

Modulation of Small RNA Signatures by Astrocytes on Early Neurodegeneration Stages; Implications for Biomarker Discovery

 , ,
, ,  , , , and
, , , and
Abstract

1. Introduction
2. Astrocyte Relevance on Early Neurodegeneration Stages
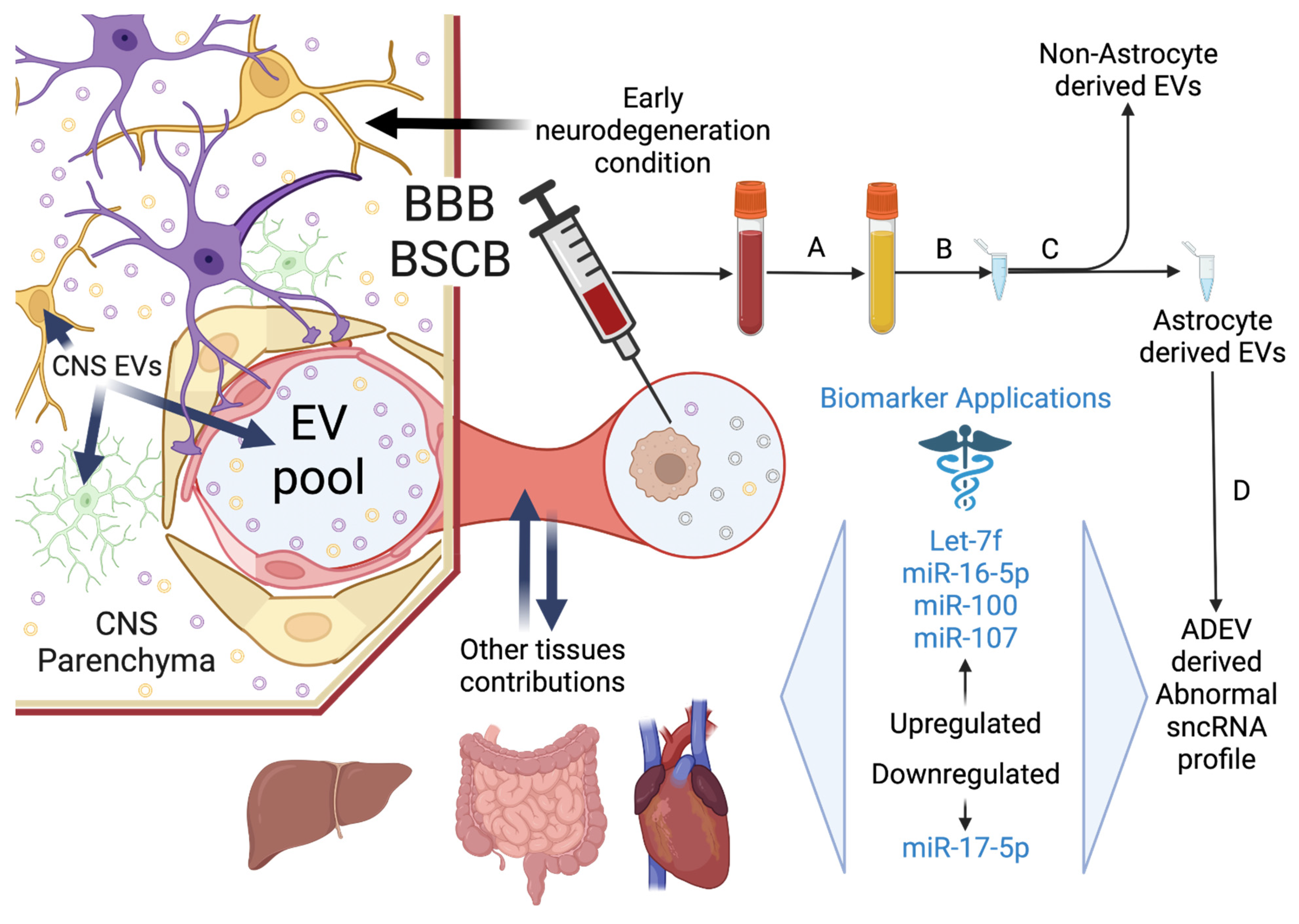
3. Astrocyte Derived Extracellular sncRNA Signatures as Biomarkers for Early Neurodegeneration
4. Relevance of Carriers of sncRNA in Astrocyte Derived Extracellular sncRNA Biomarker Design for Neurodegenerative Disease (ND)
5. Astrocyte Derived Extracellular Vesicle (ADEV) Derived sncRNA Effect on Microglia
6. Astrocyte Derived Extracellular Vesicle (ADEV) Derived sncRNA Effect on Neurons
7. Astrocyte Derived Extracellular Vesicle (ADEV) Derived sncRNA Effect on Other Astrocytes
8. Astrocyte Derived Extracellular Vesicle (ADEV) Derived sncRNA Effect on Peripheral Cells
9. Limitations of Small Non-Coding RNA Signatures of Astrocyte Derived Extracellular Vesicles (ADEVs) as Biomarkers of Early Neurodegeneration Cells
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Catanesi, M.; D’Angelo, M.; Tupone, M.G.; Benedetti, E.; Giordano, A.; Castelli, V.; Cimini, A. MicroRNAs Dysregulation and Mitochondrial Dysfunction in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 5986. [Google Scholar] [CrossRef] [PubMed]
- Feigin, V.L.; Nichols, E.; Alam, T.; Bannick, M.S.; Beghi, E.; Blake, N.; Culpepper, W.J.; Dorsey, E.R.; Elbaz, A.; Ellenbogen, R.G.; et al. Global, Regional, and National Burden of Neurological Disorders, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef]
- Erkkinen, M.G.; Kim, M.O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2018, 10, a033118. [Google Scholar] [CrossRef] [PubMed]
- Sandor, C.; Honti, F.; Haerty, W.; Szewczyk-Krolikowski, K.; Tomlinson, P.; Evetts, S.; Millin, S.; Keane, T.; McCarthy, S.A.; Durbin, R.; et al. Whole-Exome Sequencing of 228 Patients with Sporadic Parkinson’s Disease. Sci. Rep. 2017, 7, 41188. [Google Scholar] [CrossRef] [PubMed]
- Syeda, T.; Cannon, J.R. Environmental Exposures and the Etiopathogenesis of Alzheimer’s Disease: The Potential Role of BACE1 as a Critical Neurotoxic Target. J. Biochem. Mol. Toxicol. 2021, 35, e22694. [Google Scholar] [CrossRef]
- Steenhof, M.; Stenager, E.; Nielsen, N.M.; Kyvik, K.; Möller, S.; Hertz, J.M. Familial Multiple Sclerosis Patients Have a Shorter Delay in Diagnosis than Sporadic Cases. Mult. Scler. Relat. Disord. 2019, 32, 97–102. [Google Scholar] [CrossRef]
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s Disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef]
- Hao, N.; Wang, Z.; Liu, P.; Becker, R.; Yang, S.; Yang, K.; Pei, Z.; Zhang, P.; Xia, J.; Shen, L.; et al. Acoustofluidic Multimodal Diagnostic System for Alzheimer’s Disease. Biosens. Bioelectron. 2022, 196, 113730. [Google Scholar] [CrossRef]
- Ebrahimkhani, S.; Vafaee, F.; Young, P.E.; Hur, S.S.J.; Hawke, S.; Devenney, E.; Beadnall, H.; Barnett, M.H.; Suter, C.M.; Buckland, M.E. Exosomal MicroRNA Signatures in Multiple Sclerosis Reflect Disease Status. Sci. Rep. 2017, 7, 14293. [Google Scholar] [CrossRef]
- Prabhakar, P.; Retnaswami, S.; Christopher, R. Circulating MicroRNAs as Potential Biomarkers for the Identi Fi Cation of Vascular Dementia Due to Cerebral Small Vessel Disease. Age Ageing 2017, 46, 861–864. [Google Scholar] [CrossRef]
- Hansson, O. Biomarkers for Neurodegenerative Diseases. Nat. Med. 2021, 27, 954–963. [Google Scholar] [CrossRef]
- Brazaca, L.C.; Sampaio, I.; Zucolotto, V.; Janegitz, B.C. Applications of Biosensors in Alzheimer’s Disease Diagnosis. Talanta 2020, 210, 120644. [Google Scholar] [CrossRef]
- Swarbrick, S.; Wragg, N.; Ghosh, S.; Stolzing, A. Systematic Review of MiRNA as Biomarkers in Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 6156–6167. [Google Scholar] [CrossRef]
- Katsuno, M.; Sahashi, K.; Iguchi, Y.; Hashizume, A. Preclinical Progression of Neurodegenerative Diseases. Nagoya J. Med. Sci. 2018, 80, 289–298. [Google Scholar] [CrossRef]
- Le, W.; Dong, J.; Li, S.; Korczyn, A.D. Can Biomarkers Help the Early Diagnosis of Parkinson’s Disease? Neurosci. Bull. 2017, 33, 535–542. [Google Scholar] [CrossRef]
- Nandi, S.K.; Singh, D.; Upadhay, J.; Gupta, N.; Dhiman, N.; Mittal, S.K.; Mahindroo, N. Identification of Tear-Based Protein and Non-Protein Biomarkers: Its Application in Diagnosis of Human Diseases Using Biosensors. Int. J. Biol. Macromol. 2021, 193, 838–846. [Google Scholar] [CrossRef]
- Kim, K.; Kim, M.J.; Kim, D.W.; Kim, S.Y.; Park, S.; Park, C.B. Clinically Accurate Diagnosis of Alzheimer’s Disease via Multiplexed Sensing of Core Biomarkers in Human Plasma. Nat. Commun. 2020, 11, 119. [Google Scholar] [CrossRef]
- Tan, R.; Wang, Y.; Mi, X.; Li, H.; Tu, Y. A Dual-Screening Electrochemiluminescent Aptasensor Based on a Mesoporous Silica Nano-Sieve for Specific Detection of Amyloid-β Monomer. Sens. Actuators B Chem. 2022, 352 Pt 2, 131065. [Google Scholar] [CrossRef]
- Bateman, R.J.; Wen, G.; Morris, J.C.; Holtzman, D.M. Fluctuations of CSF Amyloid-β Levels. Neurology 2007, 68, 666–669. [Google Scholar] [CrossRef]
- Knopman, D.S.; Amieva, H.; Petersen, R.C.; Chételat, G.; Holtzman, D.M.; Hyman, B.T.; Nixon, R.A.; Jones, D.T. Alzheimer Disease. Nat. Rev. Dis. Prim. 2021, 7, 33. [Google Scholar] [CrossRef]
- Souza, D.G.; Almeida, R.F.; Souza, D.O.; Zimmer, E.R. The Astrocyte Biochemistry. Semin. Cell Dev. Biol. 2019, 95, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Colangelo, A.M.; Alberghina, L.; Papa, M. Astrogliosis as a Therapeutic Target for Neurodegenerative Diseases. Neurosci. Lett. 2014, 565, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Mozafari, N.; Ashrafi, H.; Azadi, A. Targeted Drug Delivery Systems to Control Neuroinflammation in Central Nervous System Disorders. J. Drug Deliv. Sci. Technol. 2021, 66, 102802. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Astrogliosis. Cold Spring Harb. Perspect. Biol. 2015, 7, a020420. [Google Scholar] [CrossRef] [PubMed]
- Kuter, K.; Olech, Ł.; Głowacka, U.; Paleczna, M. Astrocyte Support Is Important for the Compensatory Potential of the Nigrostriatal System Neurons during Early Neurodegeneration. J. Neurochem. 2019, 148, 63–79. [Google Scholar] [CrossRef]
- Guo, T.; Zhang, D.; Zeng, Y.; Huang, T.Y.; Xu, H.; Zhao, Y. Molecular and Cellular Mechanisms Underlying the Pathogenesis of Alzheimer’s Disease. Mol. Neurodegener. 2020, 15, 40. [Google Scholar] [CrossRef]
- Maragakis, N.J.; Rothstein, J.D. Mechanisms of Disease: Astrocytes in Neurodegenerative Disease. Nat. Clin. Pract. Neurol. 2006, 2, 679–689. [Google Scholar] [CrossRef]
- Su, C.; Zhao, K.; Xia, H.; Xu, Y. Peripheral Inflammatory Biomarkers in Alzheimer’s Disease and Mild Cognitive Impairment: A Systematic Review and Meta-Analysis. Psychogeriatrics 2019, 19, 300–309. [Google Scholar] [CrossRef]
- Juźwik, C.A.; Drake, S.S.; Zhang, Y.; Paradis-Isler, N.; Sylvester, A.; Amar-Zifkin, A.; Douglas, C.; Morquette, B.; Moore, C.S.; Fournier, A.E. MicroRNA Dysregulation in Neurodegenerative Diseases: A Systematic Review. Prog. Neurobiol. 2019, 182, 101664. [Google Scholar] [CrossRef]
- Gasecka, A.; Siwik, D.; Gajewska, M.; Jaguszewski, M.J.; Mazurek, T.; Filipiak, K.J.; Postuła, M.; Eyileten, C. Early Biomarkers of Neurodegenerative and Neurovascular Disorders in Diabetes. J. Clin. Med. 2020, 9, 2807. [Google Scholar] [CrossRef]
- Jain, G.; Stuendl, A.; Rao, P.; Berulava, T.; Pena Centeno, T.; Kaurani, L.; Burkhardt, S.; Delalle, I.; Kornhuber, J.; Hüll, M.; et al. A Combined MiRNA–PiRNA Signature to Detect Alzheimer’s Disease. Transl. Psychiatry 2019, 9, 250. [Google Scholar] [CrossRef]
- Qin, C.; Xu, P.P.; Zhang, X.; Zhang, C.; Liu, C.B.; Yang, D.G.; Gao, F.; Yang, M.L.; Du, L.J.; Li, J.J. Pathological Significance of TRNA-Derived Small RNAs in Neurological Disorders. Neural Regen. Res. 2020, 15, 212–221. [Google Scholar] [CrossRef]
- Prehn, J.H.M.; Jirström, E. Angiogenin and TRNA Fragments in Parkinson’s Disease and Neurodegeneration. Acta Pharmacol. Sin. 2020, 41, 442–446. [Google Scholar] [CrossRef]
