Abstract
In this paper, we carry out some research on a predator–prey system with maturation delay, a stage structure for generalist predators and a Holling type-II functional response, which has already been proposed. First, for the delayed model, we obtain the conditions for the occurrence of stability switches of the positive equilibrium and possible Hopf bifurcation values owing to the growth of the value of the delay by applying the geometric criterion. It should be pointed out that when we suppose that the characteristic equation has a pair of imaginary roots , we just need to consider due to the symmetry, which alleviates the computation requirements. Next, we investigate the nature of Hopf bifurcation. Finally, we conduct numerical simulations to verify the correctness of our findings.
1. Introduction
Interspecies relationships include competition, predation, parasitism and mutualism, where predation describes the interplay between the predator and prey. It is crucial to explore the interaction between them and then formulate an appropriate mathematical model to accurately capture the dynamics of the predator–prey model. The classic predator–prey model was first put forward by Lotka [1] and Volterra [2]. Since then, a lot of scholars have incorporated multifarious factors into the classic model to expound intricate biological processes in a more realistic way [3,4,5,6].
Since all biological processes take time to complete, one tends to take into account time delays in the modeling efforts [7,8,9,10]. This will produce delay differential equation models in most instances. Some of these models result from age structure models or elaborate formulations, which means that time lags often arise in the survival rates of the populations, such as [11]. Clearly, these models possess delay-dependent parameters, while most of delay differential equation models only have parameters independent of time delays [12,13,14,15,16].
In ecological research, stability analysis is a key task. Common stability analysis includes finite-time stability, exponential stability, local asymptotic stability and global asymptotic stability [17]. In this paper, we focus on analyzing the local asymptotic stability of the positive equilibrium. Studying the stability of the delayed system has aroused wide concern in mathematical biology since the time lag plays a surprisingly vital role in affecting dynamics of the model. It may make the model stable or unstable, relying on the length of the time delay [18]. Cooke and Grossman [12] first demonstrated that constructing a system with stability switches was possible. By combining graphical information with analytical work, Beretta and Kuang [18] proposed a geometric criterion to effectively study the stability switches of the model with only one discrete delay. Furthermore, the parameters of this model are relevant to the delay. The corresponding characteristic equation for the above model can be written as
where , are polynomials in . Recently, the prominent geometric method set forth by Beretta and Kuang [18] has been applied by a lot of scholars [16,19,20]. Subsequently, Beretta and Tang [21] extended the geometric stability switch criterion introduced in [18] to be appropriate for the more general characteristic equation
where , , and are polynomials in .
For the scenario that parameters of the model fail to be related to the delay, Gu et al. [22] put forward the crossing curves method for general systems with double delays. They studied the change of system stability for the following characteristic equation as delays vary.
where , are polynomials in . The application of this crossing curves method can be found in [23].
The geometric method developed by Gu et al. [22] was extended by An et al. [24] to explore the stability switches of a system with two discrete delays and delay-dependent parameters relevant only to one of the time delays. Clearly, the stability of such a model is absolutely decided by the roots of its characteristic equation
where , are polynomials in whose coefficients, say , are bounded functions of class . Some applications of this geometric method are available in [11,25,26].
Recently, based on the models in [27,28,29,30,31,32] and the hypothesis that the predator is generalist, that is, it can achieve an alternative food apart from the prey, Roy et al. [33] formulated the following predator–prey model with the generalist predator:
where and represent the densities of prey and generalist predator populations at time T, respectively. and are the growth rates of prey and predator populations, respectively. and denote the intra-specific competition among prey and predator populations, respectively. refers to the capture rate, a is the time of handling each captured prey, c represents the efficiency that predators convert consumed prey into new predators. In order to simplify the calculations, they rescaled model (5) by , , and then obtained the following model:
where , , and . All above parameters in model (6) are supposed to be positive. Moreover, inspired by the modeling method in [27,34,35], they modified model (6) by considering a stage structure for generalist predators. They classified the predator population into two stages: the immature stage and the mature stage. It is well known that immature predators require some time to mature. In the maturation process, there exists a natural death of immature predators. After accomplishing the maturation process, they deviate from the immature class and join the mature class. Thus, it can be seen that a maturation delay is common in a predator–prey model with a stage structure. That is to say, there is a close connection between the maturation delay and the model with stage structure. So the research of the model with a stage structure should focus on the maturation delay. Based on the above analysis and the hypothesis that immature predators are incapable of reproducing or capturing prey because they are weaker than mature predators, model (6) can be converted into
with and , , . In the above model, and represent the densities of immature and mature predator populations, respectively. denotes the death rate of immature predator populations. is the maturation delay and refers to the probability that a juvenile predator grows up to become an adult predator successfully. The biological significances of the remaining parameters are the same as those in model (6). For all we know, no one puts forward such a model apart from Roy et al. [33]. And it is more reasonable and more grounded in reality to investigate a model with a generalist predator and a maturation delay. Because and absolutely determined , they just considered the following delayed model for further investigation.
