Abstract
Language-induced asymmetry to single word reading has been well investigated in past research. Less known are the complex processes and related asymmetries occurring when a word is compared with the previous one, according to specific tasks. To this end, we used a paradigm based on 80 sequential word pair comparisons and three blocked tasks: phonological, semantic and orthographical matching judgment. Participants had to decide whether the target word (W2) did or did not match the prime word (W1), presented 2 sec before, according to the task. The event-related potentials (ERPs) evoked by W2 in 20 participants have been analyzed. The first negative wave, the Recognition Potential (RP), peaking at about 120 ms over parietal sites, showed greater amplitude at left sites in all tasks, thus revealing the typical left-lateralization. At frontal sites, only the phonological task showed left lateralization. The following N400 (300–450 ms) showed an interesting interaction: Match trials elicited greater left asymmetry on frontal regions to phonological than to semantic than to visual-perceptual tasks, whereas mismatch trials induced an inverted asymmetry, marked by greater amplitude over right frontal sites, regardless of the task. Concerning the late N400 (450–600 ms), phonological and semantic tasks showed an overlapping pattern, with left lateralization in match and right lateralization in mismatch conditions. Results point to complex task- and time-dependent hemispheric asymmetries in word matching.
1. Introduction
Forty years have passed since Kutas and Hillyard [1] first proposed a new paradigm aimed at applying the oddball task to the study of language: In a small group of sentences, they changed the final correct word with an unexpected, semantically wrong word with the aim of inducing a P300 evoked response. Unexpectedly, they found a significantly greater negativity, peaking at about 400 ms after the unrelated final word, and termed this new component N400 [1]. This study has represented the benchmark for the following investigations within the linguistic domain, and the N400 has been largely studied using various experimental paradigms see, for examples, reviews [2,3,4]. The N400 paradigm has been successfully tested in the Italian language, with violations placed in the middle rather than at the end of the sentence [5], and with an interesting drag effect extending N400 to all words following the target word. In general, results showed considerable consistency, particularly in the visual modality, revealing greater N400 amplitude on right rather than left electrodes of mismatch stimuli, e.g., [2,5,6,7,8], especially at the level of fronto-parietal cortical sites. Among others, in addition to sentence reading, single words or the prime-target word-pair paradigm have been used to induce the N400, in particular during mismatch conditions [9,10,11]. Interestingly, in their review, Van Petten and Luka [2] highlighted that, when participants carry out a rhyming judgment task on visually presented words, mismatch stimuli elicit a right-lateralized negative potential similar to the semantic N400 effect: thus, greater right N400 amplitude may represent a marker of mismatching condition, regardless of task-dependent linguistic process. In 1995, Van Petten and Rheinfelder [12] referred to this phenomenon in terms of “paradoxical lateralization,” suggesting that small rightward asymmetry represents a warped cortical projection of electrical dipole activity from left hemisphere N400 generators [2,3]. Indeed, when source estimations were carried out, the cortical region associated with the N400 component showed inconsistent results, e.g., [13,14,15]; nevertheless, studies on neurological and split-brain patients located the generator in the left-hemisphere [2,16].
Notwithstanding the great amount of literature focused on the prime-target word-pair paradigm, e.g., [3,4], and the match/mismatch N400 component in particular, less attention has been paid to the study of the complex temporal dynamics following target word onset, i.e., when the participant must decide whether the second word (W2) does or does not match the previous one (prime word, W1). Indeed, before the linguistic classification takes place in our brain, thus allowing us to distinguish whether the word pair accomplishes the task requirement, we automatically (and probably without being aware) recognize that this target stimulus (W2) is a word, rather than other visual stimuli (such as objects, pictures, faces and so on). This automatic word recognition is marked by a well-known ERP component, known as word recognition potential (RP), which appears as a negative wave at parieto-occipital sites that reaches maximum amplitude about 120–130 ms after word onset, e.g., [17,18,19,20]. At the same time, less attention has been devoted to the study of N400 lateralization and scalp distribution to match trials, and their possible association with participants’ behavioral performance.
Therefore, the present study aimed to investigate the temporal dynamics following W2 onset in a well-validated prime-target paradigm [17,21], in which word pairs were administered, phonological and semantic processes were required (in two separate tasks, i.e., the rhyming and the semantic judgment tasks) and a further orthographical matching judgment task (on word pairs) was included. As we validated our experimental paradigm on prime word (W1) stimulus analysis in a middle age group, we decided to consider this same age group for the analysis of event-related components associated with W2 processing. Starting from results found in past studies focusing on W1 stimuli, we expected W2 to elicit the typical word Recognition Potential (RP), peaking about 120–170 ms, e.g., [17,18,22] at left posterior sites. As there is no evidence suggesting early match/mismatch differences before N400 components, we expected no effects of word matching on RP. At the same time, we expected the N400 effect elicited by both match and mismatch word pairs, at fronto-centro-parietal sites, about 350–500 ms after W2 onset. With respect to cortical asymmetries, we hypothesized greater N400 amplitude at right vs. left sites (i.e., a significant rightward effect) to mismatch stimuli, regardless of linguistic processing (i.e., phonological or semantic judgment), in agreement with past literature on visual N400 effect [2,3,7,8]. Less data are available on the N400 in word-pair matching: In a study from our group [17], a left frontal N400 to W1 in word pairs was found. It was named N350 to distinguish it from the past paradigms investigated on N400. We expected that, in the match condition, W2 would elicit greater N400 amplitude at left vs. right sites (i.e., a clear left-sided N400).
2. Results
To focus on the main results, only the most salient interactions are discussed in the paper. All significant results are presented in the Supplementary Materials.
Analysis of response times (RTs) revealed a significant two-way task × condition interaction (F2,38 = 6.04, p < 0.01, HF ε = 0.83, η2 = 0.24), with faster RTs on the orthographical matching judgment task than on the phonological (all ps < 0.05) and semantic tasks (all ps < 0.001), in both match and mismatch conditions (Figure 1).
