
The “Water Problem”(sic), the Illusory Pond and Life’s Submarine Emergence—A Review

Abstract
Central to understanding “living mater” is appreciating its sheer improbability. [1].
It is through functional properties, not structure, that the organization of a purposive system is expressed. [2].
1. Introduction
2. Evaporating Pond Theory of Life’s “Origin”
3. Dirty Chemistry
4. RNA-DNAology
5. AVT Critiques: The False Requiem
6. Wächtershäuser’s Probe
7. The “Pond” in the Hellish Hadean
8. The Retreat to Mars!
9. Experimental Results Pertinent to the AVT
10. How Might the Nucleotide Penthouse be Accessed from the Submarine Alkaline Vent
11. The “Origin of Life” Community
12. What’s Next for the AVT?
Acknowledgments
Conflicts of Interest
References
- Wicken, J.S. Evolution, Thermodynamics and Information: Extending the Darwinian Program; Oxford University Press: New York, NY, USA, 1987. [Google Scholar]
- Cottrell, A. The natural philosophy of engines. Contemp. Phys. 1979, 20, 1–10. [Google Scholar] [CrossRef]
- Marshall, M. How the first life on Earth survived its biggest threat—water. Nature 2020, 588, 210–213. [Google Scholar] [CrossRef]
- Sutherland, J.D. Studies on the origin of life—The end of the beginning. Nat. Rev. Chem. 2017, 1, 1–7. [Google Scholar] [CrossRef]
- Branscomb, E.; Russell, M.J. Frankenstein or a submarine alkaline vent: Who is responsible for abiogenesis? Part 2: As life is now, so it must have been in the beginning? BioEssays 2018, 40, 1700182. [Google Scholar] [CrossRef]
- Milshteyn, D.; Damer, B.; Havig, J.R.; Deamer, D. Amphiphilic Compounds Assemble into Membranous Vesicles in Hydrothermal Hot Spring Water but Not in Seawater. Life 2018, 8, 11. [Google Scholar] [CrossRef]
- Rajamani, S.; Vlassov, A.; Benner, S.; Coombs, A.; Olasagasti, F.; Deamer, D. Lipid-assisted Synthesis of RNA-like Polymers from Mononucleotides. Orig. Life Evol. Biosph. 2008, 38, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D.; Damer, B.; Kompanichenko, V. Hydrothermal Chemistry and the Origin of Cellular Life. Astrobiology 2019, 19, 1523–1537. [Google Scholar] [CrossRef]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef]
- Frenkel-Pinter, M.; Haynes, J.W.; Martin, C.; Petrov, A.S.; Burcar, B.T.; Krishnamurthy, R.; Hud, N.V.; Leman, L.J.; Williams, L.D. Selective incorporation of proteinaceous over nonproteinaceous cationic amino acids in model prebiotic oligomerization reactions. Proc. Natl Acad. Sci. USA 2019, 116, 16338–16346. [Google Scholar] [CrossRef]
- Bonfio, C.; Russell, D.A.; Green, N.J.; Mariani, A.; Sutherland, J.D. Activation chemistry drives the emergence of functionalised protocells. Chem. Sci. 2020, 11, 10688–10697. [Google Scholar] [CrossRef]
- Schwartz, A.W.; Van der Veen, M.; Bisseling, T.; Chittenden, G.J.F. Prebiotic phosphorylation. II-nucleotide synthesis in the reaction system apatite-cyanogen-water. BioSystems 1973, 5, 119–122. [Google Scholar] [CrossRef]
- Lahav, N.; Chang, S. The possible role of solid surface area in condensation reactions during chemical evolution: Re-evaluation. J. Mol. Evol. 1976, 8, 357–380. [Google Scholar] [CrossRef]
- Lahav, N.; White, D.; Chang, S. Peptide formation in the prebiotic era: Thermal condensation of glycine in fluctuating clay environments. Science 1978, 201, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Darwin, F. (Ed.) The Life and Letters of Charles Darwin; John Murray: London, UK, 1888; Volume 3. [Google Scholar]
- Powner, M.W.; Gerland, B.; Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef]
- Mulkidjanian, A.; Bychkov, A.; Dibrova, D.; Galperin, M.; Koonin, E. Origin of first cells at terrestrial, anoxic geothermal fields. Proc. Natl. Acad. Sci. USA 2012, 109, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A.; Kim, H.J.; Carrigan, M.A. Asphalt, water, and the prebiotic synthesis of ribose, ribonucleosides, and RNA. Acc. Chem. Res. 2012, 45, 2025–2034. [Google Scholar] [CrossRef]
- Hud, N.V. Searching for lost nucleotides of the pre-RNA World with a self-refining model of early Earth. Nat. Commun. 2018, 9, 1–4. [Google Scholar]
- Ross, D.S.; Deamer, D. Dry/Wet Cycling and the Thermodynamics and Kinetics of Prebiotic Polymer Synthesis. Life 2016, 6, 28. [Google Scholar] [CrossRef]
- Damer, B.; Deamer, D. Coupled phases and combinatorial selection in fluctuating hydrothermal pools: A scenario to guide experimental approaches to the origin of cellular life. Life 2015, 5, 872–887. [Google Scholar] [CrossRef]
- Pearce, B.K.D.; Pudritz, R.E.; Semenov, D.A.; Henning, T.K. Origin of the RNA world: The fate of nucleobases in warm little ponds. Proc. Natl. Acad. Sci. USA 2017, 114, 11327–11332. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Schneider, C.; Okamura, H.; Crisp, A.; Amatov, T.; Dejmek, M.; Carell, T. Wet-dry cycles enable the parallel origin of canonical and non-canonical nucleosides by continuous synthesis. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Becker, S.; Feldmann, J.; Wiedemann, S.; Okamura, H.; Schneider, C.; Iwan, K.; Crisp, A.; Rossa, M.; Amatov, T.; Carell, T. Unified prebiotically plausible synthesis of pyrimidine and purine RNA ribonucleotides. Science 2019, 366, 76–82. [Google Scholar] [CrossRef]
- Hargrave, M.; Spencer, S.K.; Deamer, D.W. Computational models of polymer synthesis driven by dehydration/ rehydration cycles: Repurination in simulated hydrothermal fields. J. Mol. Evol. 2018, 86, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, W.R.; Mulvihill, S.J.; Deamer, D.W. Synthesis of phospholipids and membranes in prebiotic conditions. Nature 1977, 266, 78–80. [Google Scholar] [CrossRef]
- Nainytė, M.; Müller, F.; Ganazzoli, G.; Chan, C.Y.; Crisp, A.; Globisch, D.; Carell, T. Amino Acid Modified RNA Bases as Building Blocks of an Early Earth RNA-Peptide World. Chemistry 2020, 26, 14856. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A.; Bell, E.A.; Biondi, E.; Brasser, R.; Carell, T.; Kim, H.J.; Mojzsis, S.J.; Omran, A.; Pasek, M.A.; Trail, D. When did life likely emerge on Earth in an RNA-first process? arXiv 2019, arXiv:1908.11327. [Google Scholar]
- Damer, B.F. A field trip to the Archaean in search of Darwin’s warm little pond. Life 2016, 6, 21. [Google Scholar] [CrossRef]
- Clark, B.C.; Kolb, V.M. Comet Pond II: Synergistic Intersection of Concentrated Extraterrestrial Materials and Planetary Environments to Form Procreative Darwinian Ponds. Life 2018, 8, 12. [Google Scholar] [CrossRef]
- Van Kranendonk, M.J.; Baumgartner, R.; Djokic, T.; Ota, T.; Steller, L.; Garbe, U.; Nakamura, E. Elements for the Origin of Life on Land: A Deep-Time Perspective from the Pilbara Craton of Western Australia. Astrobiology 2021, 21, 39–59. [Google Scholar] [CrossRef]
- Van Kranendonk, M.J.; Deamer, D.W.; Djokic, T. Life springs: Darwin’s warm little pond revisited. Sci. Am. 2017, 317, 28–35. [Google Scholar] [CrossRef]
- Darwin, C. The Origin of Species; A Mentor Book: New York, NY, USA; Scarborough, ON, Canada, 1872. [Google Scholar]
- Yockey, H.P. Comments on “Let there be life”; Thermodynamic reflections on biogenesis and evolution” by Avshalom C. Elitzur. J. Theor. Biol. 1995, 176, 349–355. [Google Scholar] [CrossRef]
- Hud, N.; Brian, J.; Cafferty, B.J.; Krishnamurthy, R.; Williams, R.D. The Origin of RNA and ‘‘My Grandfather’s Axe’’. Chem. Biol. 2013, 20, 466–474. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J.; Turner, D. In vitro growth of iron sulphide chimneys: Possible culture chambers for origin-of-life experiments. Terra Nova 1989, 1, 238–241. [Google Scholar] [CrossRef]
- Russell, M.J. Plate tectonics and hydrothermal ore deposits. In Understanding the Earth, 2nd ed.; Brown, C.G., Hawkesworth, C.J., Wilson, R.C.L., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 204–221. [Google Scholar]
- Russell, M.J.; Hall, A.J.; Fallick, A.E.; Boyce, A.J. On hydrothermal convection systems and the emergence of life. Econ. Geol. 2005, 100, 419–438. [Google Scholar]
- Nitschke, W.; Russell, M.J. Hydrothermal focusing of chemical and chemiosmotic energy, supported by delivery of catalytic Fe, Ni, Mo/W, Co, S and Se, forced life to emerge. J. Mol. Evol. 2009, 69, 481–496. [Google Scholar] [CrossRef]
- Shapiro, R. Origins: A Skeptic’s Guide to the Creation of Life on Earth; Summit Books: New York, NY, USA, 1986. [Google Scholar]
- Lazcano, A.; Miller, S.L. On the origin of metabolic pathways. J. Mol. Evol. 1999, 49, 424–431. [Google Scholar] [CrossRef]
- Dass, A.V.; Hickman-Lewis, K.; Brack, A.; Kee, T.P.; Westall, F. Stochastic prebiotic chemistry within realistic geological systems. ChemistrySelect 2016, 1, 4906–4926. [Google Scholar] [CrossRef]
- Guttenberg, N.; Virgo, N.; Chandru, K.; Scharf, C.; Mamajanov, I. Bulk measurements of messy chemistries are needed for a theory of the origins of life. Philos. Trans. R. Soc. A 2017, 375, 20160347. [Google Scholar] [CrossRef]
- Walker, S.I.; Bains, W.; Cronin, L.; DasSarma, S.; Danielache, S.; Domagal-Goldman, S.; Kacar, B.; Kiang, N.Y.; Lenardic, A.; Reinhard, C.T.; et al. Exoplanet biosignatures: Future directions. Astrobiology 2018, 18, 779–824. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, E. What is Life? The Physical Aspect of the Living Cell; Based on Lectures Delivered under the Auspices of the Institute at Trinity College, Dublin, in February; University Press: Hong Kong, China, 1945. [Google Scholar]
- Prigogine, I. Order through fluctuations: Self-organization and social systems. In Evolution and Consciousness: Human Systems in Transition; Jantsch, E., Ed.; Waddington Reading Addison-Wesley: Reading, MA, USA, 1976; pp. 93–130. [Google Scholar]
- Branscomb, E.; Russell, M.J. Frankenstein or a submarine alkaline vent: Who is responsible for abiogenesis? Part 1: What is life—that it might create itself? BioEssays 2018, 40, e1700179. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, W. Discussions on Philosophy and Literature; Harper & Brothers Publishers: New York, NY, USA, 1855. [Google Scholar]
- Deamer, D.W.; Barchfeld, G.L. Encapsulation of macromolecules by lipid vesicles under simulated prebiotic conditions. J. Mol. Evol. 1982, 18, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Monnard, P.A.; Apel, C.L.; Kanavarioti, A.; Deamer, D.W. Influence of ionic inorganic solutes on self-assembly and polymerization processes related to early forms of life: Implications for a prebiotic aqueous medium. Astrobiology 2002, 2, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D.; Singaram, S.; Rajamani, S.; Kompanichenko, V.; Guggenheim, S. Self-assembly processes in the prebiotic environment. Phil. Trans. R. Soc. B 2006, 361, 1809–1818. [Google Scholar] [CrossRef] [PubMed]
- Mulkidjanian, A.Y. On the origin of life in the zinc world: Photosynthesizing, porous edifices built of hydrothermally precipitated zinc sulfide as cradles of life on Earth. Biol. Dir. 2009, 4, 1–39. [Google Scholar] [CrossRef]
- Mulkidjanian, A.Y.; Cherepanov, D.A.; Galperin, M.Y. Survival of the fittest before the beginning of life: Selection of the first oligonucleotide-like polymers by UV light. BMC Evol. Biol. 2003, 3, 12. [Google Scholar] [CrossRef]
- Deamer, D.; Weber, A.L. Bioenergetics and life’s origins. Cold Spring Harbor Perspect. Biol. 2010, 2, a004929. [Google Scholar]