For the delayed model (8), they studied the positivity and boundedness of solutions, the stability of all equilibriums and local bifurcations. The conclusions they have reached are colorful. However, they demonstrated that under certain conditions, model (8) would not occur with a stability switch (see Proposition 6.1 in [33]), which means that Hopf bifurcations would not occur either. The main target of this paper is to extend the work carried out by Roy et al. [33] to further study the occurrence of stability switches and Hopf bifurcation near the positive equilibrium for model (8) by dint of the geometric stability switch criterion introduced in [18]. We hypothesize that incorporating a maturation delay in a predator–prey model with a stage structure for generalist predators and a Holling type-II functional response will influence the dynamic behavior of the system, potentially leading to stability switches and Hopf bifurcations. The subsequent sections will be devoted to exploring how the maturation delay affects the dynamics of model (8).
The organization of the remainder of the paper is as follows. In Section 2, we apply a geometric criterion to present the conditions for the occurrence of stability switches and Hopf bifurcations induced by the maturation delay. Section 3 focuses on the exploration of the nature of Hopf bifurcation by solving its normal form. To confirm our theoretical findings, numerical simulations are proceeded in Section 4. Finally, we summarize this paper with a conclusion section.
2. A Geometric Criterion for Hopf Bifurcation Values
In this section, we study the occurrence of possible stability switches and Hopf bifurcation values induced by the increase in time lag. According to Subsection 5.1 in [33], we can directly obtain the following details about the existence of positive equilibriums.
Lemma 1.
The following statements are correct for the model (8).
- (i)
- If , there exists at least one interior equilibrium for the delayed model (8).
- (ii)
- If , there is at least one positive equilibrium with the following sufficient condition holding:
It is direct from Subsection 5.2 in [33] that the characteristic equation of the model (8) at is
where
When , we can further receive the following lemma about the local stability of by the use of Routh–Hurwitz criteria.
Lemma 2.
When , if the condition : , holds, then the interior equilibrium is locally asymptotically stable.
In what follows, when , we plan to explore the stability switches and Hopf bifurcations of model (8) resulting from the maturation delay by applying the geometric criterion introduced in [18].
Equation (9) can be rewritten as
where and .
Obviously, and are analytic functions in and differentiable in . To utilize the geometric criterion, we also need to verify the following conclusions (see [18]):
- (i)
- for any ;
- (ii)
- If , , then for any ;
- (iii)
- for any ;
- (iv)
- for each has at most a finite number of real zeros;
- (v)
- Any positive root of is continuous and differentiable in whenever it exists.
Next, we will corroborate the above results in turn. In addition, it is posited that the hypothesis in Lemma 2 is true, that is to say, , .
- (i)
- for any ;
- (ii)
- for any on account of ;
- (iii)
- For any , ;
- (iv)
- Due toit is clearly true that admits at most a finite number of real zeros for each ;
- (v)
- This conclusion can be verified by the use of the implicit theorem directly.
Assume that are a pair of imaginary roots of Equation (11). Because and , in line with the symmetry, we just need to take into account . Plug into Equation (11) and then separate real and imaginary parts, we obtain
which gives
On the other side of the coin, if Equation (11) admits a root , then it follows that
Hence, we can rewrite (13) as
which means that
Consequently, if satisfies (13), then it must satisfy the following equation:
Define as the set of such that is a positive root of Equation (14). If I is empty, then there does not exist any stability switch and Hopf bifurcation value, which has been discussed and proven in [33]. However, they did not consider the case that I is nonempty any more. To fill up this gap, we extend the analysis in [33] by supposing that I is nonempty. In addition, we can sum up the following proposition.