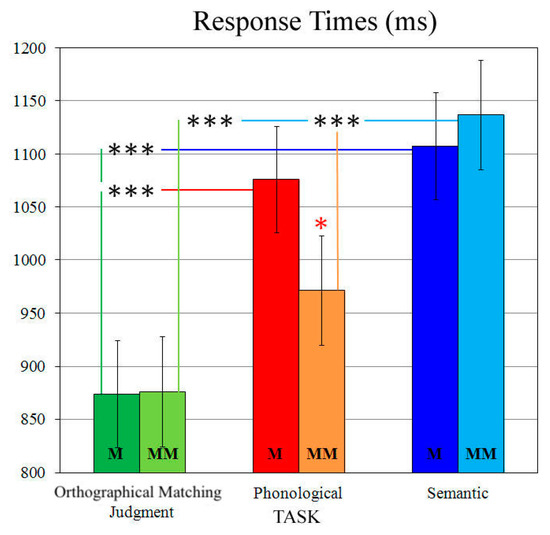
Figure 1.
Response time (RT) analysis: Significant two-way Task × Condition interaction. Asterisks: Significant post hoc comparisons (Tukey HSD, *** p < 0.001, * p < 0.05).
Furthermore, mismatch stimuli resulted in slower RTs on the sematic vs. phonological task (p < 0.001), whereas no differences between linguistic tasks were found in match conditions (Figure 1). Error rates showed neither significant main effects nor interactions (all ps > 0.15). For this reason, only RTs were considered for correlation analyses.
As can be seen in Figure 2A, the typical word RP occurred at parieto-occipital sites about 110–130 ms after W2 onset, showing more pronounced negativity at left sites for all tasks and word conditions.
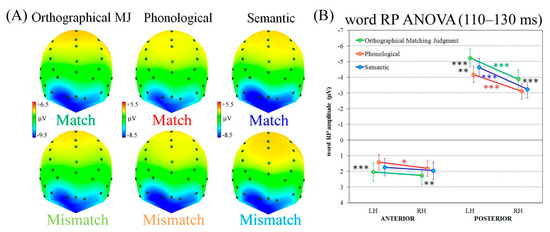
Figure 2.
Early component analysis. (A) ERP spline-interpolated maps representing the four regions of interest (ROIs) and the scalp back/top view of RP component following W2 onset for Match (top line) and Mismatch trials (bottom line). (B) Word RP analysis showing the significant three-way Task × Region × Laterality interaction. LH = left hemisphere; RH = right hemisphere. Asterisks: Significant post hoc comparisons (Tukey HSD, *** p < 0.001, ** p < 0.01, * p < 0.05).
Indeed, the significant three-way Task × Region × Laterality interaction (F2,38 = 3.30, p < 0.05, HF ε = 0.80, η2 = 0.15) revealed that all tasks exhibited a marked greater left vs. right negativity at posterior sites (all ps < 0.001). However, the left-lateralized word RP exhibited the greatest amplitude in posterior clusters during the orthographical matching judgment task than the other two linguistic tasks (all ps < 0.001). In turn, the semantic categorization task showed greater negativity at posterior left sites than the phonological one (p < 0.01). Concerning anterior regions, the phonological task only revealed (relatively) greater left vs. right lateralization (p < 0.05), whereas the other two tasks exhibited bilateral positivity (Figure 2B). The phonological task showed relatively greater negativity than the orthographical matching judgment task at both left and right sites (p < 0.001 and p < 0.01, respectively). No significant correlations were found between RP lateralization and RTs (details in Supplementary Materials).
The second time window analyzed (300–450 ms interval following W2 onset) showed a clear negative wave, the N400, with opposite laterality patterns of activation depending on match or mismatch condition. Indeed, as can be seen in the top row of Figure 3A, in the match condition a clear negativity appeared on left sites during phonological and semantic processing, but not during the orthographical matching judgment task. When words mismatched, instead, the negative wave developed at the right anterior-central electrodes, particularly during both linguistic tasks (bottom row of Figure 3A).
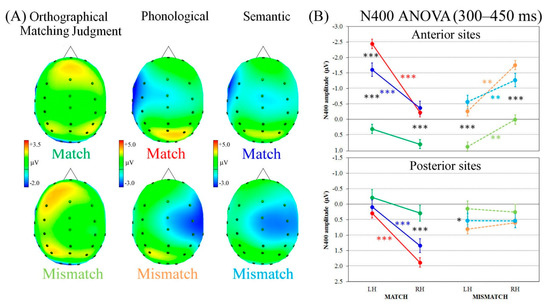
Figure 3.
N400 component analysis. (A) ERP spline-interpolated maps representing the four ROIs and the scalp top view of N400 component following W2 onset for Match (top line) and Mismatch trials (bottom line). (B) N400 analysis evidencing the significant four-way Task × Condition × Region × Laterality interaction. LH = left hemisphere; RH = right hemisphere. Asterisks: Significant post hoc comparisons (Tukey HSD, *** p < 0.001 ** p < 0.01 * p < 0.05).
Statistical analysis revealed a significant four-way Task × Condition × Region × Laterality interaction (F2,38 = 4.01, p < 0.05, HF ε = 1.00, η2 = 0.17). In terms of match conditions, both linguistic tasks exhibited significantly greater negativity over the left vs. right hemisphere, regardless of region (all ps < 0.001), whereas the orthographical matching judgment task showed a bilateral activation (Figure 3B, left panel). Furthermore, at anterior left sites, the phonological task showed greater N400 negativity than the semantic one (p < 0.001), which in turn elicited greater negativity than the orthographical matching judgment task (p < 0.001). At anterior right sites, instead, both linguistic tasks showed greater negativity than the control one (all ps < 0.001). At posterior right regions, the orthographical matching judgment task showed significantly (relatively) greater negativity than linguistic tasks (all ps < 0.001). In terms of mismatch conditions, all tasks revealed significantly greater N400 negativity at right vs. left anterior sites (all ps < 0.01), and a bilateral pattern of activation in posterior regions (Figure 3B, right panel). Furthermore, at anterior sites, both linguistic tasks showed greater negativity than the orthographical matching judgment task (all ps < 0.001). In posterior left regions, the latter task showed significantly (relatively) greater negativity than the phonological task only (p < 0.05), whereas no differences were found at right sites.
