perspective transformations are more sensitive to developmental change compared to object-based transformations.
Jansen and coworkers [
41] investigated gender differences in performance and neuronal activity in a mental rotation task with abstract and embodied figures. Participants (26 females and 32 males) completed a chronometric mental rotation task with cube figures, human figures, and body postures. The results were straightforward: depending on angular disparity, participants had a faster reaction time and a higher accuracy rate for embodied stimuli compared to cube figures. They also recorded electroencephalogram (EEG) activity pattern, and showed alterations at the late time interval, even though there were no gender differences in the behavioral data. Thus, the disparate results regarding sex differences that depend on the type of analysis (behavioral versus neurophysiological) should be more thoroughly investigated.
The behavioral data just described all agree in the observation that, concerning egocentric transformation, back orientation produces better performances than front orientation. The difference increases in childhood and senior age (both periods of life in which alterations of the cerebral white matter could be observed), in children because of a still incomplete development, and in older people because of an initial white matter decline. The results of the present study, which reports correct performances in first person perspective (“back orientation”) for all participants (controls, callosotomized and psychotics), and reduced performances in psychotic and callosotomized patients, in whom the callosal white matter is damaged, are in line with previous studies.
None of the behavioral studies above mentioned, however, addressed the issue of asymmetric cerebral competence for mental rotation. This notion, strongly suggested by the results of the present research, came out in functional studies, reviewed by Zacks [
9], and later by Tomasino and Gremese [
40], through an activation likelihood estimation (ALE) meta-analysis. Some observations, suggesting a cerebral specialization for mental rotation, also appeared in previous studies ([
18,
20,
38].
In his review and meta-analysis of neuroimaging studies [
9] Zacks posed the research question addressing to whether mental rotation relies on analog spatial representations or on motor simulation. The results of the meta-analysis conducted on 32 neuroimaging studies highlighted a total of 320 activation foci responsive to mental rotation tasks, and included the superior parietal, frontal, and inferotemporal cortex. Most of these brain regions were activated bilaterally, except for the parietal cortex, whose activity was somewhat more consistently observed in the right hemisphere, and the frontal cortex, whose activity was more consistently observed in the left hemisphere [
9].
Tomasino and Gremese [
42] selected 171 fMRI and PET experiments that investigated mental rotation processes, suggesting a different modulation of mental rotation networks respect the type of stimulus and strategy use to perform the task (motor imagery based versus visuo-spatial imagery). Studies included in motor imagery mental rotation tasks conteined explicit instructions requiring participants to imagine one’s movements or change the body position in space; in studies included in visuo-spatial mental rotation task, participants were explicitly required to imagine the stimulus rotating in the space (e.g., in looking at stimuli pair, choosing if they are identical or mirror or imagining that the table rotated while participant remained stationary; allocentric frame). A further distinction was made within the motor-imagery based mental rotation strategies, between pure motor-imagery (e.g., imagining to rotate one’s own arm until this reached the position depicted in the screen) and egocentric mental rotation (e.g., updated the position of an external object after having performed an imagined self-rotation to a new position). In both conditions, a mental rotation of stimuli was requested.
Comparison between motor-imagery-based and egocentric-based mental rotation revealed activations of the left superior parietal lobule, the right postcentral gyrus, the precentral gyrus/middle and superior frontal gyrus bilaterally, and the left inferior occipital gyrus; the inverse analysis activated the left cuneus, the left middle temporal gyrus, the left lingual gyrus and calcarine sulcus, and the right cerebellum.
In the above mentioned study [
38], event-related potentials (EPs) were calculated from continuous EEG acquired during the task. EPs mapping revealed different stable brain microstates underlying the spontaneous and disembodied-self-location tasks and the embodied self-location task, suggesting the recruitment of different neural generators. Specifically, in the first two conditions, the involvement of the temporo-parietal junction bilaterally, with a right predominant activation, was observed; also, the posterior parietal cortex and the intraparietal sulcus were activated bilaterally, more prominently in the left hemisphere. During the embodied self-location task, more activated regions were observed in the right hemisphere: the inferior occipital cortex, the postcentral gyrus, the premotor cortex, the dorsolateral prefrontal cortex, the anterior temporal cortex and weaker activation in the temporo-parietal junction. In addition, EP data also showed that brain activations during spontaneous and explicit disembodied self-transformations started earlier for the back than for the front orientation. This could suggest higher difficulty for body transformation in front of a front-oriented stimulus. Summarizing, these data seem to support a prevalent leftward asymmetry in the cortical activation for disembodied self-location, corresponding to the mental rotation for a third person model’s presentation of our research, and a rightward asymmetry for the embodied self-location task (i.e., mirror mode, visuo-spatial match).
Participants in the present behavioral study had to produce two different types of response to the same task; in the verbal modality, they observed the target stimulus and reported by voice to the experimenter in which hand (left or right) the model held the cup; in the motor modality, the subjects must raise the hand corresponding to that of the model which held the cup. With respect to type of stimuli, both tasks are laterality judgement like, since target stimuli were body images, presented in first or third perspective, holding a cup in either right or left hand. Therefore, both tasks are supposed to recruit mental rotation when responding to front-facing stimuli, even if through different procedural response-modes (verbal plus motor).
By analyzing previous functional studies [
9,
38,
42], despite the diversity of the investigation approaches (fMRI, EP), and the variables investigated (e.g., instructions, type of stimuli, theoretical definitions), it is possible to identify a common trend of scientific evidences correlating mental rotation to a sort of hemispheric specialization. Specifically, a left lateralization for the parietal cortex and a right specialization for the frontal regions. In an attempt to find likeness between these studies [
38,
42], a certain factor of concordance can be observed in identifying lateralized activations in motor, sensorimotor and parietal regions during mental rotation. Specifically, the parietal lobe seems to be leftward lateralized, while precentral and postcentral regions appear to be rightward lateralized.
The present paper demonstrates that people with partial or total callosal resection, or with callosal alterations like those reported in schizophrenia, display reduced performance in laterality testing with stimuli in third-person orientation, suggesting an alteration of mental rotation mechanisms, likely due to a defective interhemispheric communication. These results are in line with previous studies reporting different activations of cortical areas in the two hemispheres, indicating the need for an interhemispheric cooperation, and therefore for an interhemispheric transfer. Present results are also on line with previous indirect evidence suggesting callosal involvement, such as the reduced mental rotation ability of children and old people, likely due to an incomplete callosal maturation and to an initial callosal degeneration, respectively. In conclusion, the present study provides further support to the notion that the mental rotation of body images does require interhemispheric communication, thus leading to the notion of an asymmetric competence.

 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}