
Telocytes in the Female Reproductive System: Up-to-Date Knowledge, Challenges and Possible Clinical Applications



{kind=link}
Abstract
1. Introduction
2. General Information on Telocytes—Morphology and Physiological Functions
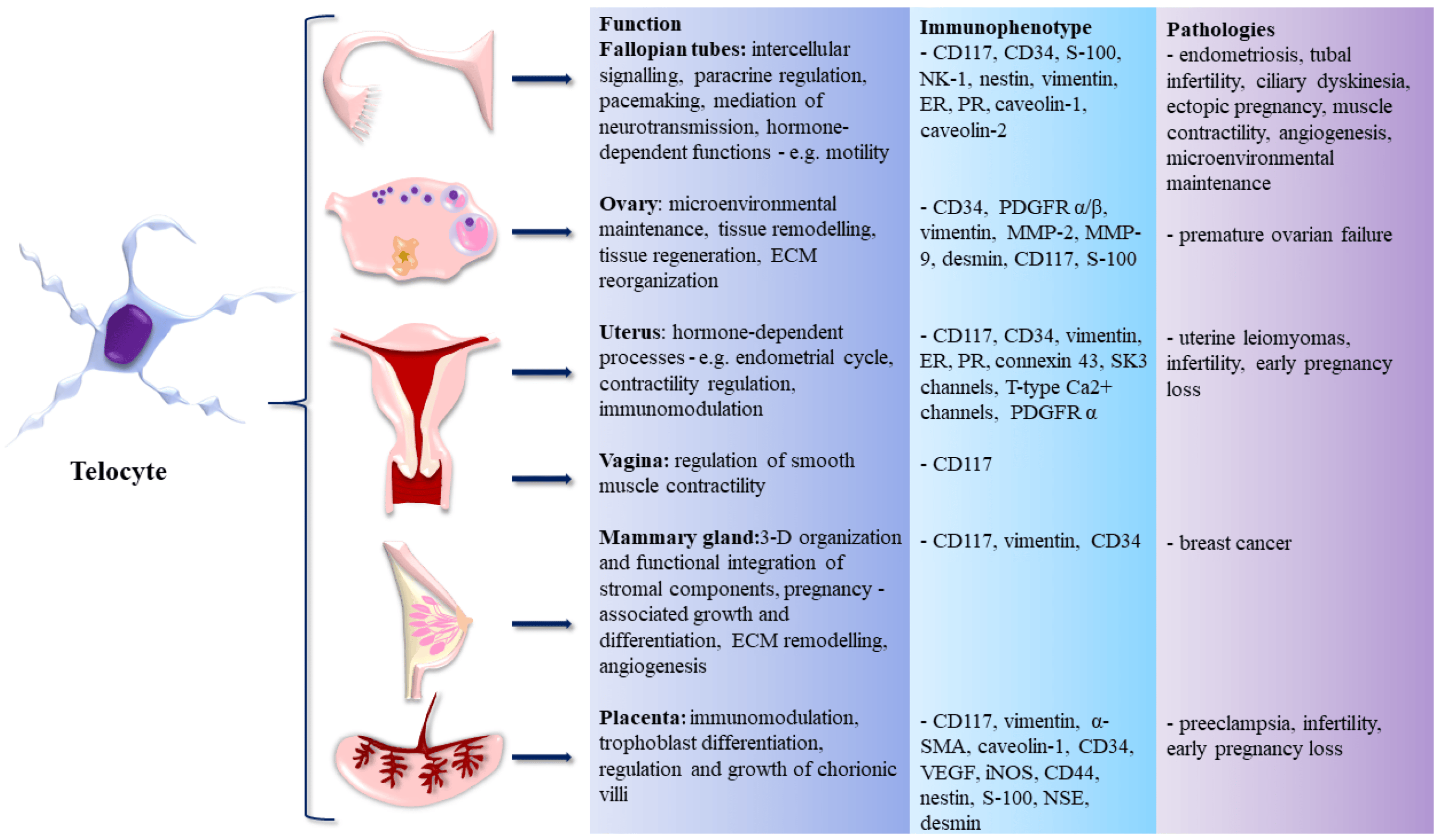
3. Morphological and Functional Specifics of FRS Telocytes
4. Immunophenotype of FRS Telocytes
5. Pathology of FRS Telocytes
6. Telocytes and FRS Regeneration
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ChAT | choline acetyltransferase |
| ESCs | endometrial stromal cells |
| ER | estrogen receptor |
| ECM | extracellular matrix |
| eGISTs | extragastrointestinal stromal tumors |
| FRS | female reproductive system |
| iNOS | inducible nitric oxide synthase |
| ICLCs | interstitial Cajal-like cells |
| ICCs | interstitial cells of Cajal |
| LLLS | low-level-laser stimulation |
| MMP | matrix metalloproteinase |
| NSE | neuron-specific enolase |
| PDGFR | platelet-derived growth factor receptor |
| PR | progesterone receptor |
| PGP 9.5 | protein gene product 9.5 |
| SMA | smooth muscle actin |
| TCs | telocytes |
| 3-D | three-dimensional |
| VEGF | vascular endothelial growth factor |
References
- Clark, G.F.; Schust, D.J. Manifestations of immune tolerance in the human female reproductive tract. Front. Immunol 2013, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Bates, G.W.; Bowling, M. Physiology of the female reproductive axis. Periodontol. 2000 2013, 61, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Schauer, C.; Tong, T.; Petitjean, H.; Blum, T.; Peron, S.; Mai, O.; Schmitz, F.; Boehm, U.; Leinders-Zufall, T. Hypothalamic gonadotropin-releasing hormone (GnRH) receptor neurons fire in synchrony with the female reproductive cycle. J. Neurophysiol. 2015, 114, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Ledger, W.L. Demographics of infertility. Reprod. Biomed. Online 2009, 18 (Suppl. S2), 11–14. [Google Scholar] [CrossRef]
- Weiderpass, E.; Labrèche, F. Malignant tumors of the female reproductive system. Saf. Health Work 2012, 3, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Janas, P.; Kucybała, I.; Radoń-Pokracka, M.; Huras, H. Telocytes in the female reproductive system: An overview of up-to-date knowledge. Adv. Clin. Exp. Med. 2018, 27, 559–565. [Google Scholar] [CrossRef]
- Junquera, C.; Martínez-Ciriano, C.; Castiella, T.; Serrano, P.; Azanza, M.J.; Junquera, S.R. Immunohistochemical and ultrastructural characteristics of interstitial cells of Cajal in the rabbit duodenum. Presence of a single cilium. J. Cell Mol. Med. 2007, 11, 776–787. [Google Scholar] [CrossRef]
- Faussone-Pellegrini, M.S.; Pantalone, D.; Cortesini, C. Smooth muscle cells, interstitial cells of Cajal and myenteric plexus interrelationships in the human colon. Acta Anat. 1990, 139, 31–44. [Google Scholar] [CrossRef]
- Popescu, L.M.; Faussone-Pellegrini, M.S. TELOCYTES—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. Biomol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef]