Pearson’s correlation analysis revealed significant positive associations between anterior laterality indices and RTs for orthographical matching judgment and phonological tasks, for the match condition only (r18 = 0.52, p = 0.018 and r18 = 0.47, p = 0.035, respectively): The greater the N400 negativity on anterior left sites, the faster the later RTs for orthographical matching judgment and phonological tasks during match conditions. No correlations appeared associated with semantic processing, nor with mismatch trials (details in Supplementary Materials).
The last time window analyzed (450–600 ms interval following W2 onset) showed a clear late negative wave, with scalp distribution and effects similar to those showed by the prior time window, therefore termed late N400, again with opposite laterality patterns of activation, depending on match or mismatch condition. Indeed, as can be seen in the top row of Figure 4A, in the match condition, a clear negativity appeared at left sites during phonological and semantic processing, as well as during the orthographical matching judgment task, although with lower negativity levels. By contrast, when words mismatched, the late N400 developed at right anterior-central electrodes, particularly during both linguistic tasks (bottom row of Figure 4A).
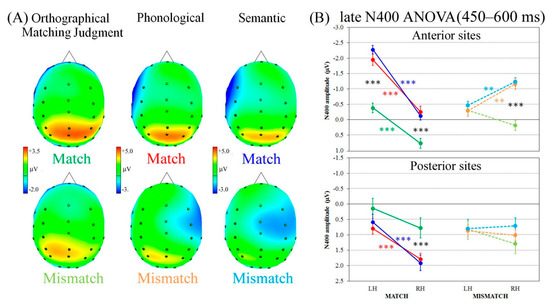
Figure 4.
Late N400 component analysis. (A) ERP spline-interpolated maps representing the four ROIs and the scalp top view of Late Negative Potential (LNP) following W2 onset for Match (top line) and Mismatch trials (bottom line). (B) Late N400 analysis revealing the significant four-way Task × Condition × Region × Laterality interaction. LH = left hemisphere; RH = right hemisphere. Asterisks: Significant post hoc comparisons (Tukey HSD, *** p < 0.001, ** p < 0.01).
Statistical analysis revealed a significant four-way Task × Condition × Region × Laterality interaction (F2,38 = 3.73, p < 0.05, HF ε = 1.00, η2 = 0.16). The linguistic tasks showed the same N400 pattern, marked by greater left vs. right lateralization during match conditions (all ps < 0.001) and right vs. left lateralization during mismatch conditions (all ps < 0.01), with no differences on negativity amplitude (Figure 4B), whereas the orthographical matching judgment task showed a left-lateralized late negative wave at anterior sites (p < 0.001) during the match condition, and a bilateral pattern of activation in posterior regions, as well as during the mismatch condition (regardless of region).
As for the previous ERP component, Pearson’s correlation analysis revealed positive links between anterior laterality indices and RTs for the match condition only, but in this later interval, all tasks showed significant associations: The greater the negativity of the late negative wave at anterior left sites, the faster the RTs for orthographical matching judgment (r18 = 0.57, p = 0.009), phonological, (r18 = 0.56, p = 0.010) and semantic (r18 = 0.50, p = 0.023) tasks during the match condition. No correlations were found for the mismatch condition (details on Supplementary Materials).
3. Discussion
The present study was carried out to clarify the temporal dynamics of processes occurring in a prime-target word-pair task, after the presentation of W2. To this end, we used a well-validated prime-target paradigm, in which word pairs were administered in two linguistic tasks, phonological and semantic categorization, compared with an orthographical matching judgment task serving as a control [21]. Although the word-pair paradigm is substantially different from the classical paradigm based on sentences and evoking a N400 [2,3,4], we expected a N400 also in the Italian language [5,6] and in serial word-pair matching [17]. In addition to the classical paradigm based on sentences, several studies have been carried out on N400 elicited by single words or word pairs [9,10,11]. Furthermore, as past research revealed late dragging effects of mismatch-elicited N400 [5], we were also interested in investigating possible late, closing processes associated with an N400 sustained across time. Indeed, our results clearly showed a sustained N400 that was divided into two intervals, one in line with past literature (300–450 ms) and the second, termed late N400, measured in the 450–600 ms time interval. The scalp distribution of the two components was relatively similar (maps in Figure 3 and Figure 4), although statistical effects differed slightly.
Behavioral results showed an interesting correlation on response times: In the match condition the speeds for both linguistic tasks (phonological and semantic) were slower than for the control task, but the two judgments required the same time. The mismatch condition revealed a clear difference among all tasks, as response times on the semantic task were slower than on the phonological task than on the visuo-spatial task. Thus, in match conditions, the two linguistic tasks required the same resources during the word-pair comparison, whereas in the mismatch condition, the semantic task required more resources. Deciding that two words do not belong to the same category is probably more demanding, due to its more complex serial processing aimed at analyzing many semantic nodes and excluding many similar confounding categories. Instead, the process leading to a choice of whether a word pair rhymes or not is symmetric, because deciding that a word pair does not rhyme is very quick and easy, to the same extent that deciding that a word pair rhymes.
