
Insect Fluctuating Asymmetry: An Example in Bolivian Peridomestic Populations of Triatoma infestans (Klug, 1834) (Hemiptera: Reduviidae)

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
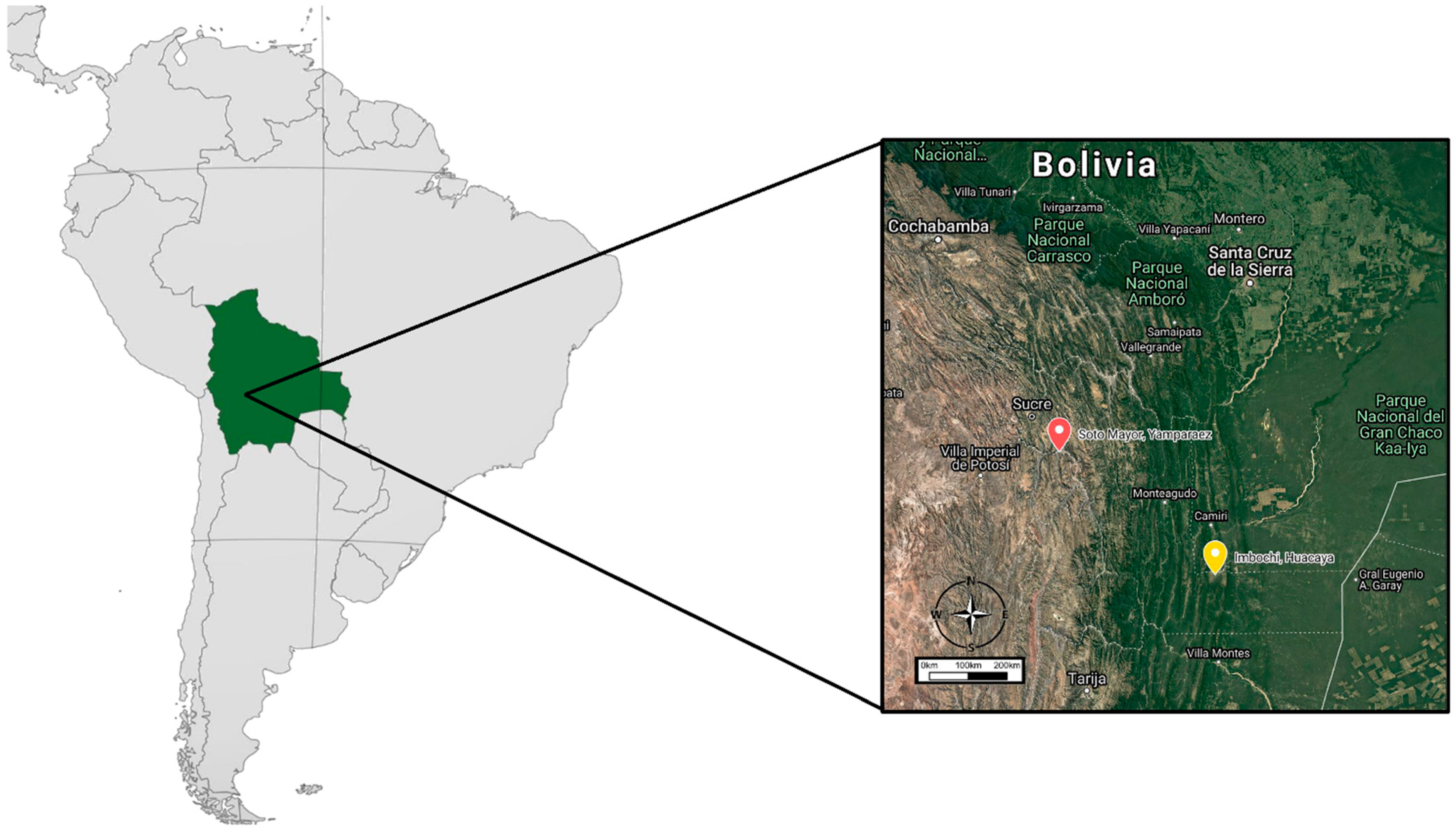
2.1. Study Area
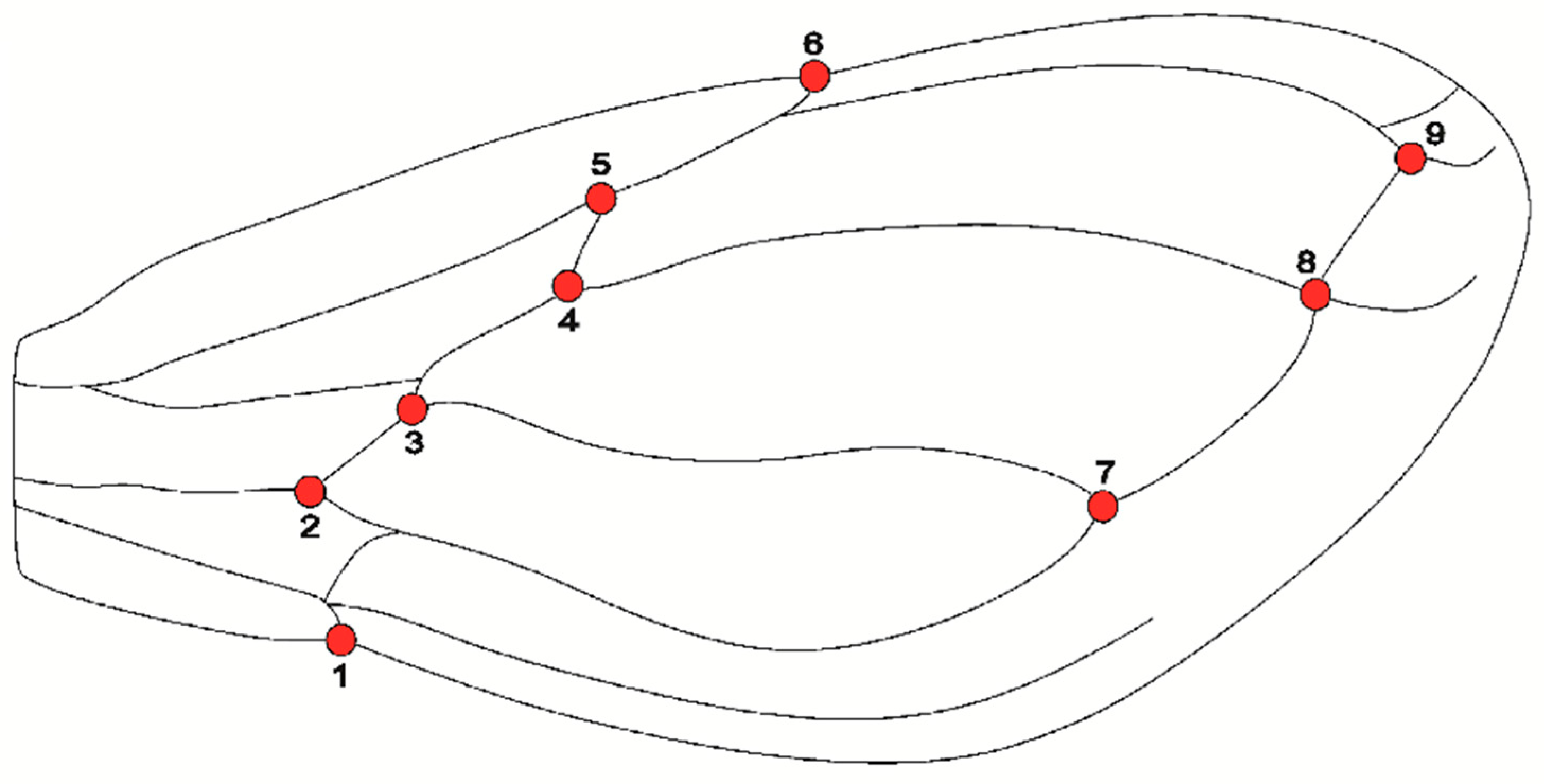
2.2. Insect Sampling and Preparation
2.3. Shape Analyses
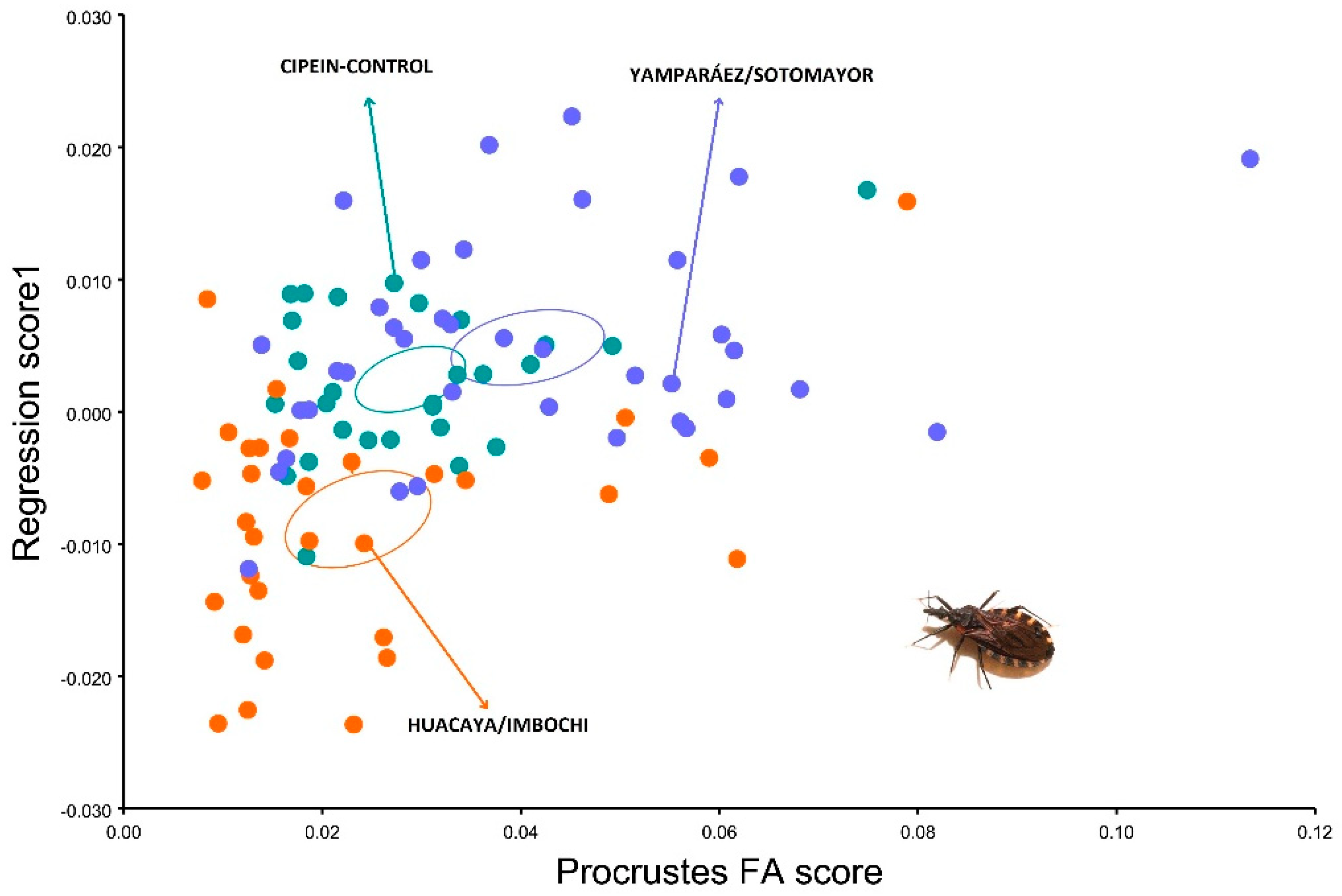
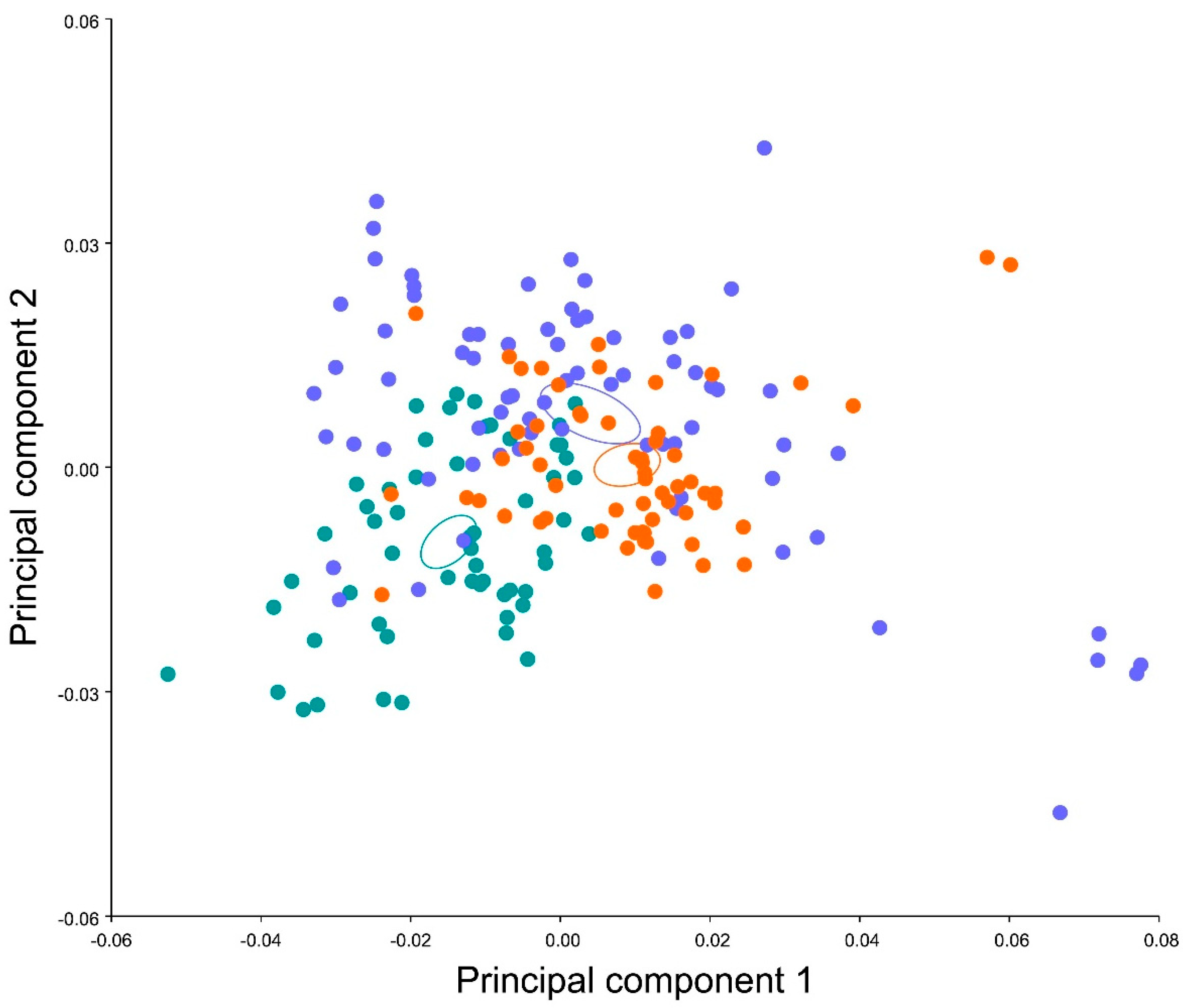
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noireau, F.; Carbajal-de-La-Fuente, A.L.; Lopes, C.M.; Diotaiuti, L. Some considerations about the ecology of Triatominae. An. Acad. Bras. Ciências 2005, 77, 431–436. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Noireau, F.; Cortez, M.G.R.; Monteiro, F.A.; Jansen, A.M.; Torrico, F. Can wild Triatoma infestans foci in Bolivia jeopardize Chagas disease control efforts? Trends Parasitol. 