- Hazen, R.M.; Sverjensky, D.A. Mineral surfaces, geochemical complexities, and the origins of life. Cold Spring Harbor Perspect. Biol. 2010, 2, a002162. [Google Scholar] [CrossRef]
- Kim, H.J.; Ricardo, A.; Illangkoon, H.I.; Kim, M.J.; Carrigan, M.A.; Frye, F.; Benner, S.A. Synthesis of carbohydrates in mineral-guided prebiotic cycles. J. Am. Chem. Soc. 2011, 133, 9457–9468. [Google Scholar] [CrossRef]
- De Guzman, V.; Shenasa, H.; Vercoutere, W.; Deamer, D. Generation of oligonucleotides under hydrothermal conditions by non-enzymatic polymerization. J. Mol. Evol. 2014, 78, 251–262. [Google Scholar] [CrossRef]
- Forsythe, J.G.; Yu, S.S.; Mamajanov, I.; Grover, M.A.; Krishnamurthy, R.; Fernández, F.M.; Hud, N.V. Ester-mediated amide bond formation driven by wet–dry cycles: A possible path to polypeptides on the prebiotic Earth. Angew. Chem. Int. Ed. Engl. 2015, 54, 9871–9875. [Google Scholar] [CrossRef]
- Hazen, R.M. Chance, necessity and the origins of life: A physical sciences perspective. Philos. Trans. R. Soc. A 2017, 375, 20160353. [Google Scholar] [CrossRef]
- Ritson, D.; Sutherland, J.D. Prebiotic synthesis of simple sugars by photoredox systems chemistry. Nat. Chem. 2012, 4, 895–899. [Google Scholar] [CrossRef]
- Shock, E.L. Chemical environments of submarine hydrothermal systems. Orig. Life Evol. Biosph. 1992, 22, 67–107. [Google Scholar] [CrossRef]
- Wood, B.J.; Walter, M.J.; Wade, J. Accretion of the Earth and segregation of its core. Nature 2006, 441, 825–833. [Google Scholar] [CrossRef]
- Russell, M.J.; Ponce, A. Six ‘Must-Have’ Minerals for Life’s Emergence: Olivine, Pyrrhotite, Bridgmanite, Serpentine, Fougerite and Mackinawite. Life 2020, 10, 291. [Google Scholar] [CrossRef]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; Duffy, C.D.; Sutherland, J.D. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar]
- Xu, J.; Chmela, V.; Green, N.J.; Russell, D.A.; Janicki, M.J.; Góra, R.W.; Szabla, R.; Bond, A.D.; Sutherland, J.D. Selective prebiotic formation of RNA pyrimidine and DNA purine nucleosides. Nature 2020, 582, 60–66. [Google Scholar] [CrossRef]
- Bonfio, C.; Godino, E.; Corsini, M.; de Biani, F.F.; Guella, G.; Mansy, S.S. Prebiotic iron–sulfur peptide catalysts generate a pH gradient across model membranes of late protocells. Nat. Catal. 2018, 1, 616–623. [Google Scholar] [CrossRef]
- Bonfio, C.; Valer, L.; Scintilla, S.; Shah, S.; Evans, D.J.; Jin, L.; Szostak, J.W.; Sasselov, D.D.; Sutherland, J.D.; Mansy, S.S. UV-light-driven prebiotic synthesis of iron–sulfur clusters. Nat. Chem. 2017, 9, 1229. [Google Scholar]
- Mann, A. Inner Workings: Making headway with the mysteries of life’s origins. Proc. Natl. Acad. Sci. USA 2021, 118, e2105383118. [Google Scholar]
- Deamer, D. Conjecture and hypothesis: The importance of reality checks. Beilst. J. Org. Chem. 2017, 13, 620–624. [Google Scholar] [CrossRef]
- Popper, K. Conjectures and Refutations. The Growth of Scientific Knowledge; Routledge: London, UK, 1963. [Google Scholar]
- Branscomb, E.; Biancalani, T.; Goldenfeld, N.; Russell, M. Escapement mechanisms and the conversion of disequilibria; the engines of creation. Phys. Rep. 2017, 677, 1–60. [Google Scholar] [CrossRef]
- Russell, M.J. Green Rust: The Simple Organizing ‘Seed’ of All Life? Life 2018, 8, 35. [Google Scholar] [CrossRef]
- Burcar, B.; Pasek, M.; Gull, M.; Cafferty, B.J.; Velasco, F.; Hud, N.V.; Menor-Salván, C. Darwin’s warm little pond: A one-pot reaction for prebiotic phosphorylation and the mobilization of phosphate from minerals in a urea-based solvent. Angew. Chem. Int. Ed. Engl. 2016, 55, 13249–13253. [Google Scholar] [CrossRef]
- Deamer, D. Where Did Life Begin? Testing Ideas in Prebiotic Analogue Conditions. Life 2021, 11, 134. [Google Scholar] [CrossRef]
- Bahcall, J.N.; Pinsonneault, M.H.; Basu, S. Solar models: Current epoch and time dependences, neutrinos, and helioseismo-logical properties. Astrophys. J. 2001, 555, 990. [Google Scholar] [CrossRef]
- Pipin, V.V.; Kosovichev, A.G. Effects of large-scale non-axisymmetric perturbations in the mean-field solar dynamo. Astrophys. J. 2015, 813, 134. [Google Scholar] [CrossRef]
- Gudel, M. The sun in time: Activity and environment. Living Rev. Sol. Phys. 2007, 4, 137. [Google Scholar] [CrossRef]
- Birch, F. Energetics of core formation. J. Geophys. Res. 1965, 69, 4377–4388. [Google Scholar] [CrossRef]
- Longuet-Higgins, M.S. The eigenfunctions of Laplace’s tidal equation over a sphere. Philos. Trans. R. Soc. Lond. A 1968, 262, 511–607. [Google Scholar] [CrossRef]
- Binder, A.B. The Moon: Its figure and orbital evolution. Geophys. Res. Lett. 1982, 9, 33–36. [Google Scholar] [CrossRef]
- Dones, L.; Tremaine, S. Why Does the Earth Spin Forward? Science 1993, 259, 350–354. [Google Scholar] [CrossRef]
- Zharkov, V.N. On the history of the lunar orbit. Astron. Vesn. 2000, 34, 1–11. [Google Scholar]
- Denis, C.; Rybicki, K.R.; Schreider, A.A.; Tomecka-Suchoń, S.; Varga, P. Length of the day and evolution of the Earth’s core in the geological past. Astron. Nachr. 2011, 332, 24–35. [Google Scholar] [CrossRef]
- Denis, C.; Schreider, A.A.; Varga, P.; Zavoti, J. Despinning of the Earth rotation in the geological past and geomagnetic paleointensities. J. Geodyn. 2002, 34, 667–685. [Google Scholar] [CrossRef]
- Sossi, P.A.; Burnham, A.D.; Badro, J.; Lanzirotti, A.; Newville, M.; O’Neill, H.S.C. Redox state of Earth’s magma ocean and its Venus-like early atmosphere. Sci. Adv. 2020, 6, eabd1387. [Google Scholar] [CrossRef] [PubMed]
- Dandonneau, Y.; Vega, A.; Loisel, H.; Du Penhoat, Y.; Menkes, C. Oceanic Rossby waves acting as a “hay rake” for ecosystem floating by-products. Science 2003, 302, 1548–1551. [Google Scholar] [CrossRef]
- Glukhovskii, M.Z.; Kuz’min, M.I. Extraterrestrial factors and their role in the Earth’s tectonic evolution in the early Precambrian. Russ. Geol. Geophys. 2015, 56, 959–977. [Google Scholar] [CrossRef]
- Marakushev, A.A.; Zinov’eva, N.G.; Paneyakh, N.A.; Marakushev, S.A. The origin and evolution of the solar system. Prostran I Vremya 2013, 2, 132–141. [Google Scholar]
- Malcuit, R.J. A Retrograde Gravitational Capture Model for the Earth-Moon System. In The Twin Sister Planets Venus and Earth; Springer: Cham, Switzerland, 2015; pp. 271–294. [Google Scholar]
- Bozóki, T.; Herein, M.; Galsa, A. Numerical evolution of the asymmetry in the compositionally inhomogeneous lower mantle driven by Earth’s rotation. Acta Geodaet. Geophys. 2017, 52, 331–343. [Google Scholar] [CrossRef]
- Lingam, M.; Loeb, A. Implications of tides for life on exoplanets. Astrobiology 2018, 18, 967–982. [Google Scholar] [CrossRef] [PubMed]
- Glikson, A.Y. Early Precambrian asteroid impact-triggered tsunami: Excavated seabed, debris flows, exotic boulders, and turbulence features associated with 3.47–2.47 Ga-old asteroid impact fallout units, Pilbara Craton, Western Australia. Astrobiology 2004, 4, 19–50. [Google Scholar] [CrossRef]
- Abramov, O.; Mojzsis, S.J. Microbial habitability of the Hadean Earth during the Late Heavy Bombardment. Nature 2009, 459, 419–422. [Google Scholar] [CrossRef]
- Sleep, N.H. Maintenance of permeable habitable subsurface environments by earthquakes and tidal stresses. Int. J. Astrobiol. 2012, 11, 257–268. [Google Scholar] [CrossRef]
- Abramov, O.; Kring, D.A.; Mojzsis, S.J. The impact environment of the hadean earth. Geochemistry 2013, 73, 227–248. [Google Scholar] [CrossRef]
- Boehnke, P.; Harrison, T.M. Illusory late heavy bombardments. Proc. Natl. Acad. Sci. USA 2016, 113, 10802–10806. [Google Scholar] [CrossRef] [PubMed]
- Zellner, N.E. Cataclysm no more: New views on the timing and delivery of lunar impactors. Orig. Life Evol. Biosph. 2017, 47, 261–280. [Google Scholar] [CrossRef] [PubMed]
- Monteux, J.; Andrault, D.; Guitreau, M.; Samuel, H.; Demouchy, S. A mushy Earth’s mantle for more than 500 Myr after the magma ocean solidification. Geophys. J. Int. 2020, 221, 1165–1181. [Google Scholar] [CrossRef]
- Solomatov, V.S. Magma Oceans and Primordial Mantle Differentiation. Treat. Geophys. 2007, 9, 91–120. [Google Scholar]
- Walter, M.J.; Nakamura, E.; Trønnes, R.G.; Frost, D.J. Experimental constraints on crystallization differentiation in a deep magma ocean. Geochim. Cosmochim. Acta 2005, 68, 4267–4284. [Google Scholar] [CrossRef]
- Sleep, N.H.; Zahnle, K.J.; Lupu, R.E. Terrestrial after-math of the Moon-forming impact. Philos. Trans. R. Soc. A 2014, 372, 20130172. [Google Scholar] [CrossRef]
- Kamber, B.S. The evolving nature of terrestrial crust from the Hadean, through the Archaean, into the Proterozoic. Precam. Res. 2015, 258, 48–82. [Google Scholar] [CrossRef]
- Morgan, J.P.; Morgan, W.J. Two-stage melting and the goldschmgeochemical evolution of the mantle: A recipe for mantle plum-pudding. Earth Planet. Sci. Lett. 1999, 170, 215–239. [Google Scholar] [CrossRef]
- Şengör, A.M.C.; Ernst, R.E.; Buchan, K.L. Elevation as indicator of mantle-plume activity. In Mantle plumes: Their identification through time. Geol. Soc. Am. 2001, 352, 183–225. [Google Scholar]
- Mann, P.; Taira, A. Global tectonic significance of the Solomon Islands and Ontong Java Plateau convergent zone. Tectonophysics 2004, 389, 137–190. [Google Scholar] [CrossRef]
- Russell, M.J.; Arndt, N.T. Geodynamic and metabolic cycles in the Hadean. Biogeosciences 2005, 2, 97–111. [Google Scholar] [CrossRef]
- Bédard, J.H. A catalytic delamination-driven model for coupled genesis of Archaean crust and sub-continental lithospheric mantle. Geochim. Cosmochim. Acta 2006, 70, 1188–1214. [Google Scholar] [CrossRef]
- Morbidelli, A.; Chambers, J.; Lunine, J.I.; Petit, J.M.; Robert, F.; Valsecchi, G.B.; Cyr, K.E. Source regions and timescales for the delivery of water to the Earth. Meteorit. Planet. Sci. 2000, 35, 1309–1320. [Google Scholar] [CrossRef]
- Bounama, C.; Franck, S.; von Bloh, W. The fate of the Earth’s ocean. Hydrol. Earth Syst. Sci. 2001, 5, 569–575. [Google Scholar] [CrossRef]
- Valley, J.W.; Lackey, J.S.; Cavosie, A.J.; Clechenko, C.C.; Spicuzza, M.J.; Basei, M.A.S.; Bindeman, I.N.; Ferreira, V.P.; Sial, A.N.; King, E.M.; et al. 4.4 billion years of crustal maturation: Oxygen isotope ratios of magmatic zircon. Contr. Miner. Petrol. 2005, 150, 561–580. [Google Scholar] [CrossRef]
- Cavosie, A.J.; Valley, J.W.; Wilde, S.A. The oldest terrestrial mineral record: A review of 4400 to 3900 ma detrital zircons from Jack Hills, Western Australia. Dev. Precamb. Geol. 2007, 15, 91–111. [Google Scholar]
- Pope, E.C.; Bird, D.K.; Rosing, M.T. Isotope composition and volume of Earth’s early oceans. Proc. Natl. Acad. Sci. USA 2012, 109, 4371–4376. [Google Scholar] [CrossRef]
- O’Neil, J.; Boyet, M.; Carlson, R.W.; Paquette, J.-L. Half a billion years of reworking of Hadean mafic crust to produce the Nuvvuagittuq Eoarchean felsic crust. Earth Planet. Sci. Lett. 2013, 379, 13–25. [Google Scholar] [CrossRef]
- Korenaga, J.; Planavsky, N.J.; Evans, D.A. Global water cycle and the coevolution of the Earth’s interior and surface environment. Phil. Trans. R. Soc. A 2017, 375, 20150393. [Google Scholar] [CrossRef]
- Genda, H. Origin of Earth’s oceans: An assessment of the total amount, history and supply of water. Geochem. J. 2016, 50, 27–42. [Google Scholar] [CrossRef]