Similar to past results, in which we focused mainly on W1 stimuli, e.g., [17,18,21], in the present study, we found the typical word Recognition Potential (RP) at posterior sites after W2. This wave reached maximum amplitude about 120 ms after target word onset, with significant leftward lateralization on all tasks. As expected, no differences in match/mismatch word condition were found, because the complex decisional processes in the linguistic domain needs far more time: indeed, the average response time was around 1000 ms, whereas the RP is a very early perceptual process. However, we found significant RP amplitude differences among tasks: the orthographical matching judgment task showed the greatest amplitude, whereas the semantic categorization exhibited greater amplitude with respect to the phonological task. At the same time, the rhyming judgment was the only task that induced leftward asymmetry at anterior sites, whereas the other two tasks exhibited bilateral positivity. These results therefore suggest that phonological processing involves a more distributed linguistic network within the left hemisphere, including both posterior and anterior language centers, like the first, automatic phases of word processing, whereas the semantic categorization task mainly involves only the posterior networks, at least in this early linguistic encoding. Finally, the orthographical matching judgment task, which forces processing of visual properties of words, showed the maximum RP amplitude, probably because no other complex analyses were required, apart from automatic word recognition, and therefore only local occipital hubs directly involved in the required process were active with respect to all other regions. Instead, linguistic tasks recruit several other regions (such as frontal ones in the phonological task) and this tends to lower the relative amplitude of the RP peak with respect to the surrounding areas. This early difference among tasks probably occurred because we administered one task at a time, thus allowing participants to ignite, within the whole block, the appropriate language network involved in each task and to perform efficiently. We found no evidence of match/mismatch differences in this early phase of linguistic processing. The leftward RP amplitude modulation, together with the different left-lateralized distribution along the antero-posterior gradient, suggests that, about 120 ms after W2 onset, the main process occurring is perceptual recognition of the stimulus as a linguistic one, whereas the task requirement, based on the complex process of W1-W2 comparison followed by a decision, is very demanding and requires, as already mentioned, about 900–1100 ms. In line with this, correlation analysis showed no association between this early component and response times.
Concerning the second component, a clear N400 effect was elicited by both matched and mismatched word pairs, and was mainly evident at fronto-central sites, about 300–450 ms after W2 onset. With respect to cortical asymmetries, a rightward N400 effect in the mismatch condition, regardless of linguistic processing (i.e., phonological or semantic judgment), was found, in agreement with past literature on the visual N400 effect [2,3,5,6,7]. In addition, the mismatch condition during the orthographical matching judgment task showed significant right lateralization, but, compared with the other two tasks, this was associated with significantly lower N400 amplitude at both left and right sites of anterior ROI. No laterality effects were found at posterior sites, regardless of the task, during the mismatch condition. A reversed lateralization was found in the match condition, which evidenced a clear left N400, marking linguistic tasks at both frontal and parietal sites. Instead, the orthographical matching judgment task showed bilateral activation. In addition, at anterior left sites, the match condition on the phonological task elicited a greater N400 amplitude than words on the semantic one, thus confirming the greater involvement of left frontal sites during phonological processing. At this stage, the phonological task was shown to be the most lateralized at left frontal sites during match trials: We hypothesize that the rhyming process required by the task boosts the underlying activation of rehearsal of the W1 in the verbal working memory in the process termed “articulatory loop,” comparing the phonological features of W1 with those of W2. This process strengthens the activity and the connectivity between anterior and posterior left regions, thus leading to enhanced left N400 lateralization at both anterior and posterior sites.
In agreement with this interpretation, the correlation analysis revealed a significant association between left frontal N400 and RTs obtained on phonological word matching: The greater the N400 amplitude at left anterior sites, the faster the later RTs on the phonological task during matching conditions, whereas no correlations appeared associated with semantic processing, nor with mismatch trials at all. Therefore, the described correlation is a functional one, as the greater the left N400, the faster were the responses to the W2. We interpret the lack of correlation of left N400 elicited by semantic task as being due to the complex process involved in the search for a threshold among the many lexical-semantic nodes elicited by both W1 and W2: Only when the proximity between the word pair decreases to a specific threshold is it possible to decide whether the two words match or not. This process is intrinsically noisy, whereas deciding whether a word pair rhymes or not is much more straightforward.
Concerning the latest component in the 450–600 ms interval, this was clearly a late N400, in continuity with the previous N400 effect, particularly at fronto-central sites. As for the previous ERP component, only the linguistic tasks (phonological and semantic) elicited rightward lateralization in anterior regions in mismatch trials, and leftward lateralization in match trials. Interestingly, whereas in the prior interval, the orthographical matching judgment task induced a right lateralization during mismatch, in this late interval, it elicited a leftward lateralization in the match condition, so it seems that, for this task, at the cortical level, the mismatch condition was detected earlier than the match one. Pearson’s correlation analyses revealed positive links between anterior laterality indices and RTs on all tasks, for the match condition only, whereas no correlations were found for the mismatch condition. Those participants exhibiting greater left hemisphere activation during the match condition had faster RTs on the corresponding behavioral trials and this was similar on all tasks. Indeed, at this stage, the decision process is complete and left frontal activation reflects the speed of the response, regardless of the task: this may reveal a general mechanism related to the matching process. Interestingly, the motor decision process is located at the right premotor frontal sites, as all participants responded with the left hand; this was planned in the original experimental design to avoid activation of left hemisphere sites for motor responses, which would have made it difficult to interpret results. As motor planning and response are processed in the right premotor and motor areas respectively, the interpretation of left N400 is not confounded by this process, which suggests that the underlying late process we found is related more to language than to motor response. We hypothesize that this mechanism is represented by verbal working memory and the underlying networks operating in the left perisylvian areas are both involved, to some extent, in all tasks [23], thus indicating a general language mechanism common to all languages [24]. In line with this, reviews of neuroimaging studies have pinpointed how the left frontal regions play an important dominant hierarchical role in all linguistic processes, including the phonological, semantic, and syntactic ones, with a specific segregation of each within Broca’s area [25,26]. Of course, with the ERP method used here, we cannot provide regional details comparable with fMRI and PET methods.