2005, 21, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Richer, W.; Kengne, P.; Cortez, M.R.; Perrineau, M.M.; Cohuet, A.; Fontenille, D.; Noireau, F. Active dispersal by wild Triatoma infestans in the Bolivian Andes. Trop. Med. Int. Health 2007, 12, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, L.; Vazquez-Prokopec, G.; Cecere, M.; Marcet, P.; Gürtler, R. Feeding rates, nutritional status and flight dispersal potential of peridomestic populations of Triatomainfestans in rural northwestern Argentina. Acta Trop. 2005, 95, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Cecere, M.C.; Vazquez-Prokopec, G.M.; Gürtler, R.E.; Kitron, U. Spatio-temporal analysis of reinfestation by Triatoma infestans (Hemiptera: Reduviidae) following insecticide spraying in a rural community in northwestern Argentina. Am. J. Trop. Med. Hyg. 2004, 71, 803. [Google Scholar] [CrossRef]
- Gürtler, R.E.; Cecere, M.C.; Vázquez-Prokopec, G.M.; Ceballos, L.A.; Gurevitz, J.M.; Fernández, M.D.P.; Kitron, U.; Cohen, J.E. Domestic animal hosts strongly influence human-feeding rates of the Chagas disease vector Triatoma infestans in Argentina. PLoS Negl. Trop. Dis. 2014, 8, e2894. [Google Scholar] [CrossRef]
- Herrera, L. Trypanosoma cruzi, the causal agent of Chagas disease: Boundaries between wild and domestic cycles in Venezuela. Front. Public Health 2014, 2, 259. [Google Scholar] [CrossRef]
- Kirchhoff, L.V. Chagas disease: American trypanosomiasis. Infect. Dis. Clin. N. Am. 1993, 7, 487–502. [Google Scholar] [CrossRef]
- World Health Organization. Chagas disease (American trypanosomiasis)—Factsheet (revised in August 2012). Wkly. Epidemiol. Rec. Relev. Épidémiologique Hebd. 2012, 87, 519–522. [Google Scholar]
- Vilaseca, C.; Méndez, M.A.; Pinto, C.F.; Lemic, D.; Benítez, H.A. Unraveling the Morphological Variation of Triatoma infestans in the Peridomestic Habitats of Chuquisaca Bolivia: A Geometric Morphometric Approach. Insects 2021, 12, 185. [Google Scholar] [CrossRef]
- Chuquisaca, P.C. Informe Anual de Índices de Infestaciónpor Triatoma infestans en el Intra y peri Domicilio en Chuquisaca; Ministerio de Salud y Deportes de Bolivia: Sucre, Bolivia, 2017.