Proposition 1.
If conditions and : hold, then Equation (14) has a unique positive root , , which means that I is nonempty.
Proof.
It is direct from (10) and that . Therefore, . Due to , we can further obtain with conditions and holding. The proof is thus completed. □
For any , the angle can be defined as the solution of
Hence, for any , we have , . Then, we can construct the maps , which is given by
where is a root of Equation (14). We further define the functions :
which are continuous and differentiable in . Then, we can obtain the following geometric criterion to certify the transversality condition and describe the direction of crossing the imaginary axis.
Theorem 1.
Assume that holds and that admits one positive root for some , then at , Equation (11) has a pair of simple conjugate pure imaginary roots , , which crosses the imaginary axis from left to right when and crosses the imaginary axis from right to left when , where is the transversality condition denoted by
Proof.
Compared to the proof of Theorem 2.2 in [18], we just need to further prove the transversality condition (18). From Theorem 2.2 in [18], we obtain that
It is direct from Equation (14) that
On the other hand, from (15), we can obtain , which implies that . Therefore, , which is equivalent to . The proof is completed. □
Clearly, for all , . Hence, if on I, then admits no zeros on I. To sum up, we show the following theorem with respect to the stability switches of and the occurrence of Hopf bifurcations.
Theorem 2.
- (i)
- If admits no positive real root , then when , is locally asymptotically stable for any and when , is locally asymptotically stable for .
- (ii)
- Assume that for some , admits the unique positive real root and satisfies . If , then is locally asymptotically stable for and unstable for . Moreover, the model (8) undergoes a Hopf bifurcation at when , . If , then is locally asymptotically stable for and unstable for .
- (iii)
- Assume that for some , has two positive real roots and satisfies , . Clearly, . If , then is locally asymptotically stable for and unstable for . In addition, a Hopf bifurcation occurs at when , . If , then is locally asymptotically stable for and unstable for .
- (iv)
- Assume that for some , has k positive real roots and that is the corresponding root of (12), , . Similar to the above process, we can probe into the stability switches and Hopf bifurcation of by judging the value of β and the sign of .
3. Direction of Hopf Bifurcation and Stability of Bifurcated Periodic Solutions
In this section, we intend to apply the center manifold theorem and normal form method put forward by Hassard et al. [36] to discuss the direction of Hopf bifurcation and the stability of bifurcated periodic solutions of model (8) at .
Let , . Therefore, the model (8) experiences a Hopf bifurcation when . Let , and replace , with , , respectively. Then model (8) can be converted to the functional differential equation in the phase space as
where , , and are given as follows.
where ,
with
By utilizing the Riesz representation theorem, it is quite easy to know that there is a matrix function filled with bounded variation (), such that
As a matter of fact, we can select
where represents the Dirac-delta function. For , we define
and
Then Equation (20) is equal to
For , the adjoint operator of can be denoted by
For , , a bilinear form is given by
where .
Through the argument in the previous section, we see that are the eigenvalues of . Hence, they are also the eigenvalues of . Moreover, we hold the assumption that and are the eigenvectors of and , which correspond to eigenvalues and , respectively.
A careful calculation gives
Direct from Equation (23), we obtain
As a result, we define
such that .
Furthermore, we can easily obtain due to .
In what follows, we intend to apply the same nations as those in [36]. We plan to work out the coordinates to describe the center manifold at . When , the solution of Equation (21) is denoted by . Define
On the center manifold , it follows that
where z and refer to local coordinates for in the direction of and . If is real, it is obvious to recognize that W is real. In consequence, we only consider real solutions.
For the solution of Equation (21) , because of , we obtain
The equation above can be rewritten as
with
Being aware that , and Equation (25), we have
Consequently, from Equation (26), we obtain
Comparing the above formula with Equation (26), we can obtain
We further require to derive the expression of and on account of their appearance in .