In conclusion, analysis of the potentials evoked by W2 in a word-pair linguistic-matching paradigm showed interesting and complex temporal dynamics of the three tasks. In the first phase, there was prevalence of a perceptual process focused on the recognition of the stimulus as a linguistic one; this stage was characterized by a clear typical left posterior negative peak (RP) at about 120 ms. Nevertheless, within this general pattern, small differences among the blocked tasks emerged, with the phonological task left lateralized at frontal sites. In the second stage, the effect of tasks diverged, a clear lateralized N400 was found in frontal clusters, peaking in the right hemisphere during mismatch, and in the left hemisphere during match trials. The phonological task elicited the strongest frontal left N400. In the last stage after the N400, corresponding to the late N400, a very similar pattern was found in continuity with prior phase, but the two linguistic tasks showed an overlapping pattern: At this level, differences between semantic and phonological tasks disappeared, whereas the difference between these and the less lateralized visuo-spatial task remained. The time course and cortical distribution of the N400 and the subsequent late N400 suggested that our component belongs to the family of the N400s, but it is important to highlight that in our word-pair matching paradigm, the N400 was much more lateralized and less central than that investigated in sentence reading. In this paradigm, linguistic matching shifted the N400 more to the left hemisphere and mismatch more to the right one. This may be related to a strong hemispheric reciprocal inhibition, with clear left hemispheric activation forced by the verbal working memory demands to recall W1 (and its task-related features) for comparison with the next W2. When W1-W2 linguistic matching occurred, the left lateralization was strengthened, but when expectancy was denied by a mismatch trial, there was relative inhibition of the left hemisphere and activation of the N400 in the right hemisphere.
The three-stage effects we found shed light on the underlying complex processes occurring during word-pair matching. As the integration of many processes is necessary in more complex and ecological frameworks of linguistic human experience, the analysis of W2 in similar paradigms may represent a promising marker for studying cognitive deficits of psychiatric (e.g., depressed or schizophrenic) and neurological patients. The present study tested the N400 only in the Italian language, so it would be interesting to know whether the same effects can be found in other languages. According to current literature, the N400 is a component quite consistent and similar across many languages, including English, Italian, Dutch, and German [6]. There is evidence that, in bilinguals, the N400 is similar in both languages, with bilinguals having slightly slower latencies and the less proficient language eliciting relatively longer latencies [27]. An N400 elicited by word priming and more pronounced in the right hemisphere was found in 24-month-old children speaking French [28]; thus, this component is relatively automatic and similar across ages, bilingualism, and languages. Finally, our word-pair matching paradigm was tested among students speaking Italian and German languages, and we found overlapping scalp distributions [23]. Therefore, the replication of this paradigm in other languages and conditions would be interesting, and it may be expected to lead to results similar to the present ones.
In the present study, we considered a middle-aged adult group, to validate the experimental paradigm and to allow W2 analysis. A possible limitation may therefore consist of the potential influence of age. However, in a past study carried out on three age groups (children, young adults, and middle-aged participants) and measuring RP and N350 components evoked by W1 [18], we found no significant differences between the two adult samples (the older one having the same mean age as the sample in the present study). From this perspective, a future study may focus on potentials evoked by W2 stimuli in different age groups (following our past research [18]), namely RP, N400, and late N400, to directly test possible age-dependent effects on hemispheric asymmetry.
4. Materials and Methods
4.1. Participants
Twenty native Italian adults (7 females, mean age 59.10, SD ± 7.11 years, range 43–69 years) gave their written consent to take part in the experiment. All participants were fully right-handed, on average 97.35% (SD ± 6.91%, range 75–100%), according to the Edinburgh Handedness Inventory [29]. As past literature showed that bilinguals, even those who are late bilingual or good at a second language, may have a different organization of language in the brain, especially in terms of functional laterality [30,31,32,33,34,35], it is important to highlight that participants of the present study were all monolingual, Italian speakers.
None of the participants was treated for any neurological or psychiatric disorder, nor was on any current medication regime. Experimental procedures were approved by the local Ethics Committee.
4.2. Apparatus and Physiological Recordings
EEG data were continuously recorded in DC mode in the psychophysiology laboratory at the Department of General Psychology (Padova), with a low-pass filter set to 100 Hz, sampling rate of 500 Hz, and amplitude resolution of 0.168 μV/bin. EEG was measured by means of 38 tin electrodes, using SynAmps amplifiers (NeuroScan Labs, Sterling, VA, USA), 31 mounted (i.e., Fp1, FPz, Fp2, F3, Fz, F4, FC3, FCz, FC4, C3, Cz, C4, CP3, CPz, CP4, P3, Pz, P4, F7, F8, FT7, FT8, T7, T8, TP7, TP8,P7, P8, O1, Oz, O2) on an elastic cap (ElectroCap) according to the International 10–20 system [36]; the other seven electrodes were applied below each eye (Io1, Io2), on the two external canthii (F9, F10), on the Nasion (Nz), and on the mastoids (M1, M2). Cz was used as an on-line recording reference for the EEG channels, and then data were converted off-line to the average reference.
4.3. Stimuli, Tasks, and Procedure
Stimuli consisted of bi- or tri-syllabic Italian content words (mean: 2.40 syllables, SD ± 0.49 syllables) selected from a frequency dictionary of 5000 written Italian words [37]. Words were presented in pairs, one at time, on a 17” computer monitor, with an inter-stimulus interval of 2 s: W1 remained on the screen for 1 s, W2 until the subject responded by pressing a keyboard button, in any case not longer than 5 s [17,21]. Word pairs were used for three tasks in separated blocks, and the same words were presented as W1 in a different randomized order (Figure 5).