- Ueda, H.; Shibuya, T. Composition of the Primordial Ocean Just after Its Formation: Constraints from the Reactions between the Primitive Crust and a Strongly Acidic, CO2-Rich Fluid at Elevated Temperatures and Pressures. Minerals 2021, 11, 389. [Google Scholar] [CrossRef]
- Johnson, B.W.; Wing, B.A. Limited Archaean continental emergence reflected in an early Archaean 18O-enriched ocean. Nat. Geosci. 2020, 13, 243–248. [Google Scholar] [CrossRef]
- Deng, J.; Du, Z.; Karki, B.B.; Ghosh, D.B.; Lee, K.K. A magma ocean origin to divergent redox evolutions of rocky planetary bodies and early atmospheres. Nat. Commun. 2020, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Trail, D.; Watson, E.B.; Tailby, N.D. The oxidation state of Hadean magmas and implications for early Earth’s atmosphere. Nature 2011, 480, 79–82. [Google Scholar] [CrossRef]
- Galimov, E.M. Redox evolution of the Earth caused by a multi-stage formation of its core. Earth Planet. Sci. Lett. 2005, 233, 263–276. [Google Scholar] [CrossRef]
- Lammer, H.; Stökl, A.; Erkaev, N.V.; Dorfi, E.A.; Odert, P.; Güdel, M.; Kulikov, Y.N.; Kislyakova, K.G.; Leitzinger, M. Origin and loss of nebula-captured hydrogen envelopes from ‘sub’-to ‘super-Earths’ in the habitable zone of Sun-like stars. Mon. Not. R. Astron. Soc. 2014, 439, 3225–3238. [Google Scholar] [CrossRef]
- Massol, H.; Hamano, K.; Tian, F.; Ikoma, M.; Abe, Y.; Chassefière, E.; Davaille, A.; Genda, H.; Güdel, M.; Hori, Y.; et al. Formation and evolution of protoatmospheres. Space Sci. Rev. 2016, 205, 153–211. [Google Scholar] [CrossRef]
- Zahnle, K.; Arndt, N.; Cockell, C.; Halliday, A.; Nisbet, E.; Selsis, F.; Sleep, N.H. Emergence of a habitable planet. Space Sci. Rev. 2007, 129, 35–78. [Google Scholar] [CrossRef]
- Yung, Y.L.; McElroy, M.B. Fixation of nitrogen in the prebiotic atmosphere. Science 1979, 203, 1002–1004. [Google Scholar] [CrossRef] [PubMed]
- Kasting, J.F.; Ackerman, T.P. Climatic consequences of very high carbon dioxide levels in the Earth’s early atmosphere. Science 1986, 234, 1383–1385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Helsdon, J.J.H.; Farley, R.D. Numerical modeling of lightning produced NOx using an explicit lightning scheme: Two-dimensional simulation as a ‘proof of concept’. J. Geophys. Res. 2003, 108, ACH 5-1–ACH 5-20. [Google Scholar]
- Dasgupta, R.; Hirschmann, M.M. Melting in the Earth’s deep upper mantle caused by carbon dioxide. Nature 2006, 440, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Hirschmann, M.M.; Tenner, T.; Aubaud, C.; Withers, A.C. Dehydration melting of nominally anhydrous mantle: The primacy of partitioning. Phys. Earth Planet. Inter. 2009, 176, 54–68. [Google Scholar] [CrossRef]
- Hirschmann, M.M. Comparative deep Earth volatile cycles: The case for C recycling from exosphere/mantle fractionation of major (H2O, C, N) volatiles and from H2O/Ce, CO2/Ba, and CO2/Nb exosphere ratios. Earth Planet. Sci. Lett. 2018, 502, 262–273. [Google Scholar] [CrossRef]
- Martin, R.S.; Mather, T.A.; Pyle, D.M. Volcanic emissions and the early earth atmosphere. Geochim. Cosmochim. Acta 2007, 71, 3673–3685. [Google Scholar] [CrossRef]
- Wong, M.L.; Charnay, B.D.; Gao, P.; Yung, Y.L.; Russell, M.J. Nitrogen Oxides in Early Earth’s Atmosphere as Electron Acceptors for Life’s Emergence. Astrobiology 2017, 17, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, S.; Grenfell, J.L.; Lammer, H.; de Vera, J.P.; Sproß, L.; Airapetian, V.S.; Sinnhuber, M.; Rauer, H. Atmospheric nitrogen when life evolved on Earth. Astrobiology 2020, 20, 1413–1426. [Google Scholar] [CrossRef] [PubMed]
- Catling, D.C.; Zahnle, K.J. The archean atmosphere. Sci. Adv. 2020, 6, eaax1420. [Google Scholar] [CrossRef]
- Datz, S.; Smith, W.T., Jr.; Taylor, E.H. Molecular association in alkali halide vapors. J. Chem. Phys. 1961, 34, 558–564. [Google Scholar] [CrossRef]
- Van Groos, A.K.; Wyllie, P.J. Melting relationships in the system NaAlSi3O8-NaCl-H2O at one kilobar pressure, with petrological applications. J. Geol. 1969, 77, 581–605. [Google Scholar] [CrossRef]
- Macleod, G.; McKeown, C.; Hall, A.J.; Russell, M.J. Hydrothermal and oceanic pH conditions of possible relevance to the origin of life. Orig. Life Evol. Biosph. 1994, 24, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Kusakabe, M.; Tanyileke, G.Z.; McCord, S.A.; Schladow, S.G. Recent pH and CO2 profiles at Lakes Nyos and Monoun, Cameroon: Implications for the degassing strategy and its numerical simulation. J. Volcanol. Geotherm. Res. 2000, 97, 241–260. [Google Scholar] [CrossRef]
- Kamenetsky, M.B.; Sobolev, A.V.; Kamenetsky, V.S.; Maas, R.; Danyushevsky, L.V.; Thomas, R.; Sobolev, N.V.; Pokhilenko, N.P. Kimberlite melts rich in alkali chlorides and carbonates: A potent metasomatic agent in the mantle. Geology 2004, 32, 845–848. [Google Scholar] [CrossRef]
- Pinti, D.L. The origin and evolution of the oceans. In Lectures in Astrobiology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 83–112. [Google Scholar]
- Hanley, J.J.; Mungall, J.E.; Pettke, T.; Spooner, E.T.C.; Bray, C.J. Fluid and halide melt inclusions of magmatic origin in the Ultramafic and Lower Banded Series, Stillwater Complex, Montana, USA. J. Petrol. 2008, 49, 1133–1160. [Google Scholar] [CrossRef]
- Kump, L.R.; Seyfried, W.E. Hydrothermal Fe fluxes during the Precambrian: Effect of low oceanic sulfate concentrations and low hydrostatic pressure on the composition of black smokers. Earth Planet. Sci. Lett. 2005, 235, 654–662. [Google Scholar] [CrossRef]
- German, C.R.; Von Damm, K.L. Hydrothermal processes. Treatise Geochem. 2003, 6, 625. [Google Scholar]
- White, L.M.; Shibuya, T.; Vance, S.D.; Christensen, L.E.; Bhartia, R.; Kidd, R.; Hoffmann, A.; Stucky, G.D.; Kanik, I.; Russell, M.J. Simulating serpentinization as it could apply to the emergence of life using the JPL hydrothermal reactor. Astrobiology 2020, 20, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, W.E.; Pester, N.J.; Tutolo, B.M.; Ding, K. The Lost City hydrothermal system: Constraints imposed by vent fluid chemistry and reaction path models on subseafloor heat and mass transfer processes. Geochim. Cosmochim. Acta 2015, 163, 59–79. [Google Scholar] [CrossRef]
- Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; 1043 Lebon, G.T.; et al. An off-axis hydrothermal vent field near the Mid-Atlantic Ridge at 30 N. Nature 2001, 412, 145–149. [Google Scholar] [CrossRef]
- Kelley, D.S.; Karson, J.A.; Früh-Green, G.L.; Yoerger, D.R.; Shank, T.M.; Butterfield, D.A.; Hayes, J.M.; Schrenk, M.O.; Olson, E.J.; Proskurowski, G.; et al. A serpentinite-hosted ecosystem: The Lost City hydrothermal field. Science 2005, 307, 1428–1434. [Google Scholar] [CrossRef]
- Lowell, R.P.; Rona, P.A. Seafloor hydrothermal systems driven by the serpentinization of peridotite. Geophys. Res. Lett. 2002, 29, 26. [Google Scholar] [CrossRef]
- Lang, S.Q.; Brazelton, W.J. Habitability of the marine serpentinite subsurface: A case study of the Lost City hydrothermal field. Philos. Trans. R. Soc. A 2020, 378, 20180429. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, B.; Muhling, J.R.; Fischer, W.W. Greenalite Nanoparticles in Alkaline Vent Plumes as Templates for the Origin of Life. Astrobiology 2021, 21, 246–259. [Google Scholar] [CrossRef]
- Mojzsis, S.J.; Harrison, T.M.; Pidgeon, R.T. Oxygen-isotope evidence from ancient zircons for liquid water at the Earth’s sur-face 4300 Myr ago. Nature 2001, 409, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Knauth, L.P. Temperature and salinity history of the Precambrian ocean: Implications for the course of microbial evolution. Palaeogeog. Palaeoclimat. Palaeoecol. 2005, 219, 53–69. [Google Scholar] [CrossRef]
- Robert, F.; Chaussidon, M. A palaeotemperature curve for the Precambrian oceans based on silicon isotopes in cherts. Nature 2006, 443, 969. [Google Scholar] [CrossRef]
- Fisher, C.M.; Vervoort, J.D. Using the magmatic record to constrain the growth of continental crust—The Eoarchean zircon Hf record of Greenland. Earth Planet. Sci. Lett. 2008, 488, 79–91. [Google Scholar] [CrossRef]
- Tartèse, R.; Chaussidon, M.; Gurenko, A.; Delarue, F.; Robert, F. Warm Archean oceans reconstructed from oxygen isotope composition of early-life remnants. Geochem. Perspect. Lett. 2017, 3, 55–65. [Google Scholar] [CrossRef]
- Greenwood, R.C.; Barrat, J.A.; Miller, M.F.; Anand, M.; Dauphas, N.; Franchi, I.A.; Sillard, P.; Starkey, N.A. Oxygen isotopic evidence for accretion of Earth’s water before a high-energy Moon-forming giant impact. Sci. Adv. 2018, 4, eaao5928. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Hall, A.J. The onset and early evolution of life. Geol. Soc. Am. 2006, 198, 1–32. [Google Scholar]
- Russell, M.J.; Hall, A.J. The emergence of life from iron monosulphide bubbles at a submarine hydrothermal redox and pH front. J. Geol. Soc. Lond. 1997, 154, 377–402. [Google Scholar] [CrossRef]
- Yamamoto, M.; Nakamura, R.; Kasaya, T.; Kumagai, H.; Suzuki, K.; Takai, K. Spontaneous and widespread electricity generation in natural deep-sea hydrothermal fields. Angew. Chem. Int. Ed. Engl. 2017, 56, 5725–5728. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Hall, A.J.; Mellersh, A.R. On the dissipation of thermal and chemical energies on the early Earth: The onsets of hydrothermal convection, chemiosmosis, genetically regulated metabolism and oxygenic photosynthesis. In Natural and Laboratory-Simulated Thermal Geochemical Processes; Ikan, R., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 325–388. [Google Scholar]
- Yamagata, Y.; Watanabe, H.; Saitoh, M.; Namba, T. Volcanic production of pyrophosphate and its relevance to prebiotic evolution. Nature 1991, 352, 516–519. [Google Scholar] [CrossRef]
- Milman-Barris, M.S.; Beckett, J.R.; Baker, M.B.; Hofmann, A.E.; Morgan, Z.; Crowley, M.R.; Vielzeuf, D.; Stolper, E. Zoning of phosphorus in igneous olivine. Contr. Min. Petrol. 2008, 155, 739–765. [Google Scholar] [CrossRef]
- Veter, M.; Foley, S.F.; Mertz-Kraus, R.; Groschopf, N. Trace elements in olivine of ultramafic lamprophyres controlled by phlogopite-rich mineral assemblages in the mantle source. Lithos 2017, 292, 81–95. [Google Scholar] [CrossRef]
- Nishizawa, M.; Saito, T.; Makabe, A.; Ueda, H.; Saitoh, M.; Shibuya, T.; Takai, K. Stable Abiotic Production of Ammonia from Nitrate in Komatiite-Hosted Hydrothermal Systems in the Hadean and Archean Oceans. Minerals 2021, 11, 321. [Google Scholar] [CrossRef]
- Sleep, N.H. Geological and geochemical constraints on the origin and evolution of life. Astrobiology 2018, 18, 1199–1219. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.V.; Zybin, K.P. Runaway breakdown and the mysteries of lightning. Phys. Today 2005, 37–43. [Google Scholar] [CrossRef]
- Ducluzeau, A.-L.; van Lis, R.; Duval, S.; Schoepp-Cothenet, B.; Russell, M.J.; Nitschke, W. Was nitric oxide the first strongly oxidizing terminal electron sink? Trends Biochem. Sci. 2009, 34, 9–15. [Google Scholar] [CrossRef]
- Adelman, Z. Technical Memorandum No. 12: Sea Salt and Lightning. June 25, 2012, 773 San Marin Drive, Suite 2115, No-vato, CA 94998 P: 415-899-0700 F: 415-899-0707. Available online: www.environcorp.com (accessed on 13 February 2021).