- Gonzalez-Valdivieso, F.E.; Sanchez Diaz, B.; Nocerino, F. Susceptibility of R. prolixux to Chlorinated Hydrocarbon Insecticides in Venezuela; World Health Organization: Geneva, Switzerland, 1971. [Google Scholar]
- Picollo, M.I.; Vassena, C.; Orihuela, P.S.; Barrios, S.; Zaidemberg, M.; Zerba, E. High resistance to pyrethroid insecticides associated with ineffective field treatments in Triatoma infestans (Hemiptera: Reduviidae) from Northern Argentina. J. Med. Entomol. 2005, 42, 637–642. [Google Scholar] [CrossRef]
- Acevedo, G.R.; Cueto, G.M.; Germano, M.; Orihuela, P.S.; Cortez, M.R.; Noireau, F.; Picollo, M.I.; Vassena, C. Susceptibility of sylvatic Triatoma infestans from Andeans valleys of Bolivia to deltamethrin and fipronil. J. Med. Entomol. 2011, 48, 828–835. [Google Scholar] [CrossRef]
- Santo-Orihuela, P.L.; Carvajal, G.; Picollo, M.I.; Vassena, C.V. Toxicological and biochemical analysis of the susceptibility of sylvatic Triatoma infestans from the Andean Valley of Bolivia to organophosphate insecticide. Memórias Inst. Oswaldo Cruz 2013, 108, 790–795. [Google Scholar] [CrossRef]
- Germano, M.; Santo-Orihuela, P.; Roca-Acevedo, G.; Toloza, A.; Vassena, C.; Picollo, M.; Mougabure-Cueto, G. Scientific evidence of three different insecticide-resistant profiles in Triatoma infestans (Hemiptera: Reduviidae) populations from Argentina and Bolivia. J. Med. Entomol. 2014, 49, 1355–1360. [Google Scholar] [CrossRef]
- Lardeux, F.; Depickère, S.; Duchon, S.; Chavez, T. Insecticide resistance of Triatoma infestans (Hemiptera, Reduviidae) vector of Chagas disease in Bolivia. Trop. Med. Int. Health 2010, 15, 1037–1048. [Google Scholar] [CrossRef]
- Santo-Orihuela, P.L.; Vassena, C.V.; Carvajal, G.; Clark, E.; Menacho, S.; Bozo, R.; Gilman, R.H.; Bern, C.; Marcet, P.L. Toxicological, enzymatic, and molecular assessment of the insecticide susceptibility profile of Triatoma infestans (Hemiptera: Reduviidae, Triatominae) populations from rural communities of Santa Cruz, Bolivia. J. Med. Entomol. 2017, 54, 187–195. [Google Scholar] [CrossRef]
- Depickère, S.; Buitrago, R.; Siñani, E.; Baune, M.; Monje, M.; Lopez, R.; Waleckx, E.; Chavez, T.; Brenière, S.F. Susceptibility and resistance to deltamethrin of wild and domestic populations of Triatoma infestans (Reduviidae: Triatominae) in Bolivia: New discoveries. Memórias Inst. Oswaldo Cruz 2012, 107, 1042–1047. [Google Scholar] [CrossRef]
- Vilaseca, C.; Méndez, M.A.; Pinto, C.F.; Benítez, H.A. Assessment of Shape Variation Patterns in Triatoma infestans (Klug 1834) (Hemiptera: Reduviidae: Triatominae): A First Report in Populations from Bolivia. Insects 2020, 11, 274. [Google Scholar] [CrossRef]
- Parsons, P. Fluctuating asymmetry: A biological monitor of environmental and genomic stress. Heredity 1992, 68, 361–364. [Google Scholar] [CrossRef]
- Abaga, N.O.Z.; Alibert, P.; Dousset, S.; Savadogo, P.W.; Savadogo, M.; Sedogo, M. Insecticide residues in cotton soils of Burkina Faso and effects of insecticides on fluctuating asymmetry in honey bees (Apis mellifera Linnaeus). Chemosphere 2011, 83, 585–592. [Google Scholar] [CrossRef]
- Benítez, H.A.; Briones, R.; Jerez, V. Intra and Inter-population morphological variation of shape and size of the Chilean magnificent beetle, Ceroglossus chilensis in the Baker River Basin, Chilean Patagonia. J. Insect Sci. 2011, 11, 1–9. [Google Scholar] [CrossRef][Green Version]
- Chang, X.; Zhai, B.; Wang, M.; Wang, B. Relationship between exposure to an insecticide and fluctuating asymmetry in a damselfly (Odonata, Coenagriidae). Hydrobiologia 2007, 586, 213–220. [Google Scholar] [CrossRef]
- Ribeiro, B.; Guedes, R.; Corrêa, A.; Santos, C. Fluctuating asymmetry in insecticide-resistant and insecticide-susceptible strains of the maize weevil, Sitophilus zeamais (Coleoptera: Curculionidae). Arch. Environ. Contam. Toxicol. 2007, 53, 77–83. [Google Scholar] [CrossRef]
- Benítez, H.; Briones, R.; Jerez, V. Fluctuating asymmetry in two populations of Ceroglossus chilensis (Eschscholtz, 1829) (Coleoptera: Carabidae) in agroecosystem of Pinus radiata d. Don, Bio-Bio region, Chile. Gayana 2008, 72, 131–139. [Google Scholar]