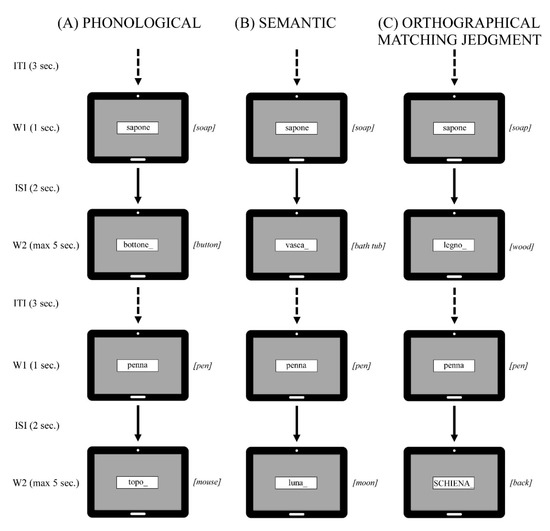
Figure 5.
Graphical representation of stimuli and tasks showing trial structure with first word presented (W1), Inter-Stimulus Interval (ISI), second word (W2) and Inter-Trial Interval (ITI) during (A) Phonological, (B) Semantic and (C) Orthographical Matching Judgment task. An example of matching (top panel) and mismatching (bottom panel) conditions is represented.
On the phonological task, participants had to decide, upon W2 presentation, whether the word pairs rhymed (e.g., butter-cutter) or not (e.g., butter-pencil); on the semantic categorization task, whether W2 was semantically related to the first (e.g., butter-bread) or not (e.g., butter-shoes); and on the orthographical matching judgment task, whether the each word in the word pair was written in the same case (e.g., butter-home or BUTTER-HOME) or not (e.g., butter-HOME). This last served as a control task, because no linguistic processing is requested. Participants pressed the button corresponding to letters “S” and “D” of a standard qwerty keyboard with the index or middle finger of their left hand to indicate their responses. Each task included 80 trials/word pairs: on all tasks, 50% matches were randomly interspersed with 50% mismatch trials. This experimental paradigm has been widely validated, particularly considering W1-evoked potentials, in both Italian samples of different ages [17,18,19], in clinical samples [20,22], and Italian vs. German monolingual individuals [23].
4.4. Data Analysis
Error rates and mean response times served as behavioral measures of task performance. Procedure, data recording, and analysis were carried out following the main international Guidelines of the EEG-ERPs field [38,39].
EEG was continuously recorded in the DC mode with Cz as an on-line reference, and stored for analysis. Data were off-line re-referenced to the average reference, in agreement with all past studies we carried out, using the same experimental paradigm [17,18,19,20,21,22]. Due to the aims of the present study, ERP analyses focused on W2 stimuli. Therefore, we cut epochs around 500 ms before and 1000 ms after W2 onset, and a 100-msec baseline preceding W2 was subtracted from the whole trial epoch. After eye movement artifacts correction [40,41], using BESA software (Brain Electrical Source Analysis, 5.1 version), each trial was visually inspected for any remaining artifacts, which were rejected. Trials corresponding to wrong behavioral responses were rejected as well. All accepted trials (67.4% ± 2.7% and 68.2% ± 2.4% for match and mismatch condition during Phonological task, 65.7 ± 5.2% and 65.9 ± 4.7% for match and mismatch condition during Semantic task, and 68.5 ± 2.2% and 69.2 ± 1.7% for match and mismatch condition during Orthographical Matching Judgment task) were averaged for each task and for each subject. As a final step, a 30 Hz filter was applied. After visual inspection of waveforms (Figure 6), three epochs were entered in the statistical comparisons, corresponding to the word RP, peaking 110–130 ms after W2 onset [17,19,20,22], the N400 component (300–450 ms; e.g., [3,4]) and the late N400 (450–600 ms) after W2 onset.
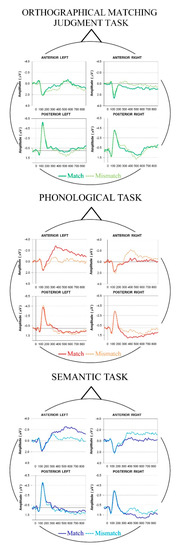
Figure 6.
Mean grand-average waveforms of all regions of interest showing the time-course of early linguistic processing following W2 onset during Orthographical Matching Judgment (top panel, in green), Phonological (middle panel, in red), and Semantic task (bottom panel, in blue) for Match (full line) and Mismatch (dotted line) trials. Negativity is displayed upward.
Mean values of the potential measured in the whole selected windows were used for statistical analysis. Electrodes were clustered into four groups/regions-of-interest to perform statistical analysis with two spatial factors of two levels each: Region and Laterality. Each quadrant comprised 4 electrodes: anterior left (AL: F3, FC3, F7, FT7), anterior right (AR: F4, FC4, F8, FT8), posterior left (PL: P3, P7, TP7, O1), and posterior right (PR: P4, P8, TP8, O2).
For each time window, the ANOVA included the following variables: task (three levels: orthographical matching judgment vs. phonological vs. semantic), condition (two levels: match vs. mismatch), region (two levels: anterior vs. posterior), and laterality (two levels: left vs. right side). Post hoc comparisons were computed using the Tukey HSD test (p < 0.05), and the Huynh-Feldt correction was applied when necessary (that is, when variables with more than two levels, e.g., task factor, were involved [42]).