- Hu, R.; Diaz, H.D. Stability of nitrogen in planetary atmospheres in contact with liquid water. Astrophys. J. 2019, 886, 8. [Google Scholar] [CrossRef]
- Airapetian, V.S.; Barnes, R.; Cohen, O.; Collinson, G.A.; Danchi, W.C.; Dong, C.F.; Del Genio, A.D.; France, K.; Garcia-Sage, K.; Glocer, A.; et al. Impact of space weather on climate and habitability of terrestrial-type exoplanets. Int. J. Astrobiol. 2020, 19, 136–194. [Google Scholar] [CrossRef]
- Liu, W.T. Wind over troubled water. Backscatter 2001, 12, 14. [Google Scholar]
- Mloszewska, A.M.; Pecoits, E.; Cates, N.L.; Mojzsis, S.J.; O’Neil, J.; Robbins, L.J.; Konhauser, K.O. The composition of Earth’s oldest iron formations: The Nuvvuagittuq Supracrustal Belt (Québec, Canada). Earth Planet. Sci. Lett. 2012, 317, 331–342. [Google Scholar] [CrossRef]
- Appel, P.W.U. On the early Archaean Isua iron-formation, west Greenland. Precamb. Res. 1980, 11, 73–87. [Google Scholar] [CrossRef]
- Appel, P.W.U. Mineral occurrences in the 3.6 Ga old Isua supracrustal belt, West Greenland. Develop. Precamb. Geol. 1990, 5, 593–603. [Google Scholar]
- Gäb, F.; Ballhaus, C.; Siemens, J.; Heuser, A.; Lissner, M.; Geisler, T.; Garbe-Schönberg, D. Siderite cannot be used as CO2 sensor for Archaean atmospheres. Geochim. Cosmochim. Acta 2017, 214, 209–225. [Google Scholar] [CrossRef]
- Halevy, I.; Alesker, M.; Schuster, E.M.; Popovitz-Biro, R.; Feldman, Y. A key role for green rust in the Precambrian oceans and the genesis of iron formations. Nat. Geosci. 2017, 10, 135–139. [Google Scholar] [CrossRef]
- Isley, A.E.; Abbott, D.H. Plume-related mafic volcanism and the deposition of banded iron formation. J. Geophys. Res. Solid Earth 1999, 104, 15461–15477. [Google Scholar] [CrossRef]
- Klein, C. Some Precambrian banded iron-formations (BIFs) from around the world: Their age, geologic setting, mineralogy, metamorphism, geochemistry, and origins. Am. Mineral. 2005, 90, 1473–1499. [Google Scholar] [CrossRef]
- Mojzsis, S.J.; Arrhenius, G.; McKeegan, K.D.; Harrison, T.M.; Nutman, A.P.; Friend, C.R.L. Evidence for life on earth before 3,800 million years ago. Nature 1996, 384, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Tosca, N.J.; Guggenheim, S.; Pufahl, P.K. An authigenic origin for Precambrian greenalite: Implications for iron formation and the chemistry of ancient seawater. Geol. Soc. Am. Bull. 2016, 128, 511–530. [Google Scholar] [CrossRef]
- Voosen, P. Ancient Earth was a water world. Sci. Mag. 2021, 1088–1089. [Google Scholar]
- Zhang, S.; Holmes, T.; Lockshin, C.; Rich, A. Spontaneous assembly of a self-complimentary oligopeptide to form a stable macroscopic membrane. Proc. Natl. Acad. Sci. USA 1993, 90, 3334–3338. [Google Scholar] [CrossRef]
- Brack, A.; Orgel, L.E. β structures of alternating polypeptides and their possible prebiotic significance. Nature 1975, 256, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.; Spach, G. Multiconformational synthetic polypeptides. J. Am. Chem. Soc. 1981, 103, 6319–6323. [Google Scholar] [CrossRef]
- Milner-White, E.J.; Russell, M.J. Predicting the conformations of proteins and peptides in early evolution. Biol. Dir. 2008, 3, 3. [Google Scholar] [CrossRef]
- Duval, S.; Baymann, F.; Schoepp-Cothenet, B.; Trolard, F.; Bourrié, G.; Grauby, O.; Branscomb, E.; Russell, M.J.; Nitschke, W. Fougerite: The not so simple progenitor of the first cells. Interface Focus 2019, 9, 20190063. [Google Scholar] [CrossRef]
- Walker, J.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. EMBO. J. 1982, 1, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Saraste, M.; Sibbald, P.R.; Wittinghofer, A. The P-loop—A common motif in ATP and GTP-binding proteins. Trends Biochem. Sci. 1990, 15, 430–434. [Google Scholar] [CrossRef]
- Watson, J.; Milner-White, E.J. A Novel Main-chain Anion-binding Site in Proteins: The Nest. J. Mol. Biol. 2002, 315, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Sühnel, J.; Weiss, M.S. New principles of protein structure: Nests, eggs and what next? Angew. Chem. Int. Ed. Engl. 2002, 41, 4663–4665. [Google Scholar] [CrossRef]
- Bianchi, A.; Giorgi, C.; Ruzza, P.; Toniolo, C.; Milner-White, E.J. A synthetic hexapeptide designed to resemble a proteinaceous P-loop nest is shown to bind inorganic phosphate. Proteins 2012, 80, 1418–1424. [Google Scholar] [CrossRef]
- Pras, M.; Schubert, M.; Zucker-Franklin, D.; Rimon, A.; Franklin, E.C. The characterization of soluble amyloid prepared in water. J. Clin. Investig. 1968, 47, 924–933. [Google Scholar]
- Argudo, P.G.; Giner-Casares, J.J. Folding and self-assembly of short intrinsically disordered peptides and protein regions. Nanoscale Adv. 2021, 3, 1789–1812. [Google Scholar] [CrossRef]
- Hansen, H.C.B.; Koch, C.B. Reduction of nitrate to ammonium by sulphate green rust: Activation energy and reaction mechanism. Clay Min. 1998, 33, 87–101. [Google Scholar] [CrossRef]
- Hansen, H.C.B.; Guldberg, S.; Erbs, M.; Koch, C.B. Kinetics of nitrate reduction by green rusts—effects of interlayer anion and Fe (II): Fe (III) ratio. Appl. Clay Sci. 2001, 18, 81–91. [Google Scholar] [CrossRef]
- Fan, R.; Huh, S.; Yan, R.; Arnold, J.; Yang, P. Gated proton transport in aligned mesoporous silica films. Nat. Mat. 2008, 7, 303–307. [Google Scholar] [CrossRef]
- Mielke, R.E.; Russell, M.J.; Wilson, P.R.; McGlynn, S.E.; Coleman, M.; Kidd, R.; Kanik, I. Design, fabrication, and test of a hydrothermal reactor for origin-of-life experiments. Astrobiology 2010, 10, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Mielke, R.E.; Robinson, K.J.; White, L.M.; McGlynn, S.E.; McEachern, K.; Bhartia, R.; Kanik, I.; Russell, M.J. Iron-sulfide-bearing chimneys as potential catalytic energy traps at life’s emergence. Astrobiology 2011, 11, 933–950. [Google Scholar] [CrossRef] [PubMed]
- Emmez, E.; Yang, B.; Shaikhutdinov, S.; Freund, H.J. Permeation of a single-layer SiO2 membrane and chemistry in confined space. J. Phys. Chem. C 2014, 118, 29034–29042. [Google Scholar] [CrossRef]
- Barge, L.M.; Abedian, Y.; Russell, M.J.; Doloboff, I.J.; Cartwright, J.H.; Kidd, R.D.; Kanik, I. From Chemical Gardens to Fuel Cells: Generation of Electrical Potential and Current Across Self-Assembling Iron Mineral Membranes. Angew. Chem. Int. Ed. Engl. 2015, 54, 8184–8187. [Google Scholar] [CrossRef]
- Silies, L.; Didzoleit, H.; Hess, C.; Stühn, B.; Andrieu-Brunsen, A. Mesoporous thin films, zwitterionic monomers, and iniferter-initiated polymerization: Polymerization in a confined space. Chem. Mat. 2015, 27, 1971–1981. [Google Scholar] [CrossRef]
- Muñoz-Santiburcio, D.; Marx, D. Chemistry in nanoconfined water. Chem. Sci. 2017, 8, 3444–3452. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Santiburcio, D.; Marx, D. Nanoconfinement in Slit Pores Enhances Water Self-Dissociation. Phys. Rev. Lett. 2017, 119, 056002. [Google Scholar] [CrossRef]
- Huang, X.L. Hydrolysis of phosphate esters catalyzed by inorganic iron oxide nanoparticles acting as biocatalysts. Astrobiology 2018, 18, 294–310. [Google Scholar] [CrossRef]
- Huang, X.L. Iron Oxide Nanoparticles: An Inorganic Phosphatase. In Nanocatalysts; IntechOpen: London, UK, 2009. [Google Scholar] [CrossRef]
- Duval, S.; Branscomb, E.; Trolard, F.; Bourrié, G.; Grauby, O.; Heresanu, V.; Schoepp-Cothenet, B.; Zuchan, K.; Russell, M.J.; Nitschke, W. On the why’s and how’s of clay minerals’ importance in life’s emergence. Appl. Clay Sci. 2020, 195, 105737. [Google Scholar] [CrossRef]
- Duval, S.; Zuchan, K.; Baymann, F.; Schoepp-Cothenet, B.; Branscomb, E.; Russell, M.J.; Nitschke, W. Minerals and the emergence of life. In Metals in Life Sciences; Kroneck, P., Sosa Torres, M.E., Eds.; Walter de Gruyter: Berlin, Germany, 2021; pp. 135–157. [Google Scholar]
- Brilmayer, R.; Kübelbeck, S.; Khalil, A.; Brodrecht, M.; Kunz, U.; Kleebe, H.J.; Buntkowsky, G.; Baier, G.; Andrieu-Brunsen, A. Influence of nanoconfinement on the pka of polyelectrolyte functionalized silica mesopores. Adv. Mat. Interf. 2020, 7, 1901914. [Google Scholar] [CrossRef]
- Hooks, M.R.; Webster, P.; Weber, J.M.; Perl, S.; Barge, L.M. Effects of Amino Acids on Iron-Silicate Chemical Garden Precipitation. Langmuir 2020, 36, 5793–5801. [Google Scholar] [CrossRef] [PubMed]
- Ochs, M.; Khalil, A.; Frömling, T.; Andrieu-Brunsen, A. Influence of Wettability on the Impedance of Ion Transport Through Mesoporous Silica Films. Adv. Mat. Interf. 2021, 2002095. [Google Scholar] [CrossRef]
- Cox, B.; Ness, F.; Tuite, M. Analysis of the generation and segregation of propagons: Entities that propagate the [PSI+] prion in yeast. Genetics 2003, 165, 23–33. [Google Scholar] [CrossRef]
- Chernoff, Y.O. Amyloidogenic domains, prions and structural inheritance: Rudiments of early life or recent acquisition? Curr. Opin. Chem. Biol. 2004, 8, 665–671. [Google Scholar] [CrossRef]
- Dixson, J.D.; Azad, R.K. Prions: Roles in development and adaptive evolution. J. Mol. Evol. 2020, 88, 427–434. [Google Scholar] [CrossRef]
- Glover, J.R.; Kowal, A.S.; Schirmer, E.C.; Patino, M.M.; Liu, J.J.; Lindquist, S. Self-seeded fibers formed by Sup35, the protein determinant of [PSI+], a heritable prion-like factor of S. cerevisiae. Cell 1997, 89, 811–819. [Google Scholar] [CrossRef]