- Benítez, H.A.; Lemic, D.; Villalobos-Leiva, A.; Bažok, R.; Órdenes-Claveria, R.; Pajač Živković, I.; Mikac, K.M. Breaking Symmetry: Fluctuating Asymmetry and Geometric Morphometrics as Tools for Evaluating Developmental Instability under Diverse Agroecosystems. Symmetry 2020, 12, 1789. [Google Scholar] [CrossRef]
- Ivanković Tatalović, L.; Anđelić, B.; Jelić, M.; Kos, T.; Benítez, H.A.; Šerić Jelaska, L. Fluctuating Asymmetry as a Method of Assessing Environmental Stress in Two Predatory Carabid Species within Mediterranean Agroecosystems. Symmetry 2020, 12, 1890. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry analyses revisited. In Developmental Instability (DI): Causes and Consequences; Oxford University Press: Oxford, UK, 2003; pp. 279–319. [Google Scholar]
- Brouwer, A.; Murk, A.; Koeman, J. Biochemical and physiological approaches in ecotoxicology. Funct. Ecol. 1990, 4, 275–281. [Google Scholar] [CrossRef]
- Eeva, T.; Tanhuanpaa, S.; Rabergh, C.; Airaksinen, S.; Nikinmaa, M.; Lehikoinen, E. Biomarkers and fluctuating asymmetry as indicators of pollution-induced stress in two hole-nesting passerines. Funct. Ecol. 2000, 14, 235–243. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating Asymmetry—Measurement, Analysis, Patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- De Coster, G.; Van Dongen, S.; Malaki, P.; Muchane, M.; Alcántara-Exposito, A.; Matheve, H.; Lens, L. Fluctuating Asymmetry and Environmental Stress: Understanding the Role of Trait History. PLoS ONE 2013, 8, e57966. [Google Scholar] [CrossRef]
- Benítez, H.A.; Lemic, D.; Bazok, R.; Gallardo-Araya, C.M.; Mikac, K.M. Evolutionary directional asymmetry and shape variation in Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae): An example using hind wings. Biol. J. Linn. Soc. 2014, 111, 110–118. [Google Scholar] [CrossRef]
- Graham, J.H.; Emlen, J.M.; Freeman, D.C.; Leamy, L.J.; Kieser, J.A. Directional asymmetry and the measurement of developmental instability. Biol. J. Linn. Soc. 1998, 64, 1–16. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; Barluenga, M.; Meyer, A. Shape analysis of symmetric structures: Quantifying variation among individuals and asymmetry. Evolution 2002, 56, 1909–1920. [Google Scholar] [CrossRef]
- Klingenberg, C. Analyzing Fluctuating Asymmetry with Geometric Morphometrics: Concepts, Methods, and Applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef]
- Benítez, H.A.; Vargas, H.A.; Püschel, T.A. Left–right asymmetry and morphological consequences of a host shift in the oligophagous Neotropical moth Macaria mirthae (Lepidoptera: Geometridae). J. Insect Conserv. 2015, 19, 589–598. [Google Scholar] [CrossRef]
- Nunes, L.A.; Araújo, E.D.D.; Marchini, L.C. Fluctuating asymmetry in Apis mellifera (Hymenoptera: Apidae) as bioindicator of anthropogenic environments. Rev. Biol. Trop. 2015, 63, 673–682. [Google Scholar] [CrossRef][Green Version]
- Nattero, J.; Piccinali, R.V.; Gaspe, M.S.; Gürtler, R.E. Fluctuating asymmetry and exposure to pyrethroid insecticides in Triatoma infestans populations in northeastern Argentina. Infect. Genet. Evol. 2019, 74, 103925. [Google Scholar] [CrossRef]
- Nattero, J.; Dujardin, J.-P.; del Pilar Fernández, M.; Gürtler, R.E. Host-feeding sources and habitats jointly affect wing developmental stability depending on sex in the major Chagas disease vector Triatoma infestans. Infect. Genet. Evol. 2015, 36, 539–546. [Google Scholar] [CrossRef]
- Nattero, J.; Piccinali, R.V.; Lopes, C.M.; Hernández, M.L.; Abrahan, L.; Lobbia, P.A.; Rodríguez, C.S.; de la Fuente, A.L.C. Morphometric variability among the species of the Sordida subcomplex (Hemiptera: Reduviidae: Triatominae): Evidence for differentiation across the distribution range of Triatoma sordida. Parasites Vectors 2017, 10, 412. [Google Scholar] [CrossRef]
- Di Iorio, O.; Gürtler, R.E. Seasonality and temperature-dependent flight dispersal of Triatoma infestans (Hemiptera: Reduviidae) and other vectors of Chagas disease in western Argentina. J. Med. Entomol. 2017, 54, 1285–1292. [Google Scholar] [CrossRef]