- Kucybala, I.; Janas, P.; Ciuk, S.; Cholopiak, W.; Klimek-Piotrowska, W.; Holda, M.K. A comprehensive guide to telocytes and their great potential in cardiovascular system. Bratisl. Lek. Listy 2017, 118, 302–309. [Google Scholar] [CrossRef]
- Cretoiu, D.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, S.M. Telocytes heterogeneity: From cellular morphology to functional evidence. Semin. Cell Dev. Biol. 2017, 64, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Gherghiceanu, M.; Suciu, L.C.; Manole, C.G.; Hinescu, M.E. Telocytes and putative stem cells in the lungs: Electron microscopy, electron tomography and laser scanning microscopy. Cell Tissue Res. 2011, 345, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Kondo, A.; Kaestner, K.H. Emerging diverse roles of telocytes. Development 2019, 146, dev175018. [Google Scholar] [CrossRef]
- Urban, L.; Miko, M.; Kajanova, M.; Bozikova, S.; Mrazova, H.; Varga, I. Telocytes (interstitial Cajal-like cells) in human Fallopian tubes. Bratisl. Lek. Listy 2016, 117, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Bei, Y.; Wang, F.; Yang, C.; Xiao, J. Telocytes in regenerative medicine. J. Cell. Mol. Med. 2015, 19, 1441–1454. [Google Scholar] [CrossRef]
- Cretoiu, D.; Xu, J.; Xiao, J.; Cretoiu, S.M. Telocytes and Their Extracellular Vesicles-Evidence and Hypotheses. Int. J. Mol. Sci. 2016, 17, 1322. [Google Scholar] [CrossRef]
- Verdile, N.; Pasquariello, R.; Cardinaletti, G.; Tibaldi, E.; Brevini, T.A.L.; Gandolfi, F. Telocytes: Active Players in the Rainbow Trout (Oncorhynchus mykiss) Intestinal Stem-Cell Niche. Animals 2021, 12, 74. [Google Scholar] [CrossRef]
- Tang, L.; Song, D.; Qi, R.; Zhu, B.; Wang, X. Roles of pulmonary telocytes in airway epithelia to benefit experimental acute lung injury through production of telocyte-driven mediators and exosomes. Cell Biol. Toxicol. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Ravalli, S.; Federico, C.; Lauretta, G.; Saccone, S.; Pricoco, E.; Roggio, F.; Di Rosa, M.; Maugeri, G.; Musumeci, G. Morphological Evidence of Telocytes in Skeletal Muscle Interstitium of Exercised and Sedentary Rodents. Biomedicines 2021, 9, 807. [Google Scholar] [CrossRef]
- Chen, X.; Zeng, J.; Huang, Y.; Gong, M.; Ye, Y.; Zhao, H.; Chen, Z.; Zhang, H. Telocytes and their structural relationships with surrounding cell types in the skin of silky fowl by immunohistochemistrical, transmission electron microscopical and morphometric analysis. Poult Sci 2021, 100, 101367. [Google Scholar] [CrossRef] [PubMed]
- Wishahi, M.; Hafiz, E.; Wishahy, A.M.K.; Badawy, M. Telocytes, c-Kit positive cells, Smooth muscles, and collagen in the ureter of pediatric patients with congenital primary obstructive megaureter: Elucidation of etiopathology. Ultrastruct. Pathol. 2021, 45, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Sukhacheva, T.V.; Nizyaeva, N.V.; Samsonova, M.V.; Cherniaev, A.L.; Burov, A.A.; Iurova, M.V.; Shchegolev, A.I.; Serov, R.A.; Sukhikh, G.T. Morpho-functional changes of cardiac telocytes in isolated atrial amyloidosis in patients with atrial fibrillation. Sci. Rep. 2021, 11, 3563. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.L.A.; Braz, J.; Martins, G.M.; Clebis, N.K.; Oliveira, M.F.; Morais, D.B.; Moura, C.E.B. Identification of telocytes in dystrophic mice testis. Einstein (Sao Paulo) 2021, 19, eAI5737. [Google Scholar] [CrossRef] [PubMed]