From Equations (21) and (24), we gain
where
Notice that , that is to say
Paying attention to the corresponding coefficients, we can obtain
From (27), we observe that for ,
Comparing the coefficients of the above equation with those in Equation (28), we have
It is direct from (29), (31) and the definition of A that
which indicates that
where , . In addition, they are all constant vectors. Next, we shall be devoted to finding appropriate and .
It is obvious that when , . So it follows that
Moreover, according to the expression of A and (29), we obtain
and
Now, substituting the first equation of (32) and (33) into (35) and observing that
and
then we obtain
which results in
that is
Thus, we can determine from (32), (37) and (38). Furthermore, we are able to calculate such values:
which describe the nature of bifurcated periodic solutions in the center manifold at the threshold value . Therewith, we can present the following theorem.
Theorem 3.
The following statements are all correct.
- (i)
- The direction of the Hopf bifurcation is determined by the sign of . If , the Hopf bifurcation is supercritical (subcritical).
- (ii)
- The stability of the bifurcated periodic solutions is decided by the sign of . If , the bifurcated periodic solutions are unstable (stable).
- (iii)
- The period of the bifurcated periodic solutions is decided by the sign of . The period increases (decreases) if .
4. Numerical Simulation
In this section, we provide a comprehensive analysis of how the maturation delay influences ecological dynamics of model (8) by giving some numerical examples. In addition, we will provide some biological interpretations.
In the first place, in accordance with the values of parameters in [33] and conditions given in the theoretical part of our paper, we choose
with initial conditions .
When , there exists one and only one positive equilibrium . Moreover, is locally asymptotically stable based on Lemma 2, just as shown in Figure 1. Biologically, when it takes no time for immature predators to mature, two populations can coexist in a stable state for a long time.

Figure 1.
Dynamic behavior of the model (8) with , , , and . In this case, the interior equilibrium is locally asymptotically stable. (a) Time series plot, (b) phase portrait.
We draw the image of the function versus with the parameters satisfying (40) by using Matlab, just as illustrated in Figure 2. Figure 2a manifests that has two zeros, one is , the other is , and , . Moreover, we find that for all from Figure 2b. Because for all , . Hence, admits no zeros on I.
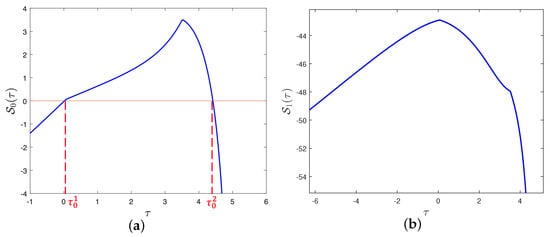
Figure 2.
Graphs of in terms of maturation delay with the values of parameters given in (40). (a) , it has two zeros, named and , (b) , it has no zeros on I.
It is direct from Figure 2 and Theorem 2 that is locally asymptotically stable for and unstable for . By considering the maturation delay as the bifurcation parameter, we further plot the bifurcation diagrams of the model (8) to demonstrate the switching process of , just as illustrated in Figure 3. From a biological perspective, if the value of the maturation delay belongs to the interval , both populations can survive and tend to a stable state. Once the maturation delay passes through the threshold value , the positive equilibrium loses it stability and a Hopf bifurcation occurs, which also means that both individuals can still exist but their densities fluctuate periodically with time. When the maturation delay exceeds the threshold value , the positive equilibrium regains its stability, that is to say, both populations can coexist for a long time. We choose some with different values for simulations which are displayed in Figure 4. When , is locally asymptotically stable, just as illustrated in Figure 4a,f. When , is unstable, just as illustrated in Figure 4b–e. These simulations indicate that stability switches occur as moves from 0.04 to 2 to 4.7.
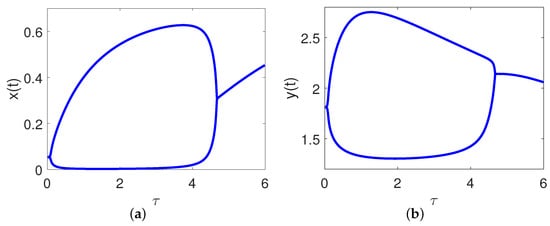
Figure 3.