- Lupi, O.; Dadalti, P.; Cruz, E.; Sanberg, P.R. Are prions related to the emergence of early life? Med. Hyp. 2006, 67, 1027–1033. [Google Scholar] [CrossRef]
- Maury, C.P.J. Self-propagating β-sheet polypeptide structures as prebiotic informational molecular entities: The amyloid world. Orig. Life Evol. Biosph. 2009, 39, 141–150. [Google Scholar] [CrossRef]
- Maury, C.P.J. Amyloid and the origin of life: Self-replicating catalytic amyloids as prebiotic informational and protometabolic entities. Cell. Mol. Life Sci. 2018, 75, 1499–1507. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.B. Natural pH gradients in hydrothermal alkali vents were unlikely to have played a role in the origin of life. J. Mol. Evol. 2016, 83, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.B. Ancient living organisms escaping from, or imprisoned in, the vents? Life 2017, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Lane, N. Proton gradients at the origin of life. BioEssays 2017, 39, 1600217. [Google Scholar]
- Ross, D.S. It is neither Frankenstein nor a submarine alkaline vent, it is just the Second Law. Bioessays 2018, 40, e1800149. [Google Scholar] [CrossRef]
- Branscomb, E.; Russell, M.J. Why the Submarine Alkaline Vent is the Most Reasonable Explanation for the Emergence of Life. BioEssays 2018, 41, e1800208. [Google Scholar] [CrossRef]
- Wächtershäuser, G. In praise of error. J. Mol. Evol. 2016, 82, 75–80. [Google Scholar] [CrossRef]
- Herschy, B.; Whicher, A.; Camprubi, E.; Watson, C.; Dartnell, L.; Ward, J.; Evans, J.R.; Lane, N. An origin-of life reactor to simulate alkaline hydrothermal vents. J. Mol. Evol. 2014, 79, 213–227. [Google Scholar] [CrossRef]
- Sojo, V.; Ohno, A.; McGlynn, S.E.; Yamada, Y.; Nakamura, R. Microfluidic reactors for carbon fixation under ambient-pressure alkaline-hydrothermal-vent conditions. Life 2019, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.; de Graaf, R.; Strandoo Rodin, M.; Ohno, A.; Lane, N.; McGlynn, S.E.; Yamada, Y.M.A.; Nakamura, R.; Barge, L.M.; Braun, D.; et al. CO2 reduction driven by a pH gradient. Proc. Natl. Acad. Sci. USA 2020, 117, 22873–22879. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Nitschke, W.; Branscomb, E. The inevitable journey to being. Philos. Trans. R. Soc. Lond. B 2013, 368, 0254. [Google Scholar] [CrossRef]
- Hansen, H.C.B.; Borggaard, O.K.; Sørensen,, J. Evaluation of the free energy of formation of Fe(II)-Fe(III) hydroxide-sulphate (green rust) and its reduction of nitrite. Geochim. Cosmochim. Acta 1994, 58, 2599–2608. [Google Scholar]
- Etique, M.; Zegeye, A.; Grégoire, B.; Carteret, C.; Ruby, C. Nitrate reduction by mixed iron (II-III) hydroxycarbonate green rust in the presence of phosphate anions: The key parameters influencing the ammonium selectivity. Water Res. 2014, 62, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Constantino, V.R.; Pinnavaia, T.J. Basic properties of Mg2+1-xAl3+x layered double hydroxides intercalated by carbonate, hydroxide, chloride, and sulfate anions. Inorg. Chem. 1995, 34, 883–892. [Google Scholar] [CrossRef]
- Hansen, H.C.B.; Koch, C.B.; Nancke-Krogh, H.; Borggaard, O.K.; Sørensen, J. Abiotic nitrate reduction to ammonium: Key role of green rust. Environ. Sci. Technol. 1996, 30, 2053–2056. [Google Scholar] [CrossRef]
- Barge, L.M.; Flores, E.; Baum, M.M.; VanderVelde, D.G.; Russell, M.J. Redox and pH gradients drive amino acid synthesis in iron oxyhydroxide mineral systems. Proc. Natl. Acad. Sci. USA 2019, 116, 4828–4833. [Google Scholar] [CrossRef]
- Barge, L.M.; Jones, J.P.; Pagano, J.J.; Martinez, E.; Bescup, J. Three-dimensional analysis of a simulated prebiotic hydrothermal chimney. ACS Earth Space Chem. 2020, 4, 1663–1669. [Google Scholar] [CrossRef]
- Andrault, D.; Bolfan-Casanova, N.; Bouhifd, M.A.; Boujibar, A.; Garbarino, G.; Manthilake, G.; Mezouar, M.; Monteux, J.; Parisiades, P.; Pesce, G. Toward a coherent model for the melting behaviour of the deep Earth’s mantle. Phys. Earth planet. Inter. 2017, 265, 67–81. [Google Scholar] [CrossRef]
- Armstrong, K.; Frost, D.J.; McCammon, C.A.; Rubie, D.C.; Ballaran, T.B. Deep magma ocean formation set the oxidation state of Earth’s mantle. Science 2019, 365, 903–906. [Google Scholar]
- Agrusta, R.; Morison, A.; Agrusta, R.; Labrosse, S.; Deguen, R.; Alboussiére, T.; Tackley, P.J.; Dubuffet, F. Mantle convection interacting with magma oceans. Geophys. J. Internatl. 2020, 220, 1878–1892. [Google Scholar]
- Mills, G.C.; Kenyon, D. The RNA world: A critique. Orig. Des. 1996, 17, 138. [Google Scholar]
- Kurland, C.G. The RNA dreamtime: Modern cells feature proteins that might have supported a prebiotic polypeptide world but nothing indicates that RNA world ever was. Bioessays 2010, 32, 866–871. [Google Scholar]
- Maden, B.E.H. No soup for starters? Autotrophy and the origins of metabolism. Trends Biochem. Sci. 1995, 20, 239–337. [Google Scholar] [CrossRef]
- Wächtershäuser, G. Before enzymes and templates: Theory of surface metabolism. Microbiol. Rev. 1988, 52, 452. [Google Scholar] [CrossRef]
- Wächtershäuser, G. Evolution of the first metabolic cycles. Proc. Natl. Acad. Sci. USA 1990, 87, 200–204. [Google Scholar] [CrossRef]
- Koonin, E.V. The cosmological model of eternal inflation and the transition from chance to biological evolution in the history of life. Biol. Direct 2007, 2, 1–21. [Google Scholar] [CrossRef]
- Sharov, A. Coenzyme world model of the origin of life. Biosystems 2016, 144, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Milner-White, E.J. Protein three-dimensional structures at the origin of life. Interface Focus 2019, 9, 20190057. [Google Scholar]
- Lane, N.; Allen, J.F.; Martin, W. How did LUCA make a living? Chemiosmosis in the origin of life. BioEssays 2010, 32, 271–280. [Google Scholar]
- Jordan, S.; Rammu, H.; Zheludev, I.N.; Hartley, A.M.; Maréchal, A.; Lane, N. Promotion of protocell self-assembly from mixed amphiphiles at the origin of life. Nat. Ecol. Evol. 2019, 3, 1705–1714. [Google Scholar]
- Kandler, O.; König, H. Cell wall polymers in Archaea (Archaebacteria). Cell. Mol. Life Sci. 1998, 54, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Russell, M.J. On the origins of cells: A hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philos. Trans. R. Soc. Lond. B 2003, 358, 59–85. [Google Scholar] [CrossRef]
- Martin, W.; Russell, M.J. On the origin of biochemistry at an alkaline hydrothermal vent. Philos. Trans. R. Soc. Lond. B 2007, 362, 1887–1925. [Google Scholar] [CrossRef] [PubMed]
- Sojo, V.; Pomiankowski, A.; Lane, N. A bioenergetic basis for membrane divergence in archaea and bacteria. PLoS Biol. 2014, 12, e1001926. [Google Scholar] [CrossRef]
- Wilson, E.O. On Human Nature; Cambridge Harvard University Press: Cambridge, UK, 1978. [Google Scholar]
- Russell, M.J. Chimneys, chemical gardens and feldspar horizons+pyrrhotine in some SEDEX deposits: Aspects of alkaline environments of deposition. In Proceedings of the Seventh IAGOD Symposium, Lulea, Sweden, 18–22 August 1986; Zachrisson, E., Ed.; Schweizerbartsche Verlagsbuch handlung: Stuttgart, Germany, 1988; pp. 183–190. [Google Scholar]
- Filtness, M.J.; Butler, I.B.; Rickard, D. The origin of life: The properties of iron sulphide membranes. Trans. Inst. Min. Metall. Sect. B 2003, 112, 171–172. [Google Scholar]
- Rickard, D.; Luther, G.W. Chemistry of Iron Sulfides. Chem. Rev. 2007, 107, 514–562. [Google Scholar] [CrossRef]
- Wang, Q.; Barge, L.M.; Steinbock, O. Production of Pyrophosphate Catalyzed by Mineral Membranes with Steep pH Gradients. Chemistry 2019, 25, 4732–4739. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, W.; Russell, M.J. Just Like the Universe the Emergence of Life had High Enthalpy and Low Entropy Beginnings. J. Cosmol. 2010, 10, 3200–3216. [Google Scholar]
- Wächtershäuser, G. Pyrite formation, the first energy source for life: A hypothesis. Syst. Appl. Microbiol. 1988, 10, 207–210. [Google Scholar] [CrossRef]
- Sleep, N.H.; Zahnle, K.; Neuhoff, P.S. Initiation of clement surface conditions on the early Earth. Proc. Natl. Acad. Sci. USA 2001, 98, 3666–3672. [Google Scholar] [CrossRef]
- Benner, S.A.; Kim, H.J. The case for a Martian origin for Earth life. In Instruments, Methods, and Missions for Astrobiology XVII; International Society for Optics and Photonics: Bellingham, WA, USA, 2015; Volume 9606, p. 9606. [Google Scholar]
- Davies, P. Did Earthlife Come from Mars. In Exobiology: Matter, Energy, and Information in the Origin and Evolution of Life in the Universe; Springer: Dordrecht, The Netherlands, 1998; pp. 241–244. [Google Scholar]
- Kirschvink, J.L.; Weiss, B.P. Mars, panspermia, and the origin of life: Where did it all begin. Palaeontol. Electron. 2002, 4, 8–15. [Google Scholar]
- Barras, C. Water of death: Why the first life was anything but wet. New Sci. 2014, 222, 36–39. [Google Scholar] [CrossRef]
- Von Helmholtz, H. The Origin of the Planetary System. In Selected Writings of Hermann von Helmholtz; Kahl, R., Ed.; Wesleyan University Press: Middletown, CT, USA, 1871; pp. 266–296. [Google Scholar]
- Marx, K. Letter to Pyotr Lavrov. In Marx & Engels by Date—Marxists Internet Archive, 1870–1895, Letter ID:73plv45. Available online: https://www.correspondence.ie/index.php?letters_function=4&letters_search_term=73plv45 (accessed on 9 May 2021).