Finally, Pearson’s correlation analyses were carried out between laterality indices obtained during task processing and the corresponding behavioral results obtained after task execution, to ascertain whether hemispheric lateralization represented an earlier physiological correlate significantly associated with later behavioral performance. Due to the aim of the present study, match and mismatch conditions were considered for physiological and behavioral measures. Laterality indices were computed as the difference of the mean activity of left minus right electrodes on RP posterior sites and N400/late N400 anterior sites. Thus, laterality indices were negative when participants had greater negative amplitude in the left hemisphere and positive when they had greater negativity in the right hemisphere.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-8994/13/1/74/s1. Table S1: Statistical analysis on RTs during W1-W2 Phonological, Semantic and Orthographical Matching Judgment tasks. Table S2: Statistical analysis on Errors (%) during W1-W2 Phonological, Semantic and Orthographical Matching Judgment tasks. Table S3: Statistical analysis on word RP (110–130 ms) after W2 onset. Table S4: Pearson’s correlations on word RP (110–130 ms) laterality indices (posterior sites only) and RTs. Table S5: Statistical analysis on N400 component (300–450 ms) after W2 onset. Table S6: Pearson’s correlations on N400 (300–450 ms) laterality indices (anterior sites only) and RTs. Table S7: Statistical analysis on late N400 component (450–600 ms) after W2 onset. Table S8: Pearson’s correlations on late N400 (450–600 ms) laterality indices (anterior sites only) and RTs.
Author Contributions
Conceptualization, investigation, methodology, validation and project administration: C.S. and A.A.; formal analysis, writing--original draft preparation and funding acquisition: C.S.; data curation, supervision and writing--review & editing: A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by PRAT 2015 from the University of Padova, project n. CPDA152872 to C.S. The present work was also carried out within the scope of the research program Dipartimenti di Eccellenza (art.1, commi 314-337 legge 232/2016), which was supported by a grant from MIUR to the Department of General Psychology, University of Padua.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of Psychology Area (protocol code 1240, date of approval 21 February 2013).
Informed Consent Statement
Informed consent was obtained from all participants involved in the study.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author. The data are not publicly available, due to privacy or ethical restrictions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kutas, M.; Hillyard, S.A. Reading senseless sentences: Brain potentials reflect semantic incongruity. Science 1980, 207, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Van Petten, C.; Luka, B.J. Neural localization of semantic context effects in electromagnetic and hemo-dynamic studies. Brain Lang. 2006, 97, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.F.; Phillips, C.; Poeppel, D. A cortical network for semantics: [de]constructing the N400. Nat. Rev. Neurosci. 2008, 9, 920–933. [Google Scholar] [CrossRef] [PubMed]
- Kutas, M.; Federmeier, K.D. Thirty Years and Counting: Finding Meaning in the N400 Component of the Event-Related Brain Potential (ERP). Annu. Rev. Psychol. 2011, 62, 621–647. [Google Scholar] [CrossRef] [PubMed]
- De Vincenzi, M.; Job, R.; Di Matteo, R.; Angrilli, A.; Penolazzi, B.; Ciccarelli, L.; Vespignani, F. Differences in the perception and time course of syntactic and semantic violations. Brain Lang. 2003, 85, 280–296. [Google Scholar] [CrossRef]
- Angrilli, A.; Penolazzi, B.; Vespignani, F.; De Vincenzi, M.; Job, R.; Ciccarelli, L.; Palomba, D.; Stegagno, L. Cortical brain responses to semantic incongruity and syntactic violation in Italian language: An Event-related Potential study. Neurosci. Lett. 2002, 322, 5–8. [Google Scholar] [CrossRef]
- Kutas, M.; Hillyard, S.A. The lateral distribution of event-related potentials during sentence processing. Neuropsychologia 1982, 20, 579–590. [Google Scholar] [CrossRef]
- Coulson, S.; Federmeier, K.D.; Van Petten, C.; Kutas, M. Right hemisphere sensitivity to word and sentence level context: Evidence from event-related brain potentials. J. Exp. Psychol. Learn. Mem. Cogn. 2005, 31, 129–147. [Google Scholar] [CrossRef]
- Coch, D. The N400 and the fourth grade shift. Dev. Sci. 2015, 18, 254–269. [Google Scholar] [CrossRef]
- Lau, E.F.; Holcomb, P.J.; Kuperberg, G.R. Dissociating N400 effects of prediction from association in single-word contexts. J. Cogn. Neurosci. 2013, 25, 484–502. [Google Scholar] [CrossRef]
- Rebai, M.; Bernard, C.; Lannou, J. The Stroop’s test evokes a negative brain potential, the N400. Int. J. Neurosci. 1997, 91, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Van Petten, C.; Rheinfelder, H. Conceptual relationships between spoken words and environmental sounds: Event-related brain potential measures. Neuropsychologia 1995, 33, 485–508. [Google Scholar] [CrossRef]
- Curran, T.; Tucker, D.M.; Kutas, M.; Posner, M.I. Topography of the N400: Brain electrical activity reflecting semantic expectancy. Electroencephalogr. Clin. Neurophysiol. 1993, 88, 188–209. [Google Scholar] [CrossRef]
- Johnson, B.W.; Hamm, J.P. High-density mapping in an N400 paradigm: Evidence for bilateral temporal lobe generators. Clin. Neurophysiol. 2000, 111, 532–545. [Google Scholar] [CrossRef]
- Frishkoff, G.A.; Tucker, D.M.; Davey, C.; Scherg, M. Frontal and posterior sources of event- related potentials in semantic comprehension. Brain Res. Cogn. Brain Res. 2004, 20, 329–354. [Google Scholar] [CrossRef]