- Das, S.; Abdel-Mageed, A.B.; Adamidi, C.; Adelson, P.D.; Akat, K.M.; Alsop, E.; Ansel, K.M.; Arango, J.; Aronin, N.; Avsaroglu, S.K.; et al. The Extracellular RNA Communication Consortium: Establishing Foundational Knowledge and Technologies for Extracellular RNA Research. Cell 2019, 177, 231–242. [Google Scholar] [CrossRef]
- Szilágyi, M.; Pös, O.; Márton, É.; Buglyó, G.; Soltész, B.; Keserű, J.; Penyige, A.; Szemes, T.; Nagy, B. Circulating Cell-Free Nucleic Acids: Main Characteristics and Clinical Application. Int. J. Mol. Sci. 2020, 21, 6827. [Google Scholar] [CrossRef]
- Reed, E.R.; Latourelle, J.C.; Bockholt, J.H.; Bregu, J.; Smock, J.; Paulsen, J.S.; Myers, R.H. MicroRNAs in CSF as Prodromal Biomarkers for Huntington Disease in the PREDICT-HD Study. Neurology 2018, 90, E264–E272. [Google Scholar] [CrossRef]
- dos Santos, C.M.T.; Barreto-Sanz, M.A.; Coreira, B.R.S.; Bell, R.; Widnall, C.; Perez, L.T.; Berteau, C.; Schulte, C.; Scheller, D.; Berg, D.; et al. MiRNA-Based Signatures in Cerebrospinal Fluid as Potential Diagnostic Tools for Early Stage Parkinson’s Disease. Oncotarget 2018, 9, 17455–17465. [Google Scholar] [CrossRef]
- Griguolo, G.; Bottosso, M.; Vernaci, G.; Miglietta, F.; Dieci, M.V.; Guarneri, V. Gene-Expression Signatures to Inform Neoadjuvant Treatment Decision in HR+/HER2− Breast Cancer: Available Evidence and Clinical Implications. Cancer Treat. Rev. 2022, 102, 102323. [Google Scholar] [CrossRef]
- Willis, C.M.; Ménoret, A.; Jellison, E.R.; Nicaise, A.M.; Vella, A.T.; Crocker, S.J. A Refined Bead-Free Method to Identify Astrocytic Exosomes in Primary Glial Cultures and Blood Plasma. Front. Neurosci. 2017, 11, 335. [Google Scholar] [CrossRef]
- Mayne, K.; White, J.A.; McMurran, C.E.; Rivera, F.J.; de la Fuente, A.G. Aging and Neurodegenerative Disease: Is the Adaptive Immune System a Friend or Foe? Front. Aging Neurosci. 2020, 12, 305. [Google Scholar] [CrossRef]
- Wallensten, J.; Nager, A.; Åsberg, M.; Borg, K.; Beser, A.; Wilczek, A.; Mobarrez, F. Leakage of Astrocyte-Derived Extracellular Vesicles in Stress-Induced Exhaustion Disorder: A Cross-Sectional Study. Sci. Rep. 2021, 11, 2009. [Google Scholar] [CrossRef] [PubMed]
- Jovicic, A.; Gitler, A.D. Distinct Repertoires of MicroRNAs Present in Mouse Astrocytes Compared to Astrocytesecreted Exosomes. PLoS ONE 2017, 12, e0171418. [Google Scholar] [CrossRef] [PubMed]
- Lafourcade, C.; Ramírez, J.P.; Luarte, A.; Fernández, A.; Wyneken, U. MIRNAS in Astrocyte-Derived Exosomes as Possible Mediators of Neuronal Plasticity: Supplementary Issue: Brain Plasticity and Repair. J. Exp. Neurosci. 2016, 10, S39916. [Google Scholar] [CrossRef] [PubMed]
- Ogaki, A.; Ikegaya, Y.; Koyama, R. Extracellular Vesicles Taken up by Astrocytes. Int. J. Mol. Sci. 2021, 22, 10553. [Google Scholar] [CrossRef] [PubMed]
- Upadhya, R.; Zingg, W.; Shetty, S.; Shetty, A.K. Astrocyte-Derived Extracellular Vesicles: Neuroreparative Properties and Role in the Pathogenesis of Neurodegenerative Disorders. J. Control. Release 2020, 323, 225–239. [Google Scholar] [CrossRef]
- Yang, L.; Niu, F.; Yao, H.; Liao, K.; Chen, X.; Kook, Y.; Ma, R.; Hu, G.; Buch, S. Exosomal MiR-9 Released from HIV Tat Stimulated Astrocytes Mediates Microglial Migration. J. Neuroimmune Pharmacol. 2018, 13, 330–344. [Google Scholar] [CrossRef]
- Castellani, R.J.; Lee, H.G.; Zhu, X.; Nunomura, A.; Perry, G.; Smith, M.A. Neuropathology of Alzheimer Disease: Pathognomonic but Not Pathogenic. Acta Neuropathol. 2006, 111, 503–509. [Google Scholar] [CrossRef]
- Kishore, U. Neurodegenerative Diseases; BoD: Norderstedt, Germany, 2013. [Google Scholar]
- Carter, S.F.; Schöll, M.; Almkvist, O.; Wall, A.; Engler, H.; Långström, B.; Nordberg, A. Evidence for Astrocytosis in Prodromal Alzheimer Disease Provided by 11C-Deuterium-L-Deprenyl: A Multitracer PET Paradigm Combining 11C-Pittsburgh Compound B and 18F-FDG. J. Nucl. Med. 2012, 53, 37–46. [Google Scholar] [CrossRef]
- Wilson, H.; Dervenoulas, G.; Pagano, G.; Tyacke, R.J.; Polychronis, S.; Myers, J.; Gunn, R.N.; Rabiner, E.A.; Nutt, D. Imidazoline 2 Binding Sites Reflecting Astroglia Pathology in Parkinson’s Disease: An in Vivo 11C-BU99008 PET Study. Brain 2019, 142, 3116–3128. [Google Scholar] [CrossRef]
- Poutiainen, P.; Jaronen, M.; Quintana, F.J.; Brownell, A.; Harvey, K. Precision Medicine in Multiple Sclerosis: Future of PET Imaging of Inflammation and Reactive Astrocytes. Front. Mol. Neurosci. 2016, 9, 85. [Google Scholar] [CrossRef]
- Linnerbauer, M.; Wheeler, M.A.; Quintana, F.J. Astrocyte Crosstalk in CNS Inflammation. Neuron 2020, 108, 608. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Astrocyte Reactivity: Subtypes, States, and Functions in CNS Innate Immunity. Trends Immunol. 2020, 41, 758–770. [Google Scholar] [CrossRef]
- Santello, M.; Toni, N.; Volterra, A. Astrocyte Function from Information Processing to Cognition and Cognitive Impairment. Nat. Neurosci. 2019, 22, 154–166. [Google Scholar] [CrossRef]
- Bylicky, M.A.; Mueller, G.P.; Day, R.M. Mechanisms of Endogenous Neuroprotective Effects of Astrocytes in Brain Injury. Oxidative Med. Cell. Longev. 2018, 2018, 6501031. [Google Scholar] [CrossRef]
- Xie, K.; Qin, Q.; Long, Z.; Yang, Y.; Peng, C.; Xi, C.; Li, L.; Wu, Z.; Daria, V.; Zhao, Y.; et al. High-Throughput Metabolomics for Discovering Potential Biomarkers and Identifying Metabolic Mechanisms in Aging and Alzheimer’s Disease. Front. Cell Dev. Biol. 2021, 9, 335. [Google Scholar] [CrossRef]
- Patil, S.; Chan, C. Palmitic and Stearic Fatty Acids Induce Alzheimer-like Hyperphosphorylation of Tau in Primary Rat Cortical Neurons. Neurosci. Lett. 2005, 384, 288–293. [Google Scholar] [CrossRef]
- Ortiz-Rodriguez, A.; Acaz-Fonseca, E.; Boya, P.; Arevalo, M.A.; Garcia-Segura, L.M. Lipotoxic Effects of Palmitic Acid on Astrocytes Are Associated with Autophagy Impairment. Mol. Neurobiol. 2018, 56, 1665–1680. [Google Scholar] [CrossRef]
- González-Giraldo, Y.; Garcia-Segura, L.M.; Echeverria, V.; Barreto, G.E. Tibolone Preserves Mitochondrial Functionality and Cell Morphology in Astrocytic Cells Treated with Palmitic Acid. Mol. Neurobiol. 2017, 55, 4453–4462. [Google Scholar] [CrossRef]
- González-Giraldo, Y.; Forero, D.A.; Echeverria, V.; Garcia-Segura, L.M.; Barreto, G.E. Tibolone Attenuates Inflammatory Response by Palmitic Acid and Preserves Mitochondrial Membrane Potential in Astrocytic Cells through Estrogen Receptor Beta. Mol. Cell. Endocrinol. 2019, 486, 65–78. [Google Scholar] [CrossRef]
- Liu, L.; Martin, R.; Kohler, G.; Chan, C. Palmitate Induces Transcriptional Regulation of BACE1 and Presenilin by STAT3 in Neurons Mediated by Astrocytes. Exp. Neurol. 2013, 248, 482–490. [Google Scholar] [CrossRef][Green Version]
- Liu, L.; Martin, R.; Chan, C. Palmitate-Activated Astrocytes via Serine Palmitoyltransferase Increase BACE1 in Primary Neurons by Sphingomyelinases. Neurobiol. Aging 2013, 34, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Benzinger, T.L.S.; Blazey, T.; Jack, C.R.; Koeppe, R.A.; Su, Y.; Xiong, C.; Raichle, M.E.; Snyder, A.Z.; Ances, B.M.; Bateman, R.J.; et al. Regional Variability of Imaging Biomarkers in Autosomal Dominant Alzheimer’s Disease. Proc. Natl. Acad. Sci. USA 2013, 110, E4502–E4509. [Google Scholar] [CrossRef] [PubMed]
- Winocur, G.; Greenwood, C.E. Studies of the Effects of High Fat Diets on Cognitive Function in a Rat Model. Neurobiol. Aging 2005, 26, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Qin, W.; Pompl, P.N.; Xiang, Z.; Wang, J.; Zhao, Z.; Peng, Y.; Cambareri, G.; Rocher, A.; Mobbs, C.V.; et al. Diet-Induced Insulin Resistance Promotes Amyloidosis in a Transgenic Mouse Model of Alzheimer’s Disease. FASEB J. 2004, 18, 902–904. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.E.; Goez, D.; Last, D.; Naor, S.; Liraz Zaltsman, S.; Sharvit-Ginon, I.; Atrakchi-Baranes, D.; Shemesh, C.; Twitto-Greenberg, R.; Tsach, S.; et al. High-Fat Diet Protects the Blood–Brain Barrier in an Alzheimer’s Disease Mouse Model. Aging Cell 2018, 17, e12818. [Google Scholar] [CrossRef] [PubMed]