Bifurcation diagrams of the model (8) by considering as the bifurcation parameter. (a) , (b) .
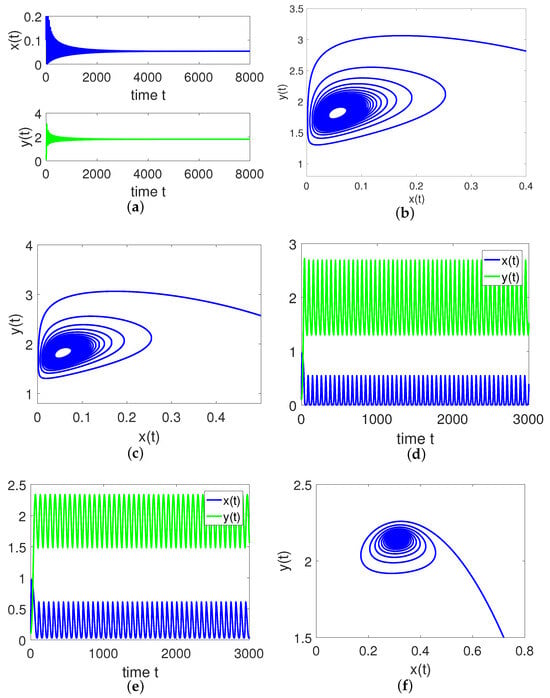
Figure 4.
Dynamics of the model (8) with different values of . (a) , (b) , (c) , (d) , (e) , (f) .
5. Conclusions
In this paper, an existing delayed predator–prey model with maturation delay, a stage structure for generalist predators and a Holling type-II functional response was investigated. We first obtained the conditions for the existence of the stability switches of the positive equilibrium and Hopf bifurcations caused by the increase in the value of the maturation delay for model (8) by utilizing the geometric criterion proposed by Beretta and Kuang [18]. After that, we discussed the properties of Hopf bifurcation by dint of the normal form method and center manifold theory. At last, we confirmed our theoretical findings by numerical simulations. Notice that, the authors in [29] considered a predator–prey model with a Holling-II type functional response and a prey refuge and then investigated the Hopf bifurcation of the model by seeing some key parameters as bifurcation parameters, such as the refuge parameter and death rate of the predator. Roy et al. [33] further introduced the maturation delay and generalist predator into the modeling framework and then proposed model (7). Based on model (7), we extended the research carried out in [33]. We further studied the stability-switching properties of the positive equilibrium and Hopf bifurcation by considering the maturation delay as a bifurcation parameter, which was not conducted in [33]. Moreover, the authors in [29,33] all applied the traditional theoretical method to study the bifurcation. Compared to the method in [29,33], we utilized a geometric method, which is novel and practical. The results we obtained in this article are very important in respect of ecology and are essential for enhancing the predictive power of the mathematical model and have significant implications for understanding and managing predator–prey interactions in real-world ecosystems. They indicate that the delay lengths affect the stability of the model. As a result, we can legitimately control the value of the maturation delay to make stable, which is beneficial to the coexistence of populations, beneficial to biodiversity protection.
Some interesting topics also deserve further considerations. For example, one can introduce additional biological delays in model (7), such as gestation delay, because the predator spends time in reproducing after consuming the prey and then try to study the stability switching properties of the positive equilibrium on the two-delay plane. If we consider other types of functional responses, what changes may take place? Moreover, a model incorporating stochastic elements to mimic random environmental fluctuations could be more realistic. Conducting a thorough sensitivity analysis of the key parameters could be conducive to identify which factors most significantly affect the stability of the model. Performing robustness checks would be helpful to ensure that the conclusions are not sensitive to specific starting points or configurations. Hence, considering a model with the above factors and further carrying out some analysis and checks are necessary. All of these are presented here for further research.