- Russell, M.J. Prospecting for Life. Interface Focus 2019, 9, 20190050. [Google Scholar] [CrossRef]
- Russell, M.J. Discussion of Shock, E.L. Hydrothermal systems as environments for the emergence of life. In Evolution of Hydrothermal Ecosystems on Earth (and Mars); John Wiley & Son: Hoboken, NJ, USA, 1996; Volume 202, pp. 40–60. [Google Scholar]
- Mitchell, P. Vectorial chemistry and the molecular mechanics of chemiosmotic coupling: Power transmission by proticity. Biochem. Soc. Trans. 1976, 4, 399–430. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, W.; Russell, M.J. Beating the acetyl coenzyme A-pathway to the origin of life. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 0258. [Google Scholar] [CrossRef]
- Russell, M.J. Structural controls of base-metal mineralization in Ireland in relation to continental drift. Trans. Inst. Min. Metall. (Appl. Earth Sci. Sect. B) 1968, 77, B117–B128. [Google Scholar]
- Russell, M.J. The geological environment of post-Caledonian base-metal mineralization in Ireland. Ph.D. Thesis, University of Durham, Durham, UK, 1972. [Google Scholar]
- Russell, M.J. Base-metal mineralization in Ireland and Scotland and the formation of Rockall Trough. In Implications of Continental Drift to the Earth Sciences; Tarling, D.H., Runcorn, S.K., Eds.; Academic Press: London, UK, 1973; Volume 1, pp. 581–597. [Google Scholar]
- Russell, M.J. Downward-excavating hydrothermal cells and Irish-type ore deposits: Importance of an underlying thick Caledonian Prism. Trans. Inst. Min. Metall. (Appl. Earth Sci. Sect. B) 1978, 89, 168–171. [Google Scholar]
- Russell, M.J. Major sediment-hosted zinc + lead deposits: Formation from hydrothermal convection cells that deepen during crustal extension. In Short Course in Sediment-Hosted Stratiform Lead-Zinc Deposits; Short Course Handbook; Mineralogical Association of Canada: Quebec City, QC, Canada, 1983; Volume 8, pp. 251–282. [Google Scholar]
- Russell, M.J.; Solomon, M.; Walshe, J.L. The genesis of sediment-hosted, exhalative zinc + lead deposits. Miner. Depos. 1981, 16, 113–127274. [Google Scholar] [CrossRef]
- Hays, S.J.; Hall, J.; Simmons, G.; Russell, M.J. Sealed microcracks in the Lewisian of NW Scotland: A record of 2 billion years of fluid circulation. Geol. Soc. Lond. 1988, 145, 819–830. [Google Scholar]
- Mills, H.; Halliday, A.N.; Ashton, J.H.; Anderson, I.K.; Russell, M.J. Origin of a giant orebody at Navan, Ireland. Nature 1987, 327, 223–225. [Google Scholar] [CrossRef]
- Fallick, A.E.; Ashton, J.H.; Boyce, A.J.; Ellam, R.M.; Russell, M.J. Bacteria were responsible for the magnitude of the world-class hydrothermal base-metal orebody at Navan, Ireland. Econ. Geol. 2001, 96, 885–890. [Google Scholar] [CrossRef]
- Russell, M.J. Manganese halo surrounding the Tynagh ore deposit, Ireland: A preliminary note. Trans. Inst. Min. Metall. (Appl. Earth Sci. Sect. B) 1974, 83, 65–66. [Google Scholar]
- Russell, M.J. Lithogeochemical environment of the Tynagh base-metal deposit, Ireland, and its bearing on ore deposition. Trans. Inst. Min. Metall. (Appl. Earth Sci. Sect. B) 1975, 84, B128–B133. [Google Scholar]
- Larter, R.C.L.; Boyce, A.J.; Russell, M.J. Hydrothermal pyrite chimneys from the Ballynoe Baryte deposit, Silvermines, County Tipperary, Ireland. Miner. Depos. 1981, 16, 309–318. [Google Scholar] [CrossRef]
- Boyce, A.J.; Coleman, M.L.; Russell, M.J. Formation of fossil hydrothermal chimneys and mounds from Silvermines, Ireland. Nature 1983, 306, 545–550. [Google Scholar] [CrossRef]
- Banks, D.A. A fossil hydrothermal worm assemblage from the Tynagh lead–zinc deposit in Ireland. Nature 1985, 313, 128–131. [Google Scholar] [CrossRef]
- Banks, D.A.; Russell, M.J. Fluid mixing during ore deposition at the Tynagh base-metal deposit, Ireland. Eur. J. Miner. 1992, 4, 921–931. [Google Scholar] [CrossRef]
- Boyce, A.J.; Fallick, A.E.; Fletcher, T.J.; Russell, M.J.; Ashton, J. Detailed sulphur isotope studies of Lower Palaeozoic-hosted pyrite below the giant Navan Zn+Pb mine, Ireland: Evidence of mass transport of crustal S to a sediment-hosted deposit. Miner. Mag. 1994, 58, 109–110. [Google Scholar] [CrossRef]
- Windman, T.; Zolotova, N.; Schwandner, F.; Shock, E.L. Formate as an energy source for microbial metabolism in chemo-synthetic zones of hydrothermal ecosystems. Astrobiology 2007, 7, 873–890. [Google Scholar] [CrossRef] [PubMed]
- Etiope, G.; Schoell, M.; Hosgörmez, H. Abiotic methane flux from the Chimaera seep and Tekirova ophiolites (Turkey): Understanding gas exhalation from low temperature serpentinization and implications for Mars. Earth Planet. Sci. Lett. 2011, 310, 96–104. [Google Scholar]
- Ranjan, S.; Todd, Z.R.; Rimmer, P.B.; Sasselov, D.D.; Babbin, A.R. Nitrogen oxide concentrations in natural waters on early Earth. Geochem. Geophys. Geosyst. 2019, 20, 2021–2039. [Google Scholar] [CrossRef]
- Airapetian, V.S. Extreme space weather in time: Effects on earth. In Extreme Events in Geospace; Buzulukova, N., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 611–632. [Google Scholar]
- Trolard, F.; Bourrié, G. Fougerite a natural layered double hydroxide in gley soil: Habitus, structure, and some properties. In Clay Minerals in Nature: Their Characterization, Modification and Application; Valaskova, M., Martynkova, G.S., Eds.; InTech: Rijeka, Croatia, 2012; pp. 171–188. [Google Scholar]
- Russell, M.J.; Hall, A.J.; Cairns-Smith, A.G.; Braterman, P.S. Submarine hot springs and the origin of life. Nature 1988, 336, 117. [Google Scholar] [CrossRef]
- Russell, M.J.; Daniel, R.M.; Hall, A.J.; Sherringham, J. A hydrothermally precipitated catalytic iron sulphide membrane as a first step toward life. J. Mol. Evol. 1994, 39, 231–243. [Google Scholar] [CrossRef]
- Russell, M.J.; Daia, D.E.; Hall, A.J. The emergence of life from FeS bubbles at alkaline hot springs in an acid ocean. In Thermophiles: The Keys to Molecular Evolution and the Origin of Life? Wiegel, J., Adams, M.W., Eds.; Taylor and Francis: London, UK; Philadelphia, PA, USA, 1998; pp. 77–126. [Google Scholar]
- Zedef, V.; Russell, M.J.; Hall, A.J.; Fallick, A.E. Genesis of Vein-Stockwork and Sedimentary Magnesite and Hydromagnesite Deposits in the Ultramafic Terranes of Southwestern Turkey: A Stable Isotope Study. Econ. Geol. 2000, 95, 429–446. [Google Scholar] [CrossRef]
- Ludwig, K.A.; Shen, C.C.; Kelley, D.S.; Cheng, H.; Edwards, R.L. U–Th systematics and 230Th ages of carbonate chimneys at the Lost City Hydrothermal Field. Geochim. Cosmochim. Acta 2011, 75, 1869–1888. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J.; Rahman, L.; Turner, D.M. Abiotic organic syntheses in deep submarine, alkaline hydrothermal systems catalysed by Fe0, mackinawite, violarite and green rust. In Eleventh Annual VM Goldschmidt Conference Abstract #LPI Contribution No. 1088; Lunar and Planetary Institute: Houston, TX, USA, 2001. [Google Scholar]
- Weber, J.M.; Barge, L.M. Iron-Silicate Chemical Garden Morphology and Silicate Reactivity with Alpha-Keto Acids. ChemSystemsChem 2021, 3, e2000058. [Google Scholar] [CrossRef]
- Branscomb, E.; Russell, M.J. On the beneficent thickness of water. Interface Focus 2019, 9, 20190061. [Google Scholar] [CrossRef]
- Astumian, R.D. Design principles for Brownian molecular machines: How to swim in molasses and walk in a hurricane. Phys. Chem. Chem. Phys. 2007, 9, 5067–5083. [Google Scholar] [CrossRef]
- Astumian, R.D. Coupled transport at the nanoscale: The unreasonable effectiveness of equilibrium theory. Proc. Natl. Acad. Sci. USA 2007, 104, 3–4. [Google Scholar] [CrossRef]
- Arya, S.; Mukhopadhyay, S. Ordered water within the collapsed globules of an amyloidogenic intrinsically disordered protein. J. Phys. Chem. B 2014, 118, 9191–9198. [Google Scholar] [CrossRef]
- Arya, S.; Singh, A.K.; Khan, T.; Bhattacharya, M.; Datta, A.; Mukhopadhyay, S. Water rearrangements upon disorder-to-order amyloid transition. J. Phys. Chem. Lett. 2016, 7, 4105–4110. [Google Scholar] [CrossRef]
- Remsing, R.C.; McKendry, I.G.; Strongin, D.R.; Klein, M.L.; Zdilla, M.J. Frustrated solvation structures can enhance electron transfer rates. J. Phys. Chem. Lett. 2015, 6, 4804–4808. [Google Scholar] [CrossRef]
- Bhullar, R.K.; Zdilla, M.J.; Klein, M.L.; Remsing, R. Effect of water frustration on water oxidation catalysis in the nanoconfined interlayers of layered manganese oxides birnessite and buserite. J. Mat. Chem. A 2021, 11. [Google Scholar]
- Milner-White, E.J.; Russell, M.J. Nests as sites for phosphates and iron-sulfur thiolates in the first membranes: 3 to 6 residue anion-binding motifs (nests). Orig. Life Evol. Biosph. 2005, 35, 19–27. [Google Scholar] [CrossRef]
- Dalal, V.; Arya, S.; Mukhopadhyay, S. Confined Water in Amyloid-Competent Oligomers of the Prion Protein. ChemPhysChem 2016, 17, 2804–2807. [Google Scholar] [CrossRef] [PubMed]
- Camino, J.D.; Gracia, P.; Cremades, N. The role of water in the primary nucleation of protein amyloid aggregation. Biophys. Chem. 2020, 269, 106520. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Nitschke, W. Methane: Fuel or exhaust at the emergence of life? Astrobiology 2017, 17, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Bols, M.L.; Hallaert, S.D.; Snyder, B.E.; Devos, J.; Plessers, D.; Rhoda, H.M.; Dusselier, M.; Schoonheydt, R.A.; Pierloot, K.; Solomon, E.I.; et al. Spectroscopic identification of the α-Fe/α-O active site in Fe-CHA zeolite for the low-temperature activation of the methane C–H bond. J. Am. Chem. Soc. 2018, 140, 12021–12032. [Google Scholar] [CrossRef]