- Ghaddar, B.; Diotel, N. Zebrafish: A New Promise to Study the Impact of Metabolic Disorders on the Brain. Int. J. Mol. Sci. 2022, 23, 5372. [Google Scholar] [CrossRef]
- Booth, H.D.E.; Hirst, W.D.; Wade-Martins, R. The Role of Astrocyte Dysfunction in Parkinson’s Disease Pathogenesis. Trends Neurosci. 2017, 40, 358. [Google Scholar] [CrossRef]
- Deng, H.; Wang, P.; Jankovic, J. The Genetics of Parkinson Disease. Ageing Res. Rev. 2018, 42, 72–85. [Google Scholar] [CrossRef]
- Chamoli, M.; Chinta, S.J.; Andersen, J.K. An Inducible MAO-B Mouse Model of Parkinson’s Disease: A Tool towards Better Understanding Basic Disease Mechanisms and Developing Novel Therapeutics. J. Neural Transm. 2018, 125, 1651–1658. [Google Scholar] [CrossRef]
- Price, B.R.; Norris, C.M.; Sompol, P.; Wilcock, D.M. An Emerging Role of Astrocytes in Vascular Contributions to Cognitive Impairment and Dementia. J. Neurochem. 2018, 144, 644–650. [Google Scholar] [CrossRef]
- Arba, F.; Mair, G.; Phillips, S.; Sandercock, P.; Wardlaw, J.M. Improving Clinical Detection of Acute Lacunar Stroke: Analysis From the IST-3. Stroke 2020, 51, 1411. [Google Scholar] [CrossRef]
- Qu, P.; Cheng, K.; Gao, Q.; Li, Y.; Wang, M. Application Value of Serum Hcy, TLR4, and CRP in the Diagnosis of Cerebral Small Vessel Disease. Evidence-based Complement. Altern. Med. 2022, 2022, 4025965. [Google Scholar] [CrossRef]
- Ponath, G.; Park, C.; Pitt, D. The Role of Astrocytes in Multiple Sclerosis. Front. Immunol. 2018, 9, 217. [Google Scholar] [CrossRef]
- Mayo, L.; Trauger, S.A.; Blain, M.; Nadeau, M.; Patel, B.; Alvarez, J.I.; Mascanfroni, I.D.; Yeste, A.; Kivisäkk, P.; Kallas, K.; et al. Regulation of Astrocyte Activation by Glycolipids Drives Chronic CNS Inflammation. Nat. Med. 2014, 20, 1147–1156. [Google Scholar] [CrossRef]
- Sochocka, M.; Diniz, B.S.; Leszek, J. Inflammatory Response in the CNS: Friend or Foe? Mol. Neurobiol. 2017, 54, 8071–8089. [Google Scholar] [CrossRef]
- Ke, L.; Niu, F.; Hu, G.; Yang, L.; Dallon, B.; Villareal, D.; Buch, S. Morphine-Mediated Release OfmiR-138 in Astrocyte-Derived Extracellular Vesicles Promotes Microglial Activation. J Extracell. Vesicles 2020, 10, e12027. [Google Scholar] [CrossRef]
- Pozo, P.H.E.; del Espinosa, P.S.; Donadi, E.A.; Martinez, E.Z.; Salazar-Uribe, J.C.; Guerrero, M.A.; Moriguti, J.C.; Colcha, M.C.; Garcia, S.E.; Naranjo, R.; et al. Cognitive Decline in Adults Aged 65 and Older in Cumbayá, Quito, Ecuador: Prevalence and Risk Factors. Cureus 2018, 10, e3269. [Google Scholar] [CrossRef]
- Muscat, S.M.; Deems, N.P.; D’Angelo, H.; Kitt, M.M.; Grace, P.M.; Andersen, N.D.; Silverman, S.N.; Rice, K.C.; Watkins, L.R.; Maier, S.F.; et al. Postoperative Cognitive Dysfunction Is Made Persistent with Morphine Treatment in Aged Rats. Neurobiol. Aging 2021, 98, 214–224. [Google Scholar] [CrossRef]
- Balaban, D.; Miyawaki, E.K.; Bhattacharyya, S.; Torre, M. The Phenomenon of Clasmatodendrosis. Heliyon 2021, 7, e07605. [Google Scholar] [CrossRef]
- Fernandes, L.M.P.; Lopes, K.S.; Santana, L.N.S.; Fontes-Júnior, E.A.; Ribeiro, C.H.M.A.; Silva, M.C.F.; de Oliveira Paraense, R.S.; Crespo-López, M.E.; Gomes, A.R.Q.; Lima, R.R.; et al. Repeated Cycles of Binge-like Ethanol Intake in Adolescent Female Rats Induce Motor Function Impairment and Oxidative Damage in Motor Cortex and Liver, but Not in Blood. Oxidative Med. Cell. Longev. 2018, 2018, 3467531. [Google Scholar] [CrossRef]
- Chaudhuri, A.D.; Dastgheyb, R.M.; Yoo, S.-W.; Trout, A.; Talbot, C.C., Jr.; Hao, H.; Witwer, K.W.; Haughey, N.J. TNF Alpha and IL-1 Beta Modify the MiRNA Cargo of Astrocyte Shed Extracellular Vesicles to Regulate Neurotrophic Signaling in Neurons. Cell Death Dis. 2018, 9, 363. [Google Scholar] [CrossRef] [PubMed]
- Collado-Pérez, R.; García-Piqueres, J.; Jiménez-Hernaiz, M.; Argente, J.; Belsham, D.D.; Frago, L.M.; Chowen, J.A. Fatty Acids Modify the MicroRNA Content of Exosomes Released by Hypothalamic Astrocytes and the Response of POMC Neurons to These Exosomes. J. Endocr. Soc. 2021, 5 (Suppl. 1), A46. [Google Scholar] [CrossRef]
- Xu, L.; Cao, H.; Xie, Y.; Zhang, Y.; Du, M.; Xu, X.; Ye, R.; Liu, X. Exosome-Shuttled MiR-92b-3p from Ischemic Preconditioned Astrocytes Protects Neurons against Oxygen and Glucose Deprivation. Brain Res. 2019, 1717, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, F.; Montesinos, J.; Ureña-peralta, J.R.; Guerri, C. TLR4 Participates in the Transmission of Ethanol-Induced Neuroinflammation via Astrocyte-Derived Extracellular Vesicles. J. Neuroinflamm. 2019, 16, 136. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Yao, X.; Jiang, Q.; Yang, Y.; He, X.; Tian, W.; Zhao, K.; Zhang, H. Astrocyte-Derived Exosomes Enriched with MiR-873a-5p Inhibit Neuroinflammation via Microglia Phenotype Modulation after Traumatic Brain Injury. J. Neuroinflamm. 2020, 17, 89. [Google Scholar] [CrossRef]
- Du, L.; Jiang, Y.; Sun, Y. Astrocyte-Derived Exosomes Carry MicroRNA-17-5p to Protect Neonatal Rats from Hypoxic-Ischemic Brain Damage via Inhibiting BNIP-2 Expression. Neurotoxicology 2021, 83, 28–39. [Google Scholar] [CrossRef]
- Barbosa, M.; Gomes, C.; Vaz, A.R.; Brites, D. Upregulation of MiR-146a Attenuates ALS Mouse Cortical Astrocytes Reactivity and Decrease MiRNA-Inflammatory Associated Exosomal Cargo. Free Radic. Biol. Med. 2018, 120, S158. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, W.; Jing, L.; Wang, Y.; Li, M.; Hou, X.; Wang, J.; Peng, T.; Teng, J.; Jia, Y. Let-7f Regulates the Hypoxic Response in Cerebral Ischemia by Targeting NDRG3. Neurochem. Res. 2017, 42, 446–454. [Google Scholar] [CrossRef]
- Daneshafrooz, N.; Joghataei, M.T.; Mehdizadeh, M.; Alavi, A.; Barati, M.; Panahi, B.; Teimourian, S.; Zamani, B. Identification of Let-7f and MiR-338 as Plasma-Based Biomarkers for Sporadic Amyotrophic Lateral Sclerosis Using Meta-Analysis and Empirical Validation. Sci. Rep. 2022, 12, 1373. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Campdelacreu, J.; Vilas, D.; Ispierto, L.; Reñé, R.; Álvarez, R.; Armengol, M.P.; Borràs, F.E.; Beyer, K. Exploratory Study on MicroRNA Profiles from Plasma-Derived Extracellular Vesicles in Alzheimer’s Disease and Dementia with Lewy Bodies. Transl. Neurodegener. 2019, 8, 31. [Google Scholar] [CrossRef]
- Huang, H.-T.; Hsien, H.H.; Wu, H.-T.; Tsai, S.-F.; Huang, H.-Y.; Kuo, Y.-M.; Chen, P.-S.; Yang, C.-S.; Tzen, S.-F. High Fat Diet Induces Mitochondria Stress and Impairs Myelin Structure in Rat Hypothalamus. Glia 2017, 65, E103–E578. [Google Scholar] [CrossRef]
- Deng, Z.; Wei, Y.; Yao, Y.; Gao, S.; Wang, X. Let-7f Promotes the Differentiation of Neural Stem Cells in Rats. Am. J. Transl. Res. 2020, 12, 5752–5761. [Google Scholar]
- Buonfiglioli, A.; Efe, I.E.; Guneykaya, D.; Ivanov, A.; Huang, Y.; Orlowski, E.; Krüger, C.; Deisz, R.A.; Markovic, D.; Flüh, C.; et al. Let-7 MicroRNAs Regulate Microglial Function and Suppress Glioma Growth through Toll-Like Receptor 7. Cell Rep. 2019, 29, 3460–3471.e7. [Google Scholar] [CrossRef]
- Yan, S.; Han, X.; Xue, H.; Zhang, P.; Guo, X.; Li, T.; Guo, X.; Yuan, G.; Deng, L.; Li, G. Let-7f Inhibits Glioma Cell Proliferation, Migration, and Invasion by Targeting Periostin. J. Cell. Biochem. 2015, 116, 1680–1692. [Google Scholar] [CrossRef]
- Li, K.; Wang, Z.-Q.; Zhang, J.-L.; Lv, P.-Y. MicroRNA Let-7f Protects against H2O2-Induced Oxidative Damage in Neuroblastoma Cells by Targeting AKT-2. Arch. Med. Sci. 2020, 16, 1–10. [Google Scholar] [CrossRef]
- Shibahara, Y.; Miki, Y.; Onodera, Y.; Hata, S.; Chan, M.S.M.; Yiu, C.C.P.; Loo, T.Y.; Nakamura, Y.; Akahira, J.I.; Ishida, T.; et al. Aromatase Inhibitor Treatment of Breast Cancer Cells Increases the Expression of Let-7f, a MicroRNA Targeting CYP19A1. J. Pathol. 2012, 227, 357–366. [Google Scholar] [CrossRef]
- Zhao, Q.-C.; Xu, Z.-W.; Peng, Q.-M.; Zhou, J.-H.; Li, Z.-Y. Enhancement of MiR-16-5p on Spinal Cord Injury-Induced Neuron Apoptosis and Inflammatory Response through Inactivating ERK1/2 Pathway. J. Neurosurg. Sci. 2020. [Google Scholar] [CrossRef]