Author Contributions
Conceptualization, Z.-W.L. and X.-Y.M.; methodology, Z.-W.L.; software, Z.-W.L. and X.-Y.M.; validation, Z.-W.L. and X.-Y.M.; formal analysis, Z.-W.L.; investigation, Z.-W.L. and X.-Y.M.; writing—original draft, Z.-W.L.; writing—reviewing and editing, Z.-W.L. and X.-Y.M.; visualization, Z.-W.L. and X.-Y.M.; supervision, X.-Y.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant numbers 12161054 and 11861044); the Excellent Postgraduate Innovation Star of Gansu Province (grant number 2023CXZX-492); the Doctoral Foundation of Lanzhou University of Technology; and the HongLiu First-Class Disciplines Development Program of Lanzhou University of Technology.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We sincerely appreciate the Editors for their careful guidance and the reviewers for their valuable comments. We thank them for all their efforts in perfecting this paper.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Lotka, A.J. Elements of physical biology. Am. J. Public Health 1926, 21, 341–343. [Google Scholar]
- Volterra, V. Variations and fluctuations of the number of individuals in animal species living together. J. Du Conseil 1928, 3, 3–51. [Google Scholar] [CrossRef]
- Tang, G.Y.; Tang, S.Y.; Cheke, R.A. Global analysis of a Holling type II predator-prey model with a constant prey refuge. Nonlinear Dynam. 2013, 76, 635–647. [Google Scholar] [CrossRef]
- Khajanchi, S.; Banerjee, S. Role of constant prey refuge on stage structure predator-prey model with ratio dependent functional response. Appl. Math. Comput. 2017, 314, 193–198. [Google Scholar] [CrossRef]
- Zhang, H.S.; Cai, Y.L.; Fu, S.M.; Wang, W.M. Impact of the fear effect in a prey-predator model incorporating a prey refuge. Appl. Math. Comput. 2019, 356, 328–337. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, Z.L.; Li, Z. Modeling the Allee effect and fear effect in predator-prey system incorporating a prey refuge. Adv. Differ. Equ. 2020, 321, 321. [Google Scholar] [CrossRef]
- Bellman, R.E.; Cooke, K.L. Differential-Difference Equations; Academic Press: New York, NY, USA, 1963. [Google Scholar]
- Kuang, Y. Delay Differential Equations with Applications in Population Dynamics; Academic Press: Boston, MA, USA, 1993. [Google Scholar]
- Smith, H. An Introduction to Delay Differential Equations with Applications to the Life Sciences; Springer: New York, NY, USA, 2011. [Google Scholar]
- Kuang, Y.; Nagy, J.D.; Eikenberry, S.E. Introduction to Mathematical Oncology; Chemical Rubber Company Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Li, S.; Yuan, S.L.; Jin, Z.; Wang, H. Bifurcation analysis in a diffusive predator-prey model with spatial memory of prey, Allee effect and maturation delay of predator. J. Differ. Equ. 2023, 357, 32–63. [Google Scholar] [CrossRef]
- Cooke, K.L.; Grossman, Z. Discrete delay, distributed delay and stability switches. J. Math. Anal. Appl. 1982, 86, 592–627. [Google Scholar] [CrossRef]
- Song, Y.L.; Cao, X.; Zhang, T.H. Bistability and delay-induced stability switches in a cancer network with the regulation of microRNA. Commun. Nonlinear Sci. 2018, 54, 302–319. [Google Scholar] [CrossRef]
- Meng, X.Y.; Li, J. Dynamical behavior of a delayed prey-predator-scavenger system with fear effect and linear harvesting. Int. J. Biomath. 2021, 14, 2150024. [Google Scholar] [CrossRef]
- Li, S.; Huang, C.D.; Song, X.Y. Novel method to detect Hopf bifurcation in a delayed fractional-order network model with bidirectional ring structure. Int. J. Biomath. 2023, 16, 2250117. [Google Scholar] [CrossRef]
- Liang, Z.W.; Meng, X.Y. Stability and Hopf bifurcation of a multiple delayed predator–prey system with fear effect, prey refuge and Crowley—Martin function. Chaos Solitons Fractals 2023, 175, 113955. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Yang, Z. Asymptotic stability for quaternion-valued fuzzy BAM neural networks via integral inequality approach. Chaos Solitons Fractals 2023, 169, 113227. [Google Scholar] [CrossRef]