- Kutas, M.; Hillyard, S.A.; Gazzaniga, M. Processing of semantic anomaly by right and left hemispheres of commissurotomy patients. Brain 1988, 111, 553–576. [Google Scholar] [CrossRef]
- Spironelli, C.; Angrilli, A. Influence of Phonological, Semantic and Orthographic tasks on the early linguistic components N150 and N350. Int. J. Psychophysiol. 2007, 64, 190–198. [Google Scholar] [CrossRef]
- Spironelli, C.; Angrilli, A. Developmental aspects of automatic word processing: Language lateralization of early ERP components in children, young adults and middle-aged subjects. Biol. Psychol. 2009, 80, 35–45. [Google Scholar] [CrossRef]
- Spironelli, C.; Penolazzi, B.; Angrilli, A. Gender differences in reading in school-aged children: An early ERP study. Dev. Neuropsychol. 2010, 35, 357–375. [Google Scholar] [CrossRef]
- Spironelli, C.; Penolazzi, B.; Vio, C.; Angrilli, A. Cortical reorganization in dyslexic children after a phonological training: Evidence from early evoked potentials. Brain 2010, 133, 3385–3395. [Google Scholar] [CrossRef]
- Spironelli, C.; Angrilli, A. Language lateralization in phonological, semantic and orthographic tasks: A slow evoked potential study. Behav. Brain Res. 2006, 175, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Spironelli, C.; Angrilli, A.; Stegagno, L. Failure of language lateralization in schizophrenia patients: An ERP study on early linguistic components. J. Psychiatry Neurosci. 2008, 33, 235–243. [Google Scholar] [PubMed]
- Angrilli, A.; Dobel, C.; Rockstroh, B.S.; Stagagno, L.; Elbert, T. EEG brain mapping of phonological and semantic tasks in Italian and German languages. Clin. Neurophysiol. 2000, 111, 706–716. [Google Scholar] [CrossRef][Green Version]
- Bless, J.J.; Westerhausen, R.; von Koss Torkildsen, J.; Gudmundsen, M.; Kompus, K.; Hugdahl, K. Laterality across languages: Results from a global dichotic listening study using a smartphone application, Laterality: Asymmetries of Body. Brain Cogn. 2015, 20, 434–452. [Google Scholar]
- Bookheimer, S. Functional MRI of language: New approaches to understanding the cortical organization of semantic processing. Annu. Rev. Neurosci. 2002, 25, 151–188. [Google Scholar] [CrossRef]
- Hagoort, P. On Broca, brain, and binding: A new framework. TRENDS Cogn. Sci. 2005, 9, 416–423. [Google Scholar] [CrossRef]
- Ardal, S.; Donald, M.W.; Meuter, R.; Muldrew, S.; Luce, M. Brain responses to semantic incongruity in bilinguals. Brain Lang. 1990, 39, 187–205. [Google Scholar] [CrossRef]
- Rämä, P.; Sirri, L.; Serres, J. Development of lexical–semantic language system: N400 priming effect for spoken words in 18- and 24-month old children. Brain Lang. 2013, 125, 1–10. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Felton, A.; Vazquez, D.; Ramos-Nunez, A.I.; Greene, M.R.; McDowell, A.; Hernandez, A.E.; Chiarello, C. Bilingualism influences structural indices of interhemispheric organization. J. Neuroling. 2017, 42, 1–11. [Google Scholar] [CrossRef]
- Huang, K.; Itoh, K.; Kwee, I.L.; Nakada, T. Neural strategies for reading Japanese and Chinese sentences: A cross-linguistic fMRI study of character-decoding and morphosyntax. Neuropsychologia 2012, 50, 2598–2604. [Google Scholar] [CrossRef] [PubMed]
- Klichowski, M.; Kroliczak, G. Numbers and functional lateralization: A visual half-field and dichotic listening study in proficient bilinguals. Neuropsychologia 2017, 100, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Klichowski, M.; Nowik, A.; Kroliczak, G.; Lewis, J.W. Functional lateralization of tool-sound and action-word processing in a bilingual brain. Health Psychol.Rep. 2020, 8, 10–30. [Google Scholar] [CrossRef]
- Krefta, M.; Michalowski, B.; Kowalczyk, J.; Kroliczak, G. Co-lateralized bilingual mechanisms for reading in single and dual language contexts: Evidence from visual half-field processing of action words in proficient bilinguals. Front. Psychol. 2015, 6, 1159. [Google Scholar] [CrossRef] [PubMed]
- Park, H.R.; Badzakova-Trajkov, G.; Waldie, K.E. Language lateralisation in late proficient bilinguals: A lexical decision fMRI study. Neuropsychologia 2012, 50, 688–695. [Google Scholar] [CrossRef]
- Oostenveld, R.; Praamstra, P. The five percent electrode system for high-resolution EEG and ERP measurements. Clin. Neurophysiol. 2001, 112, 713–719. [Google Scholar] [CrossRef]
- Bortolini, V.; Tagliavini, C.; Zampolli, A. Lessico di Frequenza Della Lingua Italiana Contemporanea [Lexical Frequency in Current Italian]; Aldo Garzanti Editore SpA: Milano, Italy, 1972. [Google Scholar]
- Keil, A.; Debener, S.; Gratton, G.; Junghöfer, M.; Kappenman, E.S.; Luck, S.J.; Luu, P.; Miller, G.A.; Yee, C.M. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology 2014, 51, 1–21. [Google Scholar] [CrossRef]
- Picton, T.W.; Bentin, S.; Berg, P.; Donchin, E.; Hillyard, S.A.; Johnson, R., Jr.; Miller, G.A.; Ritter, W.; Ruchkin, D.S.; Rugg, M.D.; et al. Guidelines for using human event-related potentials to study cognition: Recording standards and publication criteria. Psychophysiology 2000, 37, 127–152. [Google Scholar] [CrossRef]
- Berg, P.; Scherg, M. Dipole models of eye movements and blinks. Clin. Neurophysiol. 1991, 79, 36–44. [Google Scholar] [CrossRef]
- Berg, P.; Scherg, M. A multiple source approach to the correction of eye artifacts. Clin. Neurophysiol. 1994, 90, 229–241. [Google Scholar] [CrossRef]
- Huynh, H.; Feldt, L.S. Condition under which mean square rations in repeated measurements designs have exact F-distributions. J. Am. Stat. Assoc. 1970, 65, 1582–1589. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).