- Sun, Y.; Xiong, Y.; Yan, C.; Chen, L.; Chen, D.; Mi, B.; Liu, G. Downregulation of MicroRNA-16-5p Accelerates Fracture Healing by Promoting Proliferation and Inhibiting Apoptosis of Osteoblasts in Patients with Traumatic Brain Injury. Am. J. Transl. Res. 2019, 11, 4746–4760. [Google Scholar]
- Joilin, G.; Gray, E.; Thompson, A.G.; Bobeva, Y.; Talbot, K.; Weishaupt, J.; Ludolph, A.; Malaspina, A.; Leigh, P.N.; Newbury, S.F.; et al. Identification of a Potential Non-Coding RNA Biomarker Signature for Amyotrophic Lateral Sclerosis. Brain Commun. 2020, 2, fcaa053. [Google Scholar] [CrossRef]
- Wang, N.; He, L.; Yang, Y.; Li, S.; Chen, Y.; Tian, Z.; Ji, Y.; Wang, Y.; Pang, M.; Wang, Y.; et al. Integrated Analysis of Competing Endogenous RNA (CeRNA) Networks in Subacute Stage of Spinal Cord Injury. Gene 2020, 726, 144171. [Google Scholar] [CrossRef]
- Tian, F.; Yang, J.; Xia, R. Exosomes Secreted from CircZFHX3-Modified Mesenchymal Stem Cells Repaired Spinal Cord Injury Through Mir-16-5p/IGF-1 in Mice. Neurochem. Res. 2022, 47, 2076–2089. [Google Scholar] [CrossRef] [PubMed]
- You, S.; He, X.; Wang, M.; Mao, L.; Zhang, L. Tanshinone IIA Suppresses Glioma Cell Proliferation, Migration and Invasion Both In Vitro and In Vivo Partially through MiR-16-5p/Talin-1 (TLN1) Axis. Cancer Manag. Res. 2020, 12, 11309. [Google Scholar] [CrossRef] [PubMed]
- Abyadeh, M.; Tofigh, N.; Hosseinian, S.; Hasan, M.; Amirkhani, A.; Fitzhenry, M.J.; Gupta, V.; Chitranshi, N.; Salekdeh, G.H.; Haynes, P.A.; et al. Key Genes and Biochemical Networks in Various Brain Regions Affected in Alzheimer’s Disease. Cells 2022, 11, 987. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Dixon, B.J.; Doycheva, D.M.; Li, B.; Zhang, Y.; Hu, Q.; He, Y.; Guo, Z.; Nowrangi, D.; Flores, J.; et al. IRE1α Inhibition Decreased TXNIP/NLRP3 Inflammasome Activation through MiR-17-5p after Neonatal Hypoxic-Ischemic Brain Injury in Rats. J. Neuroinflamm. 2018, 15, 32. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.; Jiang, M.; Li, H. Functional Requirement of Dicer1 and MiR-17-5p in Reactive Astrocyte Proliferation after Spinal Cord Injury in the Mouse. Glia 2014, 62, 2044–2060. [Google Scholar] [CrossRef]
- Sajad, M.; Ahmed, M.M.; Thakur, S.C. An Integrated Bioinformatics Strategy to Elucidate the Function of Hub Genes Linked to Alzheimer’s Disease. Gene Rep. 2022, 26, 101534. [Google Scholar] [CrossRef]
- Estfanous, S.; Daily, K.P.; Eltobgy, M.; Deems, N.P.; Anne, M.N.K.; Krause, K.; Badr, A.; Hamilton, K.; Carafice, C.; Hegazi, A.; et al. Elevated Expression of MiR-17 in Microglia of Alzheimer’s Disease Patients Abrogates Autophagy-Mediated Amyloid-β Degradation. Front. Immunol. 2021, 12, 2839. [Google Scholar] [CrossRef]
- Hébert, S.S.; Horré, K.; Nicolaï, L.; Bergmans, B.; Papadopoulou, A.S.; Delacourte, A.; De Strooper, B. MicroRNA Regulation of Alzheimer’s Amyloid Precursor Protein Expression. Neurobiol. Dis. 2009, 33, 422–428. [Google Scholar] [CrossRef]
- Chen, B.; Yang, Y.; Wu, J.; Song, J.; Lu, J. MicroRNA-17-5p Downregulation Inhibits Autophagy and Myocardial Remodelling after Myocardial Infarction by Targeting STAT3. Autoimmunity 2021, 55, 43–51. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, M.; Guo, Q.; Lou, J.; Wang, L. Non-Small Cell Lung Cancer Cell–Derived Exosomal MiR-17-5p Promotes Osteoclast Differentiation by Targeting PTEN. Exp. Cell Res. 2021, 408, 112834. [Google Scholar] [CrossRef]
- Qin, X.; Zhu, B.; Jiang, T.; Tan, J.; Wu, Z.; Yuan, Z.; Zheng, L.; Zhao, J. MiR-17-5p Regulates Heterotopic Ossification by Targeting ANKH in Ankylosing Spondylitis. Mol. Ther. Nucleic Acids 2019, 18, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, X.; Song, X.; Tang, Y. Expression of MiR-17-5p in Gastrointestinal Stromal Tumor Tissues and Its Effect on Proliferation and Apoptosis of GIST882 Cells. Chin. J. Cancer Biother. 2022, 28, 721–727. [Google Scholar] [CrossRef]
- Deng, X.H.; Zhong, Y.; Gu, L.Z.; Shen, W.; Guo, J. MiR-21 Involve in ERK-Mediated Upregulation of MMP9 in the Rat Hippocampus Following Cerebral Ischemia. Brain Res. Bull. 2013, 94, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ren, Y.; Moore, L.; Mei, M.; You, Y.; Xu, P.; Wang, B.; Wang, G.; Jia, Z.; Pu, P.; et al. Downregulation of MiR-21 Inhibits EGFR Pathway and Suppresses the Growth of Human Glioblastoma Cells Independent of PTEN Status. Lab. Investig. 2010, 90, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Buller, B.; Liu, X.; Wang, X.; Zhang, R.L.; Zhang, L.; Hozeska-Solgot, A.; Chopp, M.; Zhang, Z.G. MicroRNA-21 Protects Neurons from Ischemic Death. FEBS J. 2010, 277, 4299–4307. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Chen, Z.; Du, H.; Liu, R.; Wang, W.; Li, H.; Ning, B. Silencing MiR-21 Induces Polarization of Astrocytes to the A2 Phenotype and Improves the Formation of Synapses by Targeting Glypican 6 via the Signal Transducer and Activator of Transcription-3 Pathway after Acute Ischemic Spinal Cord Injury. FASEB J. 2019, 33, 10859–10871. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dong, L.Y.; Li, Y.J.; Hong, Z.; Wei, W.S. MiR-21 Represses FasL in Microglia and Protects against Microglia-Mediated Neuronal Cell Death Following Hypoxia/Ischemia. Glia 2012, 60, 1888–1895. [Google Scholar] [CrossRef]
- Feng, M.G.; Liu, C.F.; Chen, L.; Feng, W.B.; Liu, M.; Hai, H.; Lu, J.M. MiR-21 Attenuates Apoptosis-Triggered by Amyloid-β via Modulating PDCD4/PI3K/AKT/GSK-3β Pathway in SH-SY5Y Cells. Biomed. Pharmacother. 2018, 101, 1003–1007. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, Y.; Luo, F.; Xu, W.; Wang, B.; Pang, Y.; Zhou, J.; Wang, X.; Liu, Q. Angiogenesis, Mediated by MiR-21, Is Involved Arsenite-Induced Carcinogenesis. Toxicol. Lett. 2013, 223, 35–41. [Google Scholar] [CrossRef]
- Liguori, M.; Nuzziello, N.; Introna, A.; Consiglio, A.; Licciulli, F.; D’Errico, E.; Scarafino, A.; Distaso, E.; Simone, I.L. Dysregulation of MicroRNAs and Target Genes Networks in Peripheral Blood of Patients with Sporadic Amyotrophic Lateral Sclerosis. Front. Mol. Neurosci. 2018, 11, 288. [Google Scholar] [CrossRef]
- Janik, P.; Fitzgerald, J.C.; Rai, S.N.; Huo, J.; Chen, M.; Peng, L.; Gong, P.; Zheng, X.; Sun, T.; Zhang, X. Baicalein Induces Mitochondrial Autophagy to Prevent Parkinson’s Disease in Rats via MiR-30b and the SIRT1/AMPK/MTOR Pathway. Front. Neurol. 2022, 12, 646817. [Google Scholar] [CrossRef]
- Mazzeo, A.; Lopatina, T.; Gai, C.; Trento, M.; Porta, M.; Beltramo, E. Functional Analysis of MiR-21-3p, MiR-30b-5p and MiR-150-5p Shuttled by Extracellular Vesicles from Diabetic Subjects Reveals Their Association with Diabetic Retinopathy. Exp. Eye Res. 2019, 184, 56–63. [Google Scholar] [CrossRef]
- Guo, H.; Pu, M.; Tai, Y.; Chen, Y.; Lu, H.; Qiao, J.; Wang, G.; Chen, J.; Qi, X.; Huang, R.; et al. Nuclear MiR-30b-5p Suppresses TFEB-Mediated Lysosomal Biogenesis and Autophagy. Cell Death Differ. 2020, 28, 320–336. [Google Scholar] [CrossRef]
- Shin, D.; Howng, S.Y.B.; Ptáček, L.J.; Fu, Y.H. MiR-32 and Its Target SLC45A3 Regulate the Lipid Metabolism of Oligodendrocytes and Myelin. Neuroscience 2012, 213, 29–37. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; An, W.; Chen, C.; Wang, W.; Zhu, C.; Chen, F.; Chen, H.; Zheng, W.; Gong, J. MiR-32 Inhibits Proliferation and Metastasis by Targeting EZH2 in Glioma. Technol. Cancer Res. Treat. 2019, 18, 1533033819854132. [Google Scholar] [CrossRef]
- Jin, Y.; Cheng, H.; Cao, J.; Shen, W. MicroRNA 32 Promotes Cell Proliferation, Migration, and Suppresses Apoptosis in Colon Cancer Cells by Targeting OTU Domain Containing 3. J. Cell. Biochem. 2019, 120, 18629–18639. [Google Scholar] [CrossRef] [PubMed]
- Scaravilli, M.; Koivukoski, S.; Gillen, A.; Bouazza, A.; Ruusuvuori, P.; Visakorpi, T.; Latonen, L. MiR-32 Promotes MYC-Driven Prostate Cancer. Oncogenesis 2022, 11, 11. [Google Scholar] [CrossRef]
- Chen, Y.C.; Hsu, P.Y.; Su, M.C.; Chen, T.W.; Hsiao, C.C.; Chin, C.H.; Liou, C.W.; Wang, P.W.; Wang, T.Y.; Lin, Y.Y.; et al. Microrna Sequencing Analysis in Obstructive Sleep Apnea and Depression: Anti-Oxidant and Maoa-Inhibiting Effects of Mir-15b-5p and Mir-92b-3p through Targeting Ptgs1-Nf-Κb-Sp1 Signaling. Antioxidants 2021, 10, 1854. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Z.; Jiang, C.; Jiang, X.; Zhang, J. MiR-92b-3p Promotes Neurite Growth and Functional Recovery via the PTEN/AKT Pathway in Acute Spinal Cord Injury. J. Cell. Physiol. 2019, 234, 23043–23052. [Google Scholar] [CrossRef]