- Elitzur, A.C. Let there be life: Thermodynamic reflections on biogenesis and evolution. J. Theor. Biol. 1994, 168, 429–459. [Google Scholar] [CrossRef]
- Qiao, L.; Duan, G.; Zhang, S.; Ren, Y.; Sun, Y.; Tang, Y.; Wan, P.; Pang, R.; Chen, Y.; Russell, A.G.; et al. Electrochemical ammonia synthesis catalyzed with a CoFe layered double hydroxide–A new initiative in clean fuel synthesis. J. Clean. Prod. 2020, 250, 119525. [Google Scholar]
- Schoepp-Cothenet, B.; van Lis, R.; Atteia, A.; Baymann, F.; Capowiez, L.; Ducluzeau, A.-L.; Duval, S.; ten Brink, F.; Russell, M.J.; Nitschke, W. On the universal core of bioenergetics. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Erastova, V.; Degiacomi, M.T.; Fraser, D.G.; Greenwell, H.C. Mineral surface chemistry control for origin of prebiotic peptides. Nat. Commun. 2017, 8, 1–9. [Google Scholar]
- Cardenas, M.B.; Rodolfo, R.S.; Lapus, M.R.; Cabria, H.B.; Fullon, J.; Gojunco, G.R.; Breecker, D.O.; Cantarero, D.M.; Evaristo, J.; Siringan, F.P.; et al. Submarine groundwater and vent discharge in a volcanic area associated with coastal acidification. Geophys. Res. Lett. 2020, 47, e2019GL085730. [Google Scholar] [CrossRef]
- Feng, Y.; Ovalle, M.; Seale, J.S.; Lee, C.K.; Kim, D.J.; Astumian, R.D.; Stoddart, J.F. Molecular Pumps and Motors. J. Am. Chem. Soc. 2021, 143, 5569–5591. [Google Scholar]
- Astumian, R.D.; Mukherjee, S.; Warshel, A. The physics and physical chemistry of molecular machines. ChemPhysChem 2016, 17, 1719–1741. [Google Scholar] [CrossRef] [PubMed]
- Kellosalo, J.; Kajander, T.; Kogan, K.; Pokharel, K.; Goldman, A. The structure and catalytic cycle of a sodium-pumping pyrophosphatase. Science 2012, 337, 473–476. [Google Scholar]
- Baykov, A.A. Energy Coupling in Cation-Pumping Pyrophosphatase—Back to Mitchell. Front. Plant Sci. 2020, 11, 107. [Google Scholar]
- Holmes, A.O.; Kalli, A.C.; Goldman, A. The function of membrane integral pyrophosphatases from whole organism to single molecule. Front. Mol. Biosci. 2019, 6, 132. [Google Scholar] [CrossRef] [PubMed]
- Astumian, R.D. Stochastically pumped adaptation and directional motion of molecular machines. Proc. Natl Acad. Sci. USA 2018, 115, 9405–9413. [Google Scholar] [CrossRef]
- Carter, C.W. Escapement mechanisms: Efficient free energy transduction by reciprocally-coupled gating. Proteins 2020, 88, 710–717. [Google Scholar] [CrossRef]
- Carter, C.W.; Wills, P.R. Reciprocally-Coupled Gating: Strange Loops in Bioenergetics, Genetics, and Catalysis. Biomolecules 2021, 11, 265. [Google Scholar] [CrossRef]
- Oster, G. Darwin’s motors. Nature 2002, 417, 25. [Google Scholar] [CrossRef]
- Hoffmann, P.M. Life’s Ratchets; Basic Books: New York, NY, USA, 2012. [Google Scholar]
- Branscomb, E.; Russell, M.J. Turnstiles and bifurcators: The disequilibrium converting engines that put metabolism on the road. Biochim. Biophys. Acta 2013, 1827, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Wander, M.C.; Rosso, K.M.; Schoonen, M.A. Structure and charge hopping dynamics in green rust. J. Phys. Chem. C 2007, 111, 11414–11423. [Google Scholar] [CrossRef]
- Taglialegna, A.; Lasa, I.; Valle, J. Amyloid structures as biofilm matrix scaffolds. J. Bacteriol. 2016, 198, 2579–2588. [Google Scholar] [CrossRef] [PubMed]
- Pfammatter, M.; Andreasen, M.; Meisl, G.; Taylor, C.G.; Adamcik, J.; Bolisetty, S.; Sánchez-Ferrer, A.; Klenerman, D.; Dobson, C.M.; Mezzenga, R.; et al. Absolute quantification of amyloid propagons by digital microfluidics. Analyt. Chem. 2017, 89, 12306–12313. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Steinbock, O. Materials synthesis and catalysis in microfluidic devices: Prebiotic chemistry in mineral membranes. ChemCatChem 2020, 1, 63–74. [Google Scholar] [CrossRef]
- DelloStritto, M.J.; Thenuwara, A.C.; Klein, M.L.; Strongin, D.R. Effect of Interlayer Co2+ on Structure and Charge Transfer in NiFe Layered Double Hydroxides. J. Phys. Chem. C 2019, 123, 13593–13599. [Google Scholar] [CrossRef]
- Ding, Y.; Cartwright, J.H.; Cardoso, S.S. Intrinsic concentration cycles and high ion fluxes in self-assembled precipitate membranes. Interface Focus 2019, 9, 20190064. [Google Scholar]
- de Herrera, A.G.; Markert, T.; Trixler, F. Abiotic nanofluidic environments induce prebiotic condensation in water. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Kulkarni, M.B.; Goel, S.G. Microfluidic devices for synthesizing nanomaterials–A review. Nano Express 2020, 1, 032004. [Google Scholar]
- Möller, F.M.; Kriegel, F.; Kieß, M.; Sojo, V.; Braun, D. Steep pH gradients and directed colloid transport in a microfluidic alkaline hydrothermal pore. Angew. Chem. Int. Ed. Engl. 2017, 56, 2340–2344. [Google Scholar]
- Semenov, S.; Semenov, S.N.; Kraft, L.J.; Ainla, A.; Zhao, M.; SBaghbanzadeh, M.; Campbell, V.E.; Kang, K.; Fox, J.M.; Whitesides, G.M. Autocatalytic, bistable, oscillatory networks of biologically relevant organic reactions. Nature 2016, 537, 656–660. [Google Scholar]
- Bose, S.K.; Lawrence, C.P.; Liu, Z.; Makarenko, K.S.; van Damme, R.M.; Broersma, H.J.; van der Wiel, W.G. Evolution of a designless nanoparticle network into reconfigurable Boolean logic. Nat. Nanotech. 2015, 10, 1048–1052. [Google Scholar] [CrossRef]
- Kotopoulou, E.; Lopez-Haro, M.; Calvino Gamez, J.J.; García-Ruiz, J.M. Nanoscale anatomy of iron-silica self-organized membranes: Implications for prebiotic chemistry. Angew. Chem. Int. Ed. Engl. 2021, 133, 1416–1422. [Google Scholar]
- Roldan, A.; Hollingsworth, N.; Roffey, A.; Islam, H.U.; Goodall, J.B.; Catlow, C.R.; Darr, J.A.; Bras, W.; Sankar, G.; Holt, K.B.; et al. Bio-inspired CO2 conversion by iron sulfide catalysts under sustainable conditions. Chem. Commun. 2015, 51, 7501–7504. [Google Scholar] [CrossRef]
- Santos-Carballal, D.; Roldan, A.; De Leeuw, N.H. CO2 reduction to acetic acid on the greigite Fe3S4{111} surface. Faraday Discuss. 2020. [Google Scholar] [CrossRef]
- Katz, J.E.; Zhang, X.; Attenkofer, K.; Chapman, K.W.; Frandsen, C.; Zarzycki, P.; Rosso, K.M.; Falcone, R.W.; Waychunas, G.A.; Gilbert, B. Electron small polarons and their mobility in iron (oxyhydr) oxide nanoparticles. Science 2012, 337, 1200–1203. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.; Pasalic, J.; Hermis, N.; Barge, L.M. Chemical Gardens as Electrochemical Systems: In Situ Characterization of Simulated Prebiotic Hydrothermal Vents by Impedance Spectroscopy. ChemPlusChem. 2020, 85, 2619–2628. [Google Scholar] [CrossRef]
- Barge, L.M.; Flores, E.; VanderVelde, D.G.; Weber, J.M.; Baum, M.M.; Castonguay, A. Effects of Geochemical and Environmental Parameters on Abiotic Organic Chemistry Driven by Iron Hydroxide Minerals. J. Geophys. Res. Planets 2020, 125, e2020JE006423. [Google Scholar] [CrossRef]
- Einsle, O.; Messerschmidt, A.; Huber, R.; Kroneck, P.M.; Neese, F. Mechanism of the six-electron reduction of nitrite to ammonia by cytochrome c nitrite reductase. J. Am. Chem. Soc. 2002, 124, 11737–11745. [Google Scholar]
- Arrhenius, G.O. Crystals and life. Helv. Chim. Acta 2003, 86, 1569–1586. [Google Scholar] [CrossRef]
- Greenwell, H.C.; Coveney, P.V. Layered double hydroxide minerals as possible prebiotic information storage and transfer compounds. Orig. Life Evol. Biosph. 2006, 36, 13–37. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, S.J.; Beckett, P. Probing complexity: Thermodynamics and computational mechanics approaches to origins studies. Interface Focus 2019, 9, 20190058. [Google Scholar] [CrossRef]
- Pask, G. Physical analogues to the growth of a concept. Mechanization of Thought Processes. In Proceedings of the Symposium 10, National Physical Laboratory, 24–27 November 1958; H.M.S.O.: London, UK, 1958; pp. 765–794. [Google Scholar]
- Stoica, A.; Zebulum, R.S.; Keymeulen, D. Mixtrinsic evolution. In Proceedings of the Third International Conference on Evolvable Systems: From Biology to Hardware (ICES2000), Edinburgh, UK, 17–19 April 2000; Volume 1801, pp. 208–217. [Google Scholar]
- Miller, J.F.; Downing, K.; Tufte, G. Evolution in materio: Looking beyond the silicon box. Proc. NASA/DoD Evolvable Hardware Workshop 2002, 167–176. [Google Scholar] [CrossRef]
- Harding, S.L.; Miller, J.F.; Rietman, E.A. Evolution in materio: Exploiting the physics of materials for computation. arXiv 2006, arXiv:cond-mat/0611462. [Google Scholar]
- Stepney, S. The neglected pillar of material computation. Phys. D 2008, 237, 1157–1164. [Google Scholar] [CrossRef]
- Stepney, S. Programming unconventional computers: Dynamics, development, self-reference. Entropy 2012, 14, 1939–1952. [Google Scholar] [CrossRef]
- Horsman, C.; Stepney, S.; Wagner, R.C.; Kendon, V. When does a physical system compute? Proc. R. Soc. A 2014, 470, 20140182. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.H.; Folven, E.; Tufte, G. Computation in artificial spin ice. In Artificial Life Conference Proceedings; MIT Press: Cambridge, MA, USA, 2018; pp. 15–22. [Google Scholar]
- Adamatzky, A. Reaction-diffusion computing. In Encyclopedia of Complexity and Systems Science; Meyers, R.A., Ed.; Springer: New York, NY, USA, 2009; pp. 7548–7565. [Google Scholar]
- Yu, L.; Mishra, I.K.; Xie, Y.; Zhou, H.; Sun, J.; Zhou, J.; Ni, Y.; Luo, D.; Yu, F.; Yu, Y.; et al. Ternary Ni2(1-x)Mo2xP nanowire arrays toward efficient and stable hydrogen evolution electrocatalysis under large-current-density. Nano Energy 2018, 53, 492–500. [Google Scholar] [CrossRef]