- Beretta, E.; Kuang, Y. Geometric stability switch criteria in delay differential systems with delay dependent parameters. SIAM J. Math. Anal. 2002, 33, 1144–1165. [Google Scholar] [CrossRef]
- Xu, X.F.; Wei, J.J. Bifurcation analysis of a spruce budworm model with diffusion and physiological structures. J. Differ. Equ. 2017, 262, 5206–5230. [Google Scholar] [CrossRef]
- Meng, X.Y.; Lu, M.M. Stability and bifurcation of a delayed prey-predator eco-epidemiological model with the impact of media. AIMS Math. 2023, 8, 17038–17066. [Google Scholar] [CrossRef]
- Beretta, E.; Tang, Y.B. Extension of a geometric stability switch criterion. Funkc. Ekvacioj 2003, 46, 337–361. [Google Scholar] [CrossRef][Green Version]
- Gu, K.Q.; Niculescu, S.I.; Chen, J. On stability crossing curves for general systems with two delays. J. Math. Anal. Appl. 2005, 311, 231–253. [Google Scholar] [CrossRef]
- Wang, C.H.; Yuan, S.L.; Wang, H. Spatiotemporal patterns of a diffusive prey-predator model with spatial memory and pregnancy period in an intimidatory environment. J. Math. Biol. 2022, 84, 12. [Google Scholar] [CrossRef]
- An, Q.; Beretta, E.; Kuang, Y.; Wang, C.C.; Wang, H. Geometric stability switch criteria in delay differential equations with two delays and delay dependent parameters. J. Differ. Equ. 2019, 266, 7073–7100. [Google Scholar] [CrossRef]
- Shu, H.Y.; Xu, W.X.; Wang, X.S.; Wu, J.H. Complex dynamics in a delay differential equation with two delays in tick growth with diapause. J. Differ. Equ. 2020, 269, 10937–10963. [Google Scholar] [CrossRef]
- Jiang, Z.C.; Zhao, Y.; Bai, X.L.; Zhang, Z.X. Bifurcation and control of a planktonic ecological system with double delays by delayed feedback control. J. Frankl. Inst. 2021, 358, 3609–3632. [Google Scholar] [CrossRef]
- Gourley, S.A.; Kuang, Y. A stage structured predator-prey model and its dependence on maturation delay and death rate. J. Math. Biol. 2004, 49, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Shi, J.P.; Wei, J.J. The effect of delay on a diffusive predator-prey system with Holling Type-II predator functional response. Commun. Pure Appl. Math. 2013, 12, 481–501. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, W.; Song, Y.F.; Zheng, Z.G.; Lu, J.H.; Chen, S.H. Hopf bifurcation analysis of a predator–prey model with Holling-II type functional response and a prey refuge. Nonlinear Dynam. 2019, 97, 1439–1450. [Google Scholar] [CrossRef]
- Bai, D.; Yu, J.; Fan, M.; Kang, Y. Dynamics for a non-autonomous predator-prey system with generalist predator. J. Math. Anal. Appl. 2020, 485, 123820. [Google Scholar] [CrossRef]
- Dey, S.; Banerjee, M.; Ghorai, S. Analytical detection of stationary turing pattern in a predator-prey system with generalist predator. Math. Model. Nat. Phenom. 2022, 17, 33. [Google Scholar] [CrossRef]
- Roy, J.; Banerjee, M. Global stability of a predator–prey model with generalist predator. Appl. Math. Lett. 2023, 142, 108659. [Google Scholar] [CrossRef]
- Roy, J.; Dey, S.; Banerjee, M. Maturation delay induced stability enhancement and shift of bifurcation thresholds in a predator–prey model with generalist predator. Math. Comput. Simul. 2023, 211, 368–393. [Google Scholar] [CrossRef]
- Liu, S.; Beretta, E. A stage-structured predator-prey model of Beddington-DeAngelis type. SIAM J. Appl. Math. 2006, 66, 1101–1129. [Google Scholar] [CrossRef]
- Banerjee, M.; Takeuchi, Y. Maturation delay for the predators can enhance stable coexistence for a class of prey-predator models. J. Theor. Biol. 2017, 412, 154–171. [Google Scholar] [CrossRef]
- Hassard, B.D.; Kazarinoff, N.D.; Wan, Y.H. Theory and Applications of Hopf Bifurcation; Cambridge University Press: Cambridge, UK, 1981. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).