- Navarro, G.; Maldonado, M. Geografía Ecológica de Bolivia: Vegetación y Ambientes Acuáticos; Centro de Ecología Simón I. Patiño, Departamento de Difusión: Cochabamba, Bolivia, 2002. [Google Scholar]
- Rohlf, F.J. TPSdig; Version 2.17; State University at Stony Brook: Stony Brook, NY, USA, 2013. [Google Scholar]
- Rohlf, F.J.; Slice, D. Extensions of the Procustes methods for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; McIntyre, G.S. Geometric morphometrics of developmental instability: Analyzing patterns of fluctuating asymmetry with procrustes methods. Evolution 1998, 52, 1363–1375. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; McIntyre, G.S.; Zaklan, S.D. Left-right asymmetry of fly wings and the evolution of body axes. Proc. R. Soc. B-Biol. Sci. 1998, 265, 1255–1259. [Google Scholar] [CrossRef]
- Palmer, A.R. Fluctuating asymmetry analyses: A primer. In Developmental Instability: Its Origins and Evolutionary Implications; Springer: Berlin/Heidelberg, Geramny, 1994; pp. 335–364. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Nijhout, H.; Davidowitz, G. Developmental perspectives on phenotypic variation, canalization, and fluctuating asymmetry. In Developmental Instability: Causes and Consequences; Oxford University Press: Oxford, UK, 2003; pp. 3–13. [Google Scholar]
- Van Dongen, S. Fluctuating asymmetry and developmental instability in evolutionary biology: Past, present and future. J. Evol. Biol. 2006, 19, 1727–1743. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Developmental instability as a research tool: Using patterns of fluctuating asymmetry to infer the developmental origins of morphological integration. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: New York, NY, USA, 2003; pp. 427–442. [Google Scholar]
- Polak, M. Developmental Instability: Causes and Consequences; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Benítez, H.A.; Parra, L.E. Asimetría fluctuante: Una herramienta morfo-funcional para medir estabilidad del desarrollo. Int. J. Morphol. 2011, 29, 1459–1469. [Google Scholar] [CrossRef]
- Parsons, P. Fluctuating asymmetry: An epigenetic measure of stress. Biol. Rev. Camb. Philos. Soc. 1990, 65, 131–145. [Google Scholar] [CrossRef]
- Benítez, H. Assessment of patterns of fluctuating asymmetry and sexual dimorphism in carabid body shape. Neotrop. Entomol. 2013, 42, 164–169. [Google Scholar] [CrossRef]
- Benítez, H.A.; Lemic, D.; Püschel, T.A.; Gašparić, H.V.; Kos, T.; Barić, B.; Bažok, R.; Živković, I.P. Fluctuating asymmetry indicates levels of disturbance between agricultural productions: An example in Croatian population of Pterostichus melas melas (Coleptera: Carabidae). Zool. Anz. 2018, 276, 42–49. [Google Scholar] [CrossRef]
- Woods, R.E.; Sgrò, C.M.; Hercus, M.J.; Hoffmann, A.A. The association between fluctuating asymmetry, trait variability, trait heritability, and stress: A multiply replicated experiment on combined stresses in Drosophila melanogaster. Evolution 1999, 53, 493–505. [Google Scholar] [CrossRef]
- Vazquez-Prokopec, G.M.; Ceballos, L.A.; Kitron, U.; Gürtler, R.E. Active dispersal of natural populations of Triatoma infestans (Hemiptera: Reduviidae) in rural northwestern Argentina. J. Med. Entomol. 2004, 41, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Lazzari, C.R. Temperature preference in Triatoma infestans (Hemiptera: Reduviidae). Bull. Entomol. Res. 1991, 81, 273–276. [Google Scholar] [CrossRef]
- Lazzari, C.; Pereira, M.; Lorenzo, M. Biología del comportamiento de los vectores de la enfermedad de Chagas. Mem. Inst. Oswaldo Cruz 2013, 108, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Roca, M.J.; Lazzari, C.R. Effects of relative humidity on the haematophagous bug Triatoma infestans: Hygropreference and eclosion success. J. Insect Physiol. 1994, 40, 901–907. [Google Scholar] [CrossRef]
- Cecere, M.C.; Gürtler, R.E.; Canale, D.; Chuit, R.; Cohen, J.E. El papel del peridomicilio en la eliminación de Triatoma infestans de comunidades rurales argentinas. Bol. Sanit. Panam. 1996, 121, 1–10. [Google Scholar]