- Hao, X.; Ma, C.; Chen, S.; Dang, J.; Cheng, X.; Zhu, D. Reverse the down Regulation of MiR-92b-3p by Hypoxia Can Suppress the Proliferation of Pulmonary Artery Smooth Muscle Cells by Targeting USP28. Biochem. Biophys. Res. Commun. 2018, 503, 3064–3077. [Google Scholar] [CrossRef]
- Paul, S.; Vázquez, L.A.B.; Uribe, S.P.; Reyes-Pérez, P.R.; Sharma, A. Current Status of MicroRNA-Based Therapeutic Approaches in Neurodegenerative Disorders. Cells 2020, 9, 1698. [Google Scholar] [CrossRef] [PubMed]
- Magri, F.; Vanoli, F.; Corti, S. MiRNA in Spinal Muscular Atrophy Pathogenesis and Therapy. J. Cell. Mol. Med. 2018, 22, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Kong, N.; Lu, X.; Li, B. Downregulation of MicroRNA-100 Protects Apoptosis and Promotes Neuronal Growth in Retinal Ganglion Cells. BMC Mol. Biol. 2014, 15, 25. [Google Scholar] [CrossRef] [PubMed]
- Wallach, T.; Mossmann, Z.J.; Szczepek, M.; Wetzel, M.; Machado, R.; Raden, M.; Miladi, M.; Kleinau, G.; Krüger, C.; Dembny, P.; et al. MicroRNA-100-5p and MicroRNA-298-5p Released from Apoptotic Cortical Neurons Are Endogenous Toll-like Receptor 7/8 Ligands That Contribute to Neurodegeneration. Mol. Neurodegener. 2021, 16, 80. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Fu, N.S.; Xing, Z.M. MiR-100 Suppresses Inflammatory Activation of Microglia and Neuronal Apoptosis Following Spinal Cord Injury via TLR4/NF-ΚB Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8713–8720. [Google Scholar] [CrossRef]
- Wang, A.P.; Li, X.H.; Gong, S.X.; Li, W.Q.; Hu, C.P.; Zhang, Z.; Li, Y.J. MIR-100 Suppresses MTOR Signaling in Hypoxia-Induced Pulmonary Hypertension in Rats. Eur. J. Pharmacol. 2015, 765, 565–573. [Google Scholar] [CrossRef]
- Cha, D.J.; Franklin, J.L.; Dou, Y.; Liu, Q.; Higginbotham, J.N.; Beckler, M.D.; Weaver, A.M.; Vickers, K.; Prasad, N.; Levy, S.; et al. KRAS-Dependent Sorting of MiRNA to Exosomes. eLife 2015, 4, e07197. [Google Scholar] [CrossRef]
- Sun, C.; Liu, J.; Duan, F.; Cong, L.; Qi, X. The Role of the MicroRNA Regulatory Network in Alzheimer’s Disease: A Bioinformatics Analysis. Arch. Med. Sci. 2022, 18, 206–222. [Google Scholar] [CrossRef]
- Kit, O.I.; Pushkin, A.A.; Alliluyev, I.A.; Timoshkina, N.N.; Gvaldin, D.Y.; Rostorguev, E.E.; Kuznetsova, N.S. Differential Expression of MicroRNAs Targeting Genes Associated with the Development of High-Grade Gliomas. Egypt. J. Med. Hum. Genet. 2022, 23, 31. [Google Scholar] [CrossRef]
- Herrero-Aguayo, V.; Sáez-Martínez, P.; Jiménez-Vacas, J.M.; Moreno-Montilla, M.T.; Montero-Hidalgo, A.J.; Pérez-Gómez, J.M.; López-Canovas, J.L.; Porcel-Pastrana, F.; Carrasco-Valiente, J.; Anglada, F.J.; et al. Dysregulation of the MiRNome Unveils a Crosstalk between Obesity and Prostate Cancer: MiR-107 Asa Personalized Diagnostic and Therapeutic Tool. Mol. Ther. Nucleic Acids 2022, 27, 1164–1178. [Google Scholar] [CrossRef]
- Hu, G.; Liao, K.; Yang, L.; Pendyala, G.; Kook, Y.; Fox, H.S.; Buch, S. Tat-Mediated Induction of MiRs-34a &-138 Promotes Astrocytic Activation via Downregulation of SIRT1: Implications for Aging in HAND. J. Neuroimmune Pharmacol. 2017, 12, 420–432. [Google Scholar] [CrossRef]
- Madsen, P.M.; Motti, D.; Karmally, S.; Szymkowski, D.E.; Lambertsen, K.L.; Bethea, J.R.; Brambilla, R. Oligodendroglial TNFR2 Mediates Membrane TNF-Dependent Repair in Experimental Autoimmune Encephalomyelitis by Promoting Oligodendrocyte Differentiation and Remyelination. J. Neurosci. 2016, 36, 5128–5143. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, H.; Li, Y.; Mao, R.; Yang, H.; Zhang, Y.; Zhang, Y.; Guo, P.; Zhan, D.; Zhang, T. Circular RNA SAMD4A Controls Adipogenesis in Obesity through the MiR-138-5p/EZH2 Axis. Theranostics 2020, 10, 4705. [Google Scholar] [CrossRef]
- Devier, D.J.; Lovera, J.F.; Lukiw, W.J. Increase in NF-ΚB-Sensitive MiRNA-146a and MiRNA-155 in Multiple Sclerosis (MS) and pro-Inflammatory Neurodegeneration. Front. Mol. Neurosci. 2015, 8, 1–5. [Google Scholar] [CrossRef]
- Liu, X.S.; Chopp, M.; Pan, W.L.; Wang, X.L.; Fan, B.Y.; Zhang, Y.; Kassis, H.; Zhang, R.L.; Zhang, X.M.; Zhang, Z.G. MicroRNA-146a Promotes Oligodendrogenesis in Stroke. Mol. Neurobiol. 2017, 54, 227–237. [Google Scholar] [CrossRef]
- Gomes, C.; Cunha, C.; Nascimento, F.; Ribeiro, J.A.; Vaz, A.R.; Brites, D. Cortical Neurotoxic Astrocytes with Early ALS Pathology and MiR-146a Deficit Replicate Gliosis Markers of Symptomatic SOD1G93A Mouse Model. Mol. Neurobiol. 2019, 56, 2137–2158. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, X.; Wu, X.; Chen, M.; Yu, W. MiR-146a Alleviates Lung Injury Caused by RSV Infection in Young Rats by Targeting TRAF-6 and Regulating JNK/ERKMAPK Signaling Pathways. Sci. Rep. 2022, 12, 3481. [Google Scholar] [CrossRef]
- Lopez-Ramirez, M.A.; Wu, D.; Pryce, G.; Simpson, J.E.; Reijerkerk, A.; King-Robson, J.; Kay, O.; De Vries, H.E.; Hirst, M.C.; Sharrack, B.; et al. MicroRNA-155 Negatively Affects Blood-Brain Barrier Function during Neuroinflammation. FASEB J. 2014, 28, 2551–2565. [Google Scholar] [CrossRef]
- Venkatesha, S.H.; Dudics, S.; Song, Y.; Mahurkar, A.; Moudgil, K.D. The miRNA Expression Profile of Experimental Autoimmune Encephalomyelitis Reveals Novel Potential Disease Biomarkers. Int. J. Mol. Sci. 2018, 19, 3990. [Google Scholar] [CrossRef]
- Li, X.; Wang, S.; Mu, W.; Barry, J.; Han, A.; Carpenter, R.L.; Jiang, B.-H.; Peiper, S.C.; Mahoney, M.G.; Aplin, A.E.; et al. Reactive Oxygen Species Reprogram Macrophages to Suppress Antitumor Immune Response through the Exosomal MiR-155-5p/PD-L1 Pathway. J. Exp. Clin. Cancer Res. 2021, 41, 41. [Google Scholar] [CrossRef]
- Fei, M.; Li, Z.; Cao, Y.; Jiang, C.; Lin, H.; Chen, Z. MicroRNA-182 Improves Spinal Cord Injury in Mice by Modulating Apoptosis and the Inflammatory Response via IKKβ/NF-ΚB. Lab. Investig. 2021, 101, 1238–1253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Tian, C.; Wu, J.; Zhang, Y.; Wang, J.; Kong, Q.; Mu, L.; Sun, B.; Ai, T.; Wang, Y.; et al. MicroRNA-182 Exacerbates Blood-Brain Barrier (BBB) Disruption by Downregulating the MTOR/FOXO1 Pathway in Cerebral Ischemia. FASEB J. 2020, 34, 13762–13775. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.M.; Lu, G.; Su, X.W.; Lyu, H.; Poon, W.S. MicroRNA-182 Regulates Neurite Outgrowth Involving the PTEN/AKT Pathway. Front. Cell. Neurosci. 2017, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, L.; Liu, Z.; Huang, T.; Wang, Y.; Ma, Y.; Fang, X.; He, Y.; Zhou, Y.; Huo, L.; et al. Research Paper MiRNA-182 Regulated MTSS1 Inhibits Proliferation and Invasion in Glioma Cells. J. Cancer 2020, 11, 5840–5851. [Google Scholar] [CrossRef]
- Zhou, F.; Fu, W.D.; Chen, L. MiRNA-182 Regulates the Cardiomyocyte Apoptosis in Heart Failure. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4917–4923. [Google Scholar] [CrossRef]
- Li, Y.; Luo, Y.; Li, B.; Niu, L.; Liu, J.; Duan, X. MiRNA-182/Deptor/MTOR Axis Regulates Autophagy to Reduce Intestinal Ischaemia/Reperfusion Injury. J. Cell. Mol. Med. 2020, 24, 7873–7883. [Google Scholar] [CrossRef]
- Nayak, B.; Khan, N.; Garg, H.; Rustagi, Y.; Singh, P.; Seth, A.; Dinda, A.K.; Kaushal, S. Role of MiRNA-182 and MiRNA-187 as Potential Biomarkers in Prostate Cancer and Its Correlation with the Staging of Prostate Cancer. Int. Braz. J. Urol. 2020, 46, 614–623. [Google Scholar] [CrossRef]
- Fu, J.; Peng, L.; Tao, T.; Chen, Y.; Li, Z.; Li, J. Regulatory Roles of the MiR-200 Family in Neurodegenerative Diseases. Biomed. Pharmacother. 2019, 119, 109409. [Google Scholar] [CrossRef]
- Higaki, S.; Muramatsu, M.; Matsuda, A.; Matsumoto, K.; Satoh, J.-I.; Michikawa, M.; Niida, S. Defensive Effect of MicroRNA-200b/c against Amyloid-Beta Peptide-Induced Toxicity in Alzheimer’s Disease Models. PLoS ONE 2018, 13, e0196929. [Google Scholar] [CrossRef]