- Arrabito, G.; Pezzilli, R.; Prestopino, G.; Medaglia, P.G. Layered double hydroxides in bioinspired nanotechnology. Crystals 2020, 10, 602. [Google Scholar] [CrossRef]
- Epstein, I.R.; Xu, B. Reaction–diffusion processes at the nano-and microscales. Nat. Nanotech. 2016, 11, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Fracchia, M.; Visibile, A.; Ahlberg, E.; Vertova, A.; Minguzzi, A.; Ghigna, P.; Rondinini, S. α-and γ-FeOOH: Stability, Reversibility, and Nature of the Active Phase under Hydrogen Evolution. ACS Appl. Energy Mat. 2018, 1, 1716–1725. [Google Scholar] [CrossRef]
- Mann, S. Life as a nanoscale phenomenon. Angew. Chem. Int. Ed. Engl. 2008, 4, 5306–5320. [Google Scholar] [CrossRef] [PubMed]
- Paolella, A.; George, C.; Povia, M.; Zhang, Y.; Krahne, R.; Gich, M.; Genovese, A.; Falqui, A.; Longobardi, M.; Guardia, P.; et al. Charge transport and electrochemical properties of colloidal greigite (Fe3S4) nanoplatelets. Chem. Mater. 2011, 23, 3762–3768. [Google Scholar] [CrossRef]
- Sano, Y.; Kyono, A.; Yoneda, Y.; Isaka, N.; Takagi, S.; Yamamoto, G.I. Structure changes of nanocrystalline mackinawite under hydrothermal conditions. J. Mineral. Petrol. Sci. 2020, 190903. [Google Scholar] [CrossRef]


{kind=link}
| Effects | Descriptions | References |
|---|---|---|
| Solar luminosity post solar wind | 72% of present flux | [75] |
| Solar radiation (UV and X-ray) | Intense: ~100 + times present | [76,77] |
| Earth–Moon distance; Earth’s spin and length of day and tides | <10% of present day; Estimates of length of day from 2 to 10 h; ~20 m tidal amplitude | [78,79,80,81,82,83,84,85,86,87,88,89,90,91] |
| Bombardment and tsunamis | Heavy but not totally vaporizing (negative feedback from heightening atmospheric pressure) | [92,93,94,95,96,97] |
| Maximum height of ocean plateaux above by mantle plumes. | ~1000 km | [98,99,100,101,102,103,104,105,106,107] |
| Ocean depth | 4–6 km | [108,109,110,111,112,113,114,115,116,117] |
| Redox state | Upper mantle buffered at quartz-fayalite-magnetite | [62,63,118,119,120] |
| Atmosphere post solar wind | CO2, N2, H2O, > SO2 >> CO, NOx | [121,122,123,124,125,126,127,128,129,130,131,132,133] |
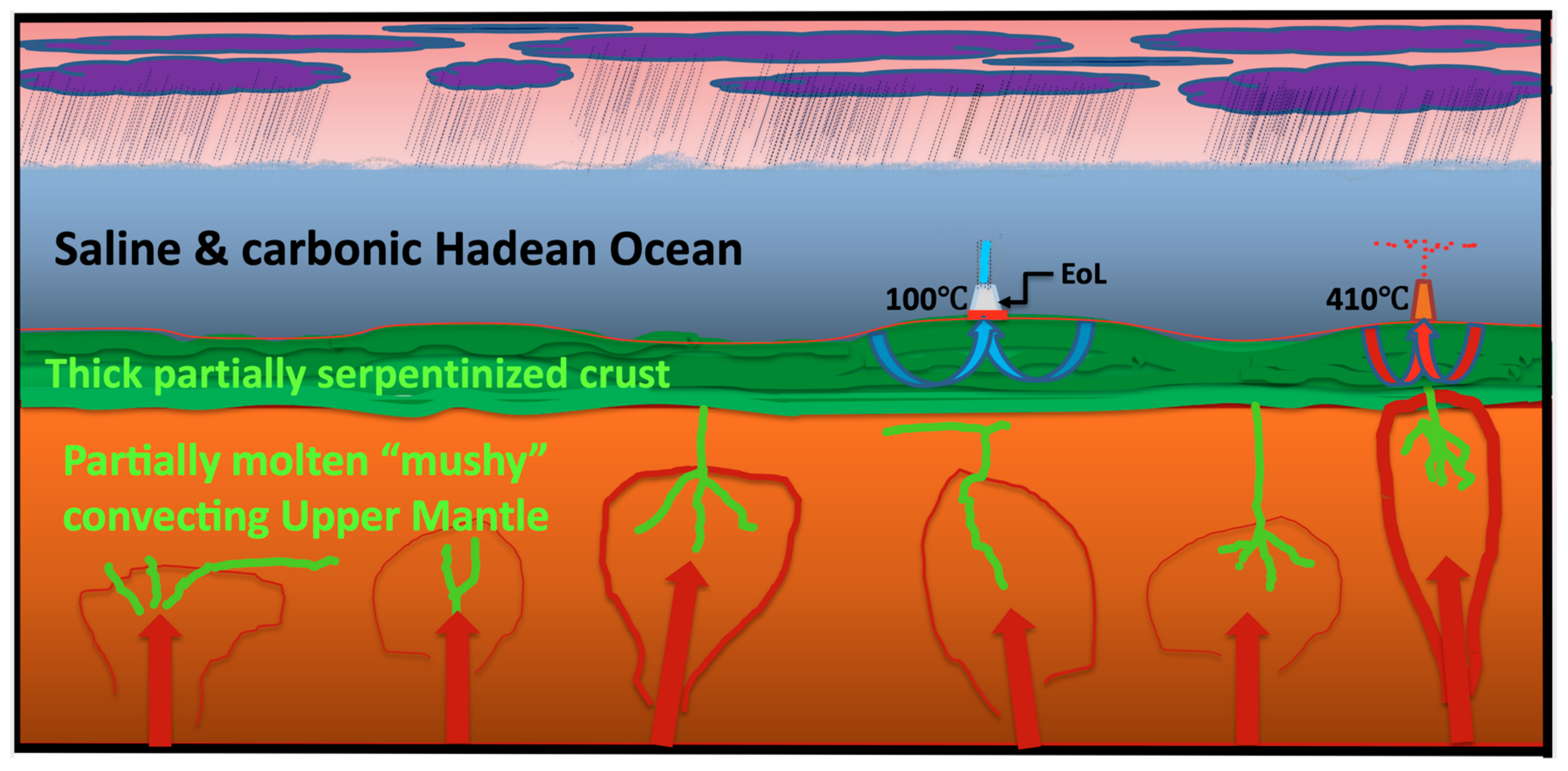
| Ocean chemistry | Saline, CO2, NO3−, NO2− + metal ions | [134,135,136,137,138,139,140] |
| Magma-driven submarine springs | Acidic, ~400 °C | [141] |
| Direct contribution of ~400 °C solution to Hadean Ocean | Fe2+, Mg2+, Mn2+, Zn2+, Co2+, Ni2+, H2S, H2, PO4−, CH4 | [142] |
| Serpentinization-driven submarine springs | Alkaline, ~120 °C | [143,144,145,146,147] |
| Direct contribution of ~120 °C alkaline spring to the hydrothermal mound | H2, CH4, HS−, HCOO− > [Fe2S2(MoS4)2]2−/4− | [142,143,144,145,146,147,148,149] |
| Ocean T & pH and chemistry | Strongly carbonic and saline 25 to <85 °C? pH 4.5 to 5.5 with minor nitrate, transition metals in solution fed from ~400 °C springs | [129,141,150,151,152,153,154,155] |
| The Earth electronic and protonic ~1 volt battery | Eh of H2 v. H2O at delta pH 4 to 5 | [136,156,157,158,159] |
| Olivine source of pyrophosphate delivered to ocean via vulcanism | Hydrolysis of volcanic P4O10 to produce P3O93− and P4O124− | [160,161,162] |
| Lightning | Produces NO from CO2 + N2 | [131,163,164,165,166,167,168,169] |
| Wind speed (cf. “Roaring Forties”) | 12 ms–1 estimate | [170] |
| Wave height | 10 m estimate | [86,170] |
| Chemical sediments | Banded iron formation, fougerite, chert, greenalite, mackinawite | [171,172,173,174,175,176,177,178,179] |
| Prediction/Expectation | References | Tests | References |
|---|---|---|---|
| Hydrothermal circulation during rift tectonics generates metal sulfide mineral deposits. | [268] | Successful field test, discovery of giant base metal deposit, Navan, Ireland | [269,270] |
| Seawater-derived Downward hydrothermal convection driven by crustal heat and exothermic reactions | [271,272,273,274] | Stratigraphic, structural, tectonic and lithochemical field work; Lead isotope analyses | [275,276,277] |
| Some hydrothermal minerals precipitate on sea-floor | [269,270,271,272,273] | Delineation of extensive Mn aureole centered on Irish ore deposit led to the first discovery of fossil hydrothermal chimneys formed through mixing with seawater | [277,278,279,280,281,282] |
| Some sulfur derived from crustal sources | [157,273,283] | Isotopic analysis reveals crustal source as do hydrothermal experiments | [196,197,276] |
| Sulfide dissolves in alkaline hydrothermal solution | [157] | Lab demonstrations | [196,197,225] |
| Serpentinization reaction to formate | [284,285,286,287,288,289,290,291] | Lab demonstration | [226] |
| Serpentinization reaction produces H2 >>CH4, although CH4 is entrained from oceanic crust | [292] | Prompts amendment to the AVT, viz., to the denitrifying methanotrophic acetogenesis | [267] |
| Lightning and space weather radiation produces NOx that rapidly dissolve as nitrate/nitrite in Hadean Ocean | [126] | Theoretic studies generally support this hypothesis though not those of Ranjan et al. 2019 [286] | [131] |
| Source of ammonia at vent from nitrate/nitrite reduction | [126,163] | Eight electron reduction of nitrate to ammonia with green rust | [193,194,230] |
| Further reduction of NOx to hydrazine N2H4 | [80] | Awaiting test | |
| Off-ridge submarine vents will be moderate temperature, H2-bearing, alkaline and long-lived (≥105 years) and would have been the site of life’s emergence | [36,136,157,264,293] | Discovery of Lost City moderate temperature alkaline hydrothermal vent in North Atlantic Ocean | [146,265] |
| Green rust, mackinawite/greigite, amorphous silica barrier/ membrane | [36,39,72,294] | Successful lab demonstration | [36,159,197,232,251,295] |
| Eh and pH gradients ~700 mV and 4–5 units pH (~300 mV) to meet electronic and protonic requirements ~1 V | [157] | FeS barriers hold a 700 mV and a 5 unit pH disequilibrium in lab test | [199,231,254] |
| CO2 reduction forced by H2 and delta pH 4 units | [157] | Chemical disequilibria as per Nernst equation | [226] |
| The immateriality of the “water problem” in nano-confinement | [72,296,297,298,299,300,301,302,303,304,305] | Lab and molecular dynamic simulations | [201,202] |
| Aminations of carboxylic acids | [72] | Lab demonstrations. Amination of pyruvate to alanine | [231] |
| Green rust as proto- pyrophosphatase | [228] | Pi + Pi → PPi to equilibrium in microfluidic reactor | [254] |
| ΔpH as pmf | [63,156,157] | Undemonstrated | |
| Oxidation of methane in green rust | [72,267,306] | Undemonstrated, pending | Cf. [307] |
| Theoretical polymerization of amino acids in nano-confined water to produce peptide membranes necessarily pre-LUCA | [72] | Undemonstrated, waiting experiment | |
| Expansion from the vent via ocean floor to produce the first deep biosphere | [156] | Hypothesis |
| Surface Pond | References | Submarine Alkaline Vent | References | |
|---|---|---|---|---|
| “Free energy” | UV, reverse 2 pH unit pmf | [7,16,59,60,68] | Electrochemical gradients, natural 4 pH unit pmf | [156,157,199,219,226,231,254,291] |
| Electron donors | UV radiation, reduced sulfur & organic compounds, Fe2+ | [3,10,30,65] | Fe2+, H2, ē, CH4, HCOO- | [5,36,39,63,72,156,157,247,248,267,284,291,306,309,310] |
| Electron acceptors | Ferricyanide insidevesicle | [9] | Ambient Fe3+, NO, NO2−, NO3− (CO2) | [39,131,157,166,291] |
| Initial boundary | Lipids | [7,49,51,64] | Green rust, FeS, silica | [36,37,38,39,72,157,291] |
| Organic takeover | Lipids | [7,9] | Peptides | [181,184,185,190,191,192,215,216,237,311] |
| Primary ingredients | Lipids HCN, CH3N, H2S, H2O2, quinones, ferricyanide | [9] | CO2, H2, CH4, H2O, NO3, NO, NH2, NH3, HPO43−, HS−, Fe2+, Ni2+, Co2+, Mo4+/6+ | [116,129,130,131,132,133,134,135,136,137,138,143,196,197,225,291,312] |
| Other suggestions orderivatives | Acrolein, 2-aminooxazol, quinones, ferricyanide, RNA, DNA, NADH, FMN | [11,16,30,60,64,65,66,67] | NH3 + carboxylic acids → amino acids, N2H4 → heterocycles, e.g., pyrazoles, imides, NAD(P), flavins, quinones | [185,193,194,230,310] |
| Disequilibria conversion mechanisms | Wet/dry cycling aggregation | [7] | Visco-elastic allosteric conformational changes/binding change mechanism/pumping/gating/ electrostatic effects | [5,47,71,72,181,182,183,184,185,186,187,188,189,190,191,192,221,228,310,313,314,315,316,317,318,319,320,321,322,323,324] |
| Condensations | Wet/dry cycling | [9,20,21,24,57] | Nanoconfined water in green rust interlayers, silica, mackinawite | [72,80,88,89,90,96,97,100,101,102,103,104,123,125,302] |
| Reproduction | RNA world | [9,17,22,51,64] | Amyloid peptide | [72,211,212,213,214,215,216,325,326] |
| Waste disposal | None considered | --- | In alkaline spring effluent | [36,47,72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russell, M.J. The “Water Problem”(sic), the Illusory Pond and Life’s Submarine Emergence—A Review. Life 2021, 11, 429. https://doi.org/10.3390/life11050429
Russell MJ. The “Water Problem”(sic), the Illusory Pond and Life’s Submarine Emergence—A Review. Life. 2021; 11(5):429. https://doi.org/10.3390/life11050429
Chicago/Turabian StyleRussell, Michael J. 2021. "The “Water Problem”(sic), the Illusory Pond and Life’s Submarine Emergence—A Review" Life 11, no. 5: 429. https://doi.org/10.3390/life11050429
APA StyleRussell, M. J. (2021). The “Water Problem”(sic), the Illusory Pond and Life’s Submarine Emergence—A Review. Life, 11(5), 429. https://doi.org/10.3390/life11050429