- De Villar, M.; Zerba, E.; Wood, E.; De Licastro, S. Neurogenesis and occurrence of cholinesterase in eggs of Triatoma infestans. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1980, 65, 65–70. [Google Scholar] [CrossRef]
- Leamy, L.J.; Klingenberg, C.P. The genetics and evolution of fluctuating asymmetry. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 1–21. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Centroid Size/Effect | SS | MS | df | F | p | Pillai tr. | p (Param) |
|---|---|---|---|---|---|---|---|
| Individual | 1,267,588 | 0.469477 | 27 | 12.13 | <0.0001 | ||
| Side | 1,312,120 | 131.212 | 1 | 33.90 | <0.0001 | ||
| Ind*Side | 1,044,953 | 0.038702 | 27 | 1.85 | 0.026 | ||
| Error 1 | 116,915 | 0.020888 | 56 | ||||
| Shape/Effect | |||||||
| Individual | 0.06646813 | 0.000175842 | 378 | 2.52 | <0.0001 | 9.38 | <0.0001 |
| Side | 0.00176474 | 0.000126003 | 14 | 1.81 | 0.0358 | 0.76 | 0.0203 |
| Ind*Side | 0.0263932 | 0.000697337 | 378 | 3.08 | <0.0001 | 7.57 | <0.0001 |
| Error | 0.0177762 | 0.0000226737 | 784 |
| Centroid Size/Effect | SS | MS | df | F | p | Pillai tr. | p (Param) |
|---|---|---|---|---|---|---|---|
| Individual | 46,357,914 | 1,252,917 | 37 | 4.99 | <0.0001 | ||
| Side | 1,439,198 | 1,439,198 | 1 | 5.73 | 0.0218 | ||
| Ind*Side | 9,286,172 | 0.250978 | 37 | 20.33 | <0.0001 | ||
| Error 1 | 0.938393 | 0.012347 | 76 | ||||
| Shape/Effect | |||||||
| Individual | 0.20439244 | 0.0003945800 | 518 | 2.01 | <0.0001 | 8.81 | <0.0001 |
| Side | 0.00641765 | 0.0004584037 | 14 | 2.34 | 0.0039 | 0.64 | 0.008 |
| Ind*Side | 0.10166526 | 0.0001962650 | 518 | 2.61 | <0.0001 | 9.17 | <0.0001 |
| Error | 0.08004337 | 0.000075287 | 1064 |
| Centroid Size/Effect | SS | MS | df | F | p | Pillai tr. | p (Param) |
|---|---|---|---|---|---|---|---|
| Individual | 661,088,646,217 | 2,203,628,821 | 30 | 1.00 | 0.4993 | ||
| Side | 220,987,792 | 220,987,792 | 1 | 1.00 | 0.3245 | ||
| Ind*side | 6,606,556,014 | 2,202,185,338 | 30 | 1.00 | 0.4835 | ||
| Error 1 | 1,363,261,983 | 219,880,965 | 62 | ||||
| Shape/Effect | |||||||
| Individual | 0.10800141 | 0.000257146 | 420 | 4.09 | <0.0001 | 10.75 | <0.0001 |
| Side | 0.00132375 | 0.0000945536 | 14 | 1.5 | 0.1065 | 0.89 | <0.0001 |
| Ind*Side | 0.02643423 | 0.0000629386 | 420 | 5.28 | <0.0001 | 7.48 | <0.0001 |
| Error | 0.08004337 | 0.000075287 | 1064 |
| Population | Sex | N° of Insects | MS (Ind*Side) | Pillai tr. |
|---|---|---|---|---|
| Yamparáez/Sotomayor | Female | 14 | 0.0001970676 | 7.98 |
| Male | 24 | 0.0001892431 | 8.43 | |
| Huacaya/Imbochi | Female | 14 | 0.0000751902 | 7.05 |
| Male | 17 | 0.0000546154 | 6.22 | |
| CIPEIN | Female | 14 | 0.0000692699 | 7.18 |
| Male | 14 | 0.0000724677 | 7.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilaseca, C.; Pinto, C.F.; Órdenes-Claveria, R.; Laroze, D.; Méndez, M.A.; Benítez, H.A. Insect Fluctuating Asymmetry: An Example in Bolivian Peridomestic Populations of Triatoma infestans (Klug, 1834) (Hemiptera: Reduviidae). Symmetry 2022, 14, 526. https://doi.org/10.3390/sym14030526
Vilaseca C, Pinto CF, Órdenes-Claveria R, Laroze D, Méndez MA, Benítez HA. Insect Fluctuating Asymmetry: An Example in Bolivian Peridomestic Populations of Triatoma infestans (Klug, 1834) (Hemiptera: Reduviidae). Symmetry. 2022; 14(3):526. https://doi.org/10.3390/sym14030526
Chicago/Turabian StyleVilaseca, Carolina, Carlos F. Pinto, Rodrigo Órdenes-Claveria, David Laroze, Marco A. Méndez, and Hugo A. Benítez. 2022. "Insect Fluctuating Asymmetry: An Example in Bolivian Peridomestic Populations of Triatoma infestans (Klug, 1834) (Hemiptera: Reduviidae)" Symmetry 14, no. 3: 526. https://doi.org/10.3390/sym14030526
APA StyleVilaseca, C., Pinto, C. F., Órdenes-Claveria, R., Laroze, D., Méndez, M. A., & Benítez, H. A. (2022). Insect Fluctuating Asymmetry: An Example in Bolivian Peridomestic Populations of Triatoma infestans (Klug, 1834) (Hemiptera: Reduviidae). Symmetry, 14(3), 526. https://doi.org/10.3390/sym14030526