- Yu, J.; Qin, M.; Li, J.; Cui, S. LncRNA SNHG4 Sponges MiR-200b to Inhibit Cell Apoptosis in Diabetic Retinopathy. Arch. Physiol. Biochem. 2021, 6, 1–6. [Google Scholar] [CrossRef]
- Huang, J.; Liang, X.; Wang, J.; Kong, Y.; Zhang, Z.; Ding, Z.; Song, Z.; Guo, Q.; Zou, W. MiR-873a-5p Targets A20 to Facilitate Morphine Tolerance in Mice. Front. Neurosci. 2019, 13, 347. [Google Scholar] [CrossRef]
- Fan, Y.; Li, J.; Yang, Q.; Gong, C.; Gao, H.; Mao, Z.; Yuan, X.; Zhu, S.; Xue, Z. Dysregulated Long Non-Coding RNAs in Parkinson’s Disease Contribute to the Apoptosis of Human Neuroblastoma Cells. Front. Neurosci. 2019, 13, 1320. [Google Scholar] [CrossRef]
- Lin, Y.H.; Guo, L.; Yan, F.; Dou, Z.Q.; Yu, Q.; Chen, G. Long Non-Coding RNA HOTAIRM1 Promotes Proliferation and Inhibits Apoptosis of Glioma Cells by Regulating the MiR-873-5p/ZEB2 Axis. Chin. Med. J. 2020, 133, 174–182. [Google Scholar] [CrossRef]
- Pitkänen, A.; Paananen, T.; Kyyriäinen, J.; Das Gupta, S.; Heiskanen, M.; Vuokila, N.; Bañuelos-Cabrera, I.; Lapinlampi, N.; Kajevu, N.; Andrade, P.; et al. Biomarkers for Posttraumatic Epilepsy. Epilepsy Behav. 2021, 121, 107080. [Google Scholar] [CrossRef]
- Califf, R.M. Minireview Biomarker Definitions and Their Applications. Exp. Biol. Med. 2018, 243, 213–221. [Google Scholar] [CrossRef]
- Majkić-Singh, N. What Is a Biomarker? From Its Discovery to Clinical Application. J. Med. Biochem. 2011, 30, 186–192. [Google Scholar] [CrossRef]
- Younas, N.; Flores, L.C.F.; Hopfner, F.; Höglinger, G.U.; Zerr, I. A New Paradigm for Diagnosis of Neurodegenerative Diseases: Peripheral Exosomes of Brain Origin. Transl. Neurodegener. 2022, 11, 28. [Google Scholar] [CrossRef]
- Brownlee, W.J. Misdiagnosis of Multiple Sclerosis. Neurology 2019, 92, 15–16. [Google Scholar] [CrossRef]
- Happich, M.; Kirson, N.Y.; Desai, U.; King, S.; Birnbaum, H.G.; Reed, C.; Belger, M.; Lenox-Smith, A.; Price, D. Excess Costs Associated with Possible Misdiagnosis of Alzheimer’s Disease among Patients with Vascular Dementia in a UK CPRD Population. J. Alzheimer’s Dis. 2016, 53, 171–183. [Google Scholar] [CrossRef]
- Tang, W.; Hu, Z.; Muallem, H.; Gulley, M.L. Clinical Implementation of RNA Signatures for Pharmacogenomic Decision-Making. Pharmgenom. Pers. Med. 2011, 2011, 95–107. [Google Scholar] [CrossRef]
- Qian, Y.; Daza, J.; Itzel, T.; Betge, J.; Zhan, T.; Marm, F.; Teufel, A. Prognostic Cancer Gene Expression Signatures: Current Status and Challenges. Cells 2021, 10, 648. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.J.; Wong, B.Y.X.; Vaidyanathan, R.; Sreejith, S.; Chia, S.Y.; Kandiah, N.; Ng, A.S.L.; Zeng, L. Altered Cerebrospinal Fluid Exosomal MicroRNA Levels in Young-Onset Alzheimer’s Disease and Frontotemporal Dementia. J. Alzheimer’s Dis. Rep. 2021, 5, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hong, G.; Lin, M.; Shi, Y.; Wang, L.; Jiang, F.; Zhang, F.; Wang, Y.; Guo, Z. Identification of Molecular Alterations in Leukocytes from Gene Expression Profiles of Peripheral Whole Blood of Alzheimer’s Disease. Sci. Rep. 2017, 7, 14027. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, T.; Zhang, Z.; Yang, G.Y. The Function of Astrocyte Mediated Extracellular Vesicles in Central Nervous System Diseases. Front. Cell Dev. Biol. 2020, 8, 568889. [Google Scholar] [CrossRef] [PubMed]
- Chim, S.S.C.; Shing, T.K.F.; Hung, E.C.W.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.K.; Lo, Y.M.D. Detection and Characterization of Placental MicroRNAs in Maternal Plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating MicroRNAs as Stable Blood-Based Markers for Cancer Detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef]
- Vidal, M. Exosomes: Revisiting Their Role as “Garbage Bags". Traffic 2019, 20, 815–828. [Google Scholar] [CrossRef]
- Meldolesi, J. Extracellular Vesicles (Exosomes and Ectosomes) Play Key Roles in the Pathology of Brain Diseases. Mol. Biomed. 2021, 2, 18. [Google Scholar] [CrossRef]
- Mussbacher, M.; Pirabe, A.; Brunnthaler, L.; Schrottmaier, W.C.; Assinger, A. Horizontal MicroRNA Transfer by Platelets—Evidence and Implications. Front. Physiol. 2021, 12, 678362. [Google Scholar] [CrossRef]
- Mao, S.; Sun, Q.; Xiao, H.; Zhang, C.; Li, L. Secreted MiR-34a in Astrocytic Shedding Vesicles Enhanced the Vulnerability of Dopaminergic Neurons to Neurotoxins by Targeting Bcl-2. Protein Cell 2015, 6, 529–540. [Google Scholar] [CrossRef]
- Battistelli, M.; Falcieri, E. Apoptotic Bodies: Particular Extracellular Vesicles Involved in Intercellular Communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef]
- Valle-Tamayo, N.; Pérez-González, R.; Chiva-Blanch, G.; Belbin, O.; Serrano-Requena, S.; Sirisi, S.; González, A.C.; Giró, O.; Sánchez-Aced, É.; Dols-Icardo, O.; et al. Enrichment of Astrocyte-Derived Extracellular Vesicles from Human Plasma. J. Vis. Exp. 2022, 2022, e64107. [Google Scholar] [CrossRef]
- Dickens, A.M.; Tovar-Y-Romo, L.B.; Yoo, S.-W.W.; Trout, A.L.; Bae, M.; Kanmogne, M.; Megra, B.; Williams, D.W.; Witwer, K.W.; Gacias, M.; et al. Astrocyte-Shed Extracellular Vesicles Regulate the Peripheral Leukocyte Response to Inflammatory Brain Lesions. Sci. Signal. 2017, 10, eaai7696. [Google Scholar] [CrossRef]
- García-Romero, N.; Carrión-Navarro, J.; Esteban-Rubio, S.; Lázaro-Ibáñez, E.; Peris-Celda, M.; Alonso, M.M.; Guzmán-De-Villoria, J.; Fernández-Carballal, C.; de Mendivil, A.O.; García-Duque, S.; et al. DNA Sequences within Glioma-Derived Extracellular Vesicles Can Cross the Intact Blood-Brain Barrier and Be Detected in Peripheral Blood of Patients. Oncotarget 2017, 8, 1416. [Google Scholar] [CrossRef]
- Chung, N.S.; Wasan, K.M. Potential Role of the Low-Density Lipoprotein Receptor Family as Mediators of Cellular Drug Uptake. Adv. Drug Deliv. Rev. 2004, 56, 1315–1334. [Google Scholar] [CrossRef]
- Wei, Z.; Batagov, A.O.; Schinelli, S.; Wang, J.; Wang, Y.; El Fatimy, R.; Rabinovsky, R.; Balaj, L.; Chen, C.C.; Hochberg, F.; et al. Coding and Noncoding Landscape of Extracellular RNA Released by Human Glioma Stem Cells. Nat. Commun. 2017, 8, 1145. [Google Scholar] [CrossRef]
- Bayraktar, R.; Van Roosbroeck, K.; Calin, G.A. Cell-to-cell Communication: MicroRNAs as Hormones. Mol. Oncol. 2017, 11, 1673. [Google Scholar] [CrossRef]
- Dehouck, B.; Fenart, L.; Dehouck, M.P.; Pierce, A.; Torpier, G.; Cecchelli, R. A New Function for the LDL Receptor: Transcytosis of LDL across the Blood–Brain Barrier. J. Cell Biol. 1997, 138, 877–889. [Google Scholar] [CrossRef]
- Rhea, E.M.; Banks, W.A. Interactions of Lipids, Lipoproteins, and Apolipoproteins with the Blood-Brain Barrier. Pharm. Res. 2021, 38, 1469–1475. [Google Scholar] [CrossRef]
- You, Y.; Muraoka, S.; Jedrychowski, M.P.; Hu, J.; McQuade, A.K.; Young-Pearse, T.; Aslebagh, R.; Shaffer, S.A.; Gygi, S.P.; Blurton-Jones, M.; et al. Human Neural Cell Type-Specific Extracellular Vesicle Proteome Defines Disease-Related Molecules Associated with Activated Astrocytes in Alzheimer’s Disease Brain. J. Extracell. Vesicles 2022, 11, e12183. [Google Scholar] [CrossRef]
- Gosselin, R.-D.; Meylan, P.; Decosterd, I. Extracellular Microvesicles from Astrocytes Contain Functional Glutamate Transporters: Regulation by Protein Kinase C and Cell Activation. Front. Cell. Neurosci. 2013, 7, 251. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Gerstberger, S.; Hafner, M.; Tuschl, T. A Census of Human RNA-Binding Proteins. Nat. Rev. Genet. 2014, 15, 829–845. [Google Scholar] [CrossRef] [PubMed]
- Galvanin, A.; Dostert, G.; Ayadi, L.; Marchand, V.; Motorin, Y. Diversity and Heterogeneity of Extracellular RNA in Human Plasma. Biochimie 2019, 164, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Shurtleff, M.J.; Yao, J.; Qin, Y.; Nottingham, R.M.; Temoche-Diaz, M.M.; Schekman, R.; Lambowitz, A.M. Broad Role for YBX1 in Defining the Small Noncoding RNA Composition of Exosomes. Proc. Natl. Acad. Sci. USA 2017, 114, E8987–E8995. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gan, Y.; Xu, G.; Hua, K.; Liu, D. Exosomes from MSCs Overexpressing MicroRNA-223-3p Attenuate Cerebral Ischemia through Inhibiting Microglial M1 Polarization Mediated Inflammation. Life Sci. 2020, 260, 118403. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Yao, H.; Chaudhuri, A.D.; Duan, M.; Yelamanchili, S.V.; Wen, H.; Cheney, P.D.; Fox, H.S.; Buch, S. Exosome-Mediated Shuttling of MicroRNA-29 Regulates HIV Tat and Morphine-Mediated Neuronal Dysfunction. Cell Death Dis. 2012, 3, e381. [Google Scholar] [CrossRef]
- You, Y.; Borgmann, K.; Edara, V.V.; Stacy, S.; Ghorpade, A.; Ikezu, T. Activated Human Astrocyte-Derived Extracellular Vesicles Modulate Neuronal Uptake, Differentiation and Firing. J. Extracell. Vesicles 2020, 9, 1706801. [Google Scholar] [CrossRef]
- Morioka, K.; Marmor, Y.; Sacramento, J.A.; Lin, A.; Shao, T.; Miclau, K.R.; Clark, D.R.; Beattie, M.S.; Marcucio, R.S.; Miclau, T.; et al. Differential Fracture Response to Traumatic Brain Injury Suggests Dominance of Neuroinflammatory Response in Polytrauma. Sci. Rep. 2019, 9, 12199. [Google Scholar] [CrossRef]
- Watson, C.N.; Belli, A.; Di Pietro, V. Small Non-Coding RNAs: New Class of Biomarkers and Potential Therapeutic Targets in Neurodegenerative Disease. Front. Genet. 2019, 10, 364. [Google Scholar] [CrossRef]
- Khan, N.Z.; Cao, T.; He, J.; Ritzel, R.M.; Li, Y.; Henry, R.J.; Colson, C.; Stoica, B.A.; Faden, A.I.; Wu, J. Spinal Cord Injury Alters MicroRNA and CD81+ Exosome Levels in Plasma Extracellular Nanoparticles with Neuroinflammatory Potential. Brain. Behav. Immun. 2021, 92, 165–183. [Google Scholar] [CrossRef]
- Gupta, S.; Rawat, S.; Arora, V.; Kottarath, S.K.; Dinda, A.K.; Vaishnav, P.K.; Nayak, B.; Mohanty, S. An Improvised One-Step Sucrose Cushion Ultracentrifugation Method for Exosome Isolation from Culture Supernatants of Mesenchymal Stem Cells. Stem Cell Res. Ther. 2018, 9, 180. [Google Scholar] [CrossRef]
- Osti, D.; del Bene, M.; Rappa, G.; Santos, M.; Matafora, V.; Richichi, C.; Faletti, S.; Beznoussenko, G.V.; Mironov, A.; Bachi, A.; et al. Clinical Significance of Extracellular Vesicles in Plasma from Glioblastoma Patients. Clin. Cancer Res. 2019, 25, 266–276. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| sncRNA | Condition | Role Associated with Early Neurodegeneration Conditions or Risk of Neurodegeneration | Role in Neurodegenerative Disease or Effects in CNS Cells | Additional Roles and Effects in Peripheral Cells |
|---|---|---|---|---|
| Let-7f | Il-1β stimulation, ischemia | Upregulated in ADEV after Il-1β stimulation in primary rat astrocytes [82], upregulated in rat pMCAO ischemia model after lesion [89] | Sporadic ALS downregulated biomarker [90], upregulated in AD hippocampus [91]. Targets involved in FoxO and MAPK signaling pathways and apoptosis [90]. Increased after HFD in rat [92]. • Promotes differentiation in rat neural stem cells [93]. Downregulates NDRG3 expression in rat cortical neurons to regulate hypoxia response (proapoptotic) [89]. ‡ Activate TLR7 [94]. ⁑ Downregulated in glioma cell lines, inhibits proliferation and migration, increases apoptosis [95]. ∞, * Protection against oxidative damage [96]. ∞, ‡ | Tumor suppressor miRNA, targets the aromatase gene (CYP19A1) [97]. ∞ |
| miR-16-5p | SCI, TBI, Il-1β and TNF-α stimulation | Upregulated in ADEV after Il-1β and TNF-α stimulation in primary rat astrocytes [82]. Upregulated in rat tissue after SCI [98]. Downregulated in human serum and mouse model after TBI [99]. | Biomarker, dysregulated in ALS serum, showed lower expression in slower progressing ALS [100]. Key implication in subacute stage of SCI [101]. • Reduce dendritic complexity and growth, spike rates and burst activity after inflammatory stimulus. Downregulates NTRK3 and Bcl2 [82]. Stimulates apoptosis and inflammatory proteins, Targets Apelin-13 inactivating ERK1/2 pathway [98]. ‡ Increases apoptosis, causes inflammation [102]. ⁑ Downregulated in glioma cell lines, targets TLN1 to increase glioma viability, proliferation, migration and invasion after TIIA [103]. ∞, * | Decreases fracture healing. Negatively regulates Bcl-2 and Cyclin-D1, therefore suppressing osteogenic differentiation, and osteoblast proliferation and survival. Inhibited proliferation promoting cell-cycle arrest and apoptosis [99]. π |
| miR-17-5p | OGD, HIBD | Downregulated in ADEV after HIBD in rat [87]. | Potential regulator of robust differentially expressed genes causing downregulation of GABAergic synapse and signaling pathways in AD [104]. It also counter IRE1a pathway downregulating TXNIP, NL3P inflammasome activation and Il-1β production [105]. Reduces inflammation related proteins after HIBD with lower production of TNF-α and IL-1β [87]. • Reduces neuronal death and apoptosis after HIBD [87]. ‡ Promotes proliferation of activated astrocytes after SCI [106]. * Upregulated in AD, in microglia adjacent to Aβ deposits. Targets autophagy receptor NBR1 inhibiting clearance of Aβ [107,108]. ⁑ Targets APP expression [109], BNIP2, SOD, GSH-Px and CAT expression, and reduces apoptosis and inflammation after OGD [87]. ∞, ⁑ | Expression increased after myocardial infarction. Inhibition associated with cardiomyocyte survival through STAT3 targeting [110]. γ Promotes osteoclastogenesis via targeting PTEN [111]. Promotes osteogenic differentiation and ossification, and cytokines such as VEGF [112]. π Upregulated in various cancers, Reduced proliferation in GIST tissue samples, targets KIT expression [113]. ∞ |
| mir-21 | Ischemia | Identified in ADEV from ALS mSOD1 mouse model with neurodegeneration stage not specified, upregulated [88]. Overexpressed in hippocampus after ischemia in rat [114]. | Reduced expression in axons in alcoholism and depression [115]. Suppresses OGD induced apoptosis, and Faslg pro-apoptotic factor levels. Upregulated in neurons of the IBZ [116]. ‡ Upregulation in mSOD1 ADEV mouse model is stopped after miR-146a induction [88]. After ISCI injury, stimulates polarization of reactive neurotrophic neuroprotective astrocytes [117]. * Repress FasL in microglia o inhibit neurotoxic hypoxia activated microglia [118]. ⁑ Diminish apoptosis modulating tumor suppressor PDC4I3K/AKT/GSK-3β, including apoptosis triggered by neurotoxic Aβ1–42 in SH-SY5Y [119]. PTEN independent oncogene [115]. ∞ ‡ | Upregulates VEGF promoting angiogenesis in transformed and non-transformed non CNS tumoral cell lines [120]. ∞ |
| miR-30b-5p | Ischemia | Upregulated in ADEV under IPC in rat [84]. | Differentially upregulated in MS, associated with non-progressive forms of the disease [9]. Downregulated in ALS [121]. Elevated in PD 6- hydroxydopamine induced rat models [122]. • | Involved in diabetic retinopathy with possible biomarker applications, regulates angiogenesis [123]. ⸰ Targets SIRT1 inhibiting autophagy of the mitochondria [122]. Suppresses lysosomal biogenesis and autophagy by inhibiting TFEB targets pre-transcriptionally [124]. ∞ |
| miR-32 | HIBD | Downregulated in ADEV after HIBD in rat [87]. | Involved in the maintenance of myelin fine tuning SLC45A3 and CLDN-11 expression [125]. ˟ Downregulated in glioma, targeted ABCC4 and EZH2, it reduces proliferation and migration [126]. ∞, * | Upregulated in some cancer tissues, reduces apoptosis and promotes proliferation and migration targeting OTUD3 and promoting MYC [127,128]. ∞ |
| miR-92b-3p | Ischemia, OSA, ASCI | Upregulated in ADEV under IPC in rat [84]. | Reduces inflammation after ischemic stroke. Downregulated in OSA, apnea, hypopnea [129]. Downregulated after ASCI, promotes functional recovery after ASCI [130]. • Diminishes apoptosis, cell death, mitochondrial dysfunction and favors neurite growth [84,130], including IHR induced apoptosis. Decreases ROS production, MAOA hyperactivity and PTEN expression. Promotes phosphorylation of AKT, and GAP43 and NF-200 expression [129,130]. ‡ Inhibits IHR-induced NF-κB1, PTGS1, TNF-α, and TGF-β expression [129]. + | Downregulated under hypoxia conditions in PASMCs, reverse proliferation and cell cycle induced under hypoxia conditions [131]. ζ Inhibits IHR-induced apoptosis and CXCL5 and ADRB1 expression [129]. ∞, α Regulate proliferation, apoptosis, differentiation, and metastasis [131]. ∞ |
| mir-100 | Il-1β stimulation | Upregulated in ADEV under Il-1β and ATP induction in primary rat astrocytes [82]. | Downregulated in autosomal recessive NDD SMA [132,133]. ⸸ Induces apoptosis in retinal ganglion cells exposed to H2O2 [134]. ‡ Activates TLR8 receptors post-translationally causing indirect neuronal microenvironment dysregulation, activates cytokine and chemokine release [135]. Downregulates in activated microglia, ameliorates motor function loss after SCI by targeting TLR4 and NF-κB [136]. ⁑ | Activates cytokine and chemokine release in macrophages [135]. α Downregulated in hypoxia induced proliferation of PASMCs, suppress mTOR expression leading to inhibition of proliferation [137]. ζ Overexpressed in EVs from CRC cells mutant KRAS expressing [138]. ∞, ο |
| miR-107 | TNF-α stimulation | Upregulated in ADEV after TNF-α stimulation in primary rat astrocytes [82]. | Involved in AD, targets BACE1, CDK5, ADAM1, increases neuronal differentiation [139]. • Higher expression correlates with lower overall patient survival in high grade gliomas [140]. ∞ | Alter key aggressiveness characteristics of prostate cancer cells such as proliferation, modulates lipid metabolism, adjacent non-tumoral tissue shows downregulation. Its expression in cancer correlates with levels in plasma [141]. ∞ |
| mir-138 | Morphine-mediated neuroinflammatory microenvironment | Upregulated in ADEV under morphine stimulated conditions in mouse [77]. | Activates astrocytes induced by Tat in HAND [142]. * Internalizes mir-138 charged ADEV. Activation of microglia through direct activation of the TLR7-NF-kB axis [77]. ⁑ Promotes early differentiation of oligodendrocytes [143]. ˟ | Inhibits adipocyte differentiation reducing EZH2 expression [144]. μ |
| miR-146a | Early asymptomatic mSOD1 ALS model, ethanol activated neuroinflammation | Upregulated in ADEV after ethanol induction in mouse astrocytes [85], Identified in ADEV from ALS mSOD1 mouse model with neurodegeneration stage not specified, downregulated [88]. | Involvement in pathogenesis of MS, AD, prion disease, neurotropic virus and metal sulfate induced toxicity [145] and neuroprotective in specific contexts in ALS and stroke [88,146]. • Upregulated after stroke in NPC. Increases myelinization protein expression and differentiation towards oligodendrocyte lineage [146] ⸸, ˟ Increases inflammation and its own expression in cortical neurons [85]. ‡ Attenuates miR-21 and miR-155 expression in ALS mSOD1 mouse model, decreases astrocyte reactivity and decreases proinflammatory miRNA associated exosomal cargo production [88]. Downregulated in ALS [147]. * | Downregulated by infection (RCV virus), it diminish TRAF-6 expression, JNK activation and lung inflammatory infiltration, reduces L-1β, IL -6 and TNF-α production [148]. ε |
| miR-155 | EAE Neuroinflammation, proinflammatory cytokines | Identified in ADEV from ALS mSOD1 mouse model with neurodegeneration stage not specified, upregulated [88]. Downregulated in spinal cord of mice with EAE [149]. | Globally upregulated in AD, related with inflammation targets CFH [139,145]. Upregulated in EAE MS mouse model [150], it expression is very high in MS lesions, favors proinflammatory conditions and negatively regulates BBB [145,149]. • Downregulated in ADEV in ALS mSOD1 mouse model by miR-146a induction [88] * Upregulated by proinflammatory cytokines [88]. Downregulates fatty acid metabolism associated genes [149]. + | Increases macrophage migration, mediates activation of mononuclear phagocytes, [151]. α Constitutively highly expressed [145], promotes differentiation of TH17 cells and activation of T-cells and dendritic cells [149]. δ ROS diminish miR-155-5p expression in tumor exosomes leading to immunosuppressive tumor growth [151]. ∞ |
| mir-182 | SCI, HIBD, LPS Ethanol induced neuro-inflammation | Downregulated in ADEV after HIBD in rat [87], upregulated in ADEV after ethanol induction in mouse astrocytes [85]. Downregulated in SCI of mice [152]. | Anti-inflammatory miR [152]. After ischaemia exacerbates BBB dysfunction [153]. • Enriched in neurons. Increased dendrite tree complexity, axon and neurite outgrowth, favoring expression of neurofilament-M and neurofilament-L, and AKT phosphorylation [154]. Improves SCI reducing apoptosis [152]. ⸸, ‡ Increases in ethanol-treated wild type astrocytes in a TLR4-dependent response [85]. * Inflammatory suppressor (downregulates TNF-α, IL-6, IL-1β), apoptosis reduction by caspase-3 downregulation. Decreases expression after LPS [152]. ∞, ⁑ Targeted MTSS1 tumor suppressor transcript to inhibit proliferation and migration in glioma [155]. ∞ | Inhibited apoptosis regulating PDCD4 and PACS2, under non-ischemic heart failure [156]. γ Downregulated after Ischemia Reperfusion (I/R). It reduces autophagy stimulating mTOR and targeting Deptor, thus reducing lesion area after I/R [157]. ο Biomarker in prostate cancer [158]. ∞ |
| mir-200b | Ethanol induced neuro-inflammation | Upregulated in ADEV after ethanol induction in mouse astrocytes [85], downregulated in ADEV after HIBD in rat [87]. | Targets APP gene downregulating amyloid beta (Aβ), however Aβ42 inhibits its expression (possibly halts AD progression) [159]. • Increases in ethanol treated wild type astrocytes in a TLR4-dependent response [85]. * Upregulated by Aβ, its transfection reduces Aβ secretion in conditioned media, relieve memory impairments and downregulates targets such as IRS-1pSer potentially diminishing insulin resistance [160]. ‡ | Under glucotoxicity increases apoptosis of human retinal pigment epithelial cells [161]. ε |
| miR-873a-5p | TBI | Upregulated after TBI induction in mouse [86]. | Improve neurological deficits associated with TBI, exhibiting a neuroprotective role regulating inflammatory signals [86]. Targets A20 (TNFAIP3) and targeted by HOTAIRM1 a miR-sponge associated with neuronal apoptosis [162,163]. • Released by activated astrocytes [86]. * Decreases ERK and NF-κB p65 phosphorylation inhibiting LPS induced M1 phenotype and inflammatory signaling, promotes microglia M2 polarization after TBI [86]. ⁑ | Proapoptotic inhibitor of cell growth, downregulated in glioblastoma [164]. ∞ |
| sncRNA | Neuroprotection | Neurodegeneration |
|---|---|---|
| Let-7f | [93,96] | |
| miR-16-5p | [102,197] | |
| miR-17-5p | [87,105] | [108] |
| mir-21 | [116,117,119] | |
| miR-30b-5p | [122] | |
| miR-32 | [125] | |
| miR-92b-3p | [84,129,130] | |
| mir-100 | [136] | [134,135] |
| miR-107 | [139] | |
| mir-138 | [76,142] | |
| miR-146a | [88,146] | [85,145] |
| miR-155 | [88,145,149,151] | |
| mir-182 | [152,154] | [153] |
| mir-200b | [159,160] | |
| miR-873a-5p | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Cepeda, L.; Castro, J.D.; Aristizábal-Pachón, A.F.; González-Giraldo, Y.; Pinzón, A.; Puentes-Rozo, P.J.; González, J. Modulation of Small RNA Signatures by Astrocytes on Early Neurodegeneration Stages; Implications for Biomarker Discovery. Life 2022, 12, 1720. https://doi.org/10.3390/life12111720
López-Cepeda L, Castro JD, Aristizábal-Pachón AF, González-Giraldo Y, Pinzón A, Puentes-Rozo PJ, González J. Modulation of Small RNA Signatures by Astrocytes on Early Neurodegeneration Stages; Implications for Biomarker Discovery. Life. 2022; 12(11):1720. https://doi.org/10.3390/life12111720
Chicago/Turabian StyleLópez-Cepeda, Leonardo, Juan David Castro, Andrés Felipe Aristizábal-Pachón, Yeimy González-Giraldo, Andrés Pinzón, Pedro J. Puentes-Rozo, and Janneth González. 2022. "Modulation of Small RNA Signatures by Astrocytes on Early Neurodegeneration Stages; Implications for Biomarker Discovery" Life 12, no. 11: 1720. https://doi.org/10.3390/life12111720
APA StyleLópez-Cepeda, L., Castro, J. D., Aristizábal-Pachón, A. F., González-Giraldo, Y., Pinzón, A., Puentes-Rozo, P. J., & González, J. (2022). Modulation of Small RNA Signatures by Astrocytes on Early Neurodegeneration Stages; Implications for Biomarker Discovery. Life, 12(11), 1720. https://doi.org/10.3390/life12111720