- Felisbino, S.L.; Sanches, B.D.A.; Delella, F.K.; Scarano, W.R.; Dos Santos, F.C.A.; Vilamaior, P.S.L.; Taboga, S.R.; Justulin, L.A. Prostate telocytes change their phenotype in response to castration or testosterone replacement. Sci. Rep. 2019, 9, 3761. [Google Scholar] [CrossRef]
- Nicolescu, M.I.; Rusu, M.C.; Voinea, L.M.; Vrapciu, A.D.; Bâră, R.I. Lymphatic lacunae of the human eye conjunctiva embedded within a stroma containing CD34(+) telocytes. J. Cell. Mol. Med. 2020, 24, 8871–8875. [Google Scholar] [CrossRef]
- Wagener, M.G.; Leonhard-Marek, S.; Häger, J.D.; Pfarrer, C. CD117- and vimentin-positive telocytes in the bovine teat sphincter. Anat. Histol. Embryol. 2018, 47, 268–270. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Faussone-Pellegrini, M.S. The Telocyte Subtypes. Adv. Exp. Med. Biol. 2016, 913, 115–126. [Google Scholar]
- Liu, T.; Wang, S.; Li, Q.; Huang, Y.; Chen, C.; Zheng, J. Telocytes as potential targets in a cyclophosphamide-induced animal model of premature ovarian failure. Mol. Med. Rep. 2016, 14, 2415–2422. [Google Scholar] [CrossRef]
- Mazzoni, T.S.; Viadanna, R.R.; Quagio-Grassiotto, I. Presence, localization and morphology of TELOCYTES in developmental gonads of fishes. J. Morphol. 2019, 280, 654–665. [Google Scholar] [CrossRef]
- Mokhtar, D.M. Characterization of the fish ovarian stroma during the spawning season: Cytochemical, immunohistochemical and ultrastructural studies. Fish. Shellfish. Immunol. 2019, 94, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, D.M.; Hussein, M.M. Microanalysis of Fish Ovarian Follicular Atresia: A Possible Synergic Action of Somatic and Immune Cells. Microsc. Microanal. 2020, 26, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Ciontea, S.M.; Cretoiu, D.; Hinescu, M.E.; Radu, E.; Ionescu, N.; Ceausu, M.; Gherghiceanu, M.; Braga, R.I.; Vasilescu, F.; et al. Novel type of interstitial cell (Cajal-like) in human fallopian tube. J. Cell. Mol. Med. 2005, 9, 479–523. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Ciontea, S.M.; Cretoiu, D. Interstitial Cajal-like cells in human uterus and fallopian tube. Ann. N. Y. Acad. Sci. 2007, 1101, 139–165. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Cretoiu, D.; Suciu, L.; Popescu, L.M. Interstitial Cajal-like cells of human Fallopian tube express estrogen and progesterone receptors. J. Mol. Histol. 2009, 40, 387–394. [Google Scholar] [CrossRef]
- Cretoiu, D.; Ciontea, S.M.; Popescu, L.M.; Ceafalan, L.; Ardeleanu, C. Interstitial Cajal-like cells (ICLC) as steroid hormone sensors in human myometrium: Immunocytochemical approach. J. Cell. Mol. Med. 2006, 10, 789–795. [Google Scholar] [CrossRef]
- Roatesi, I.; Radu, B.M.; Cretoiu, D.; Cretoiu, S.M. Uterine Telocytes: A Review of Current Knowledge. Biol. Reprod. 2015, 93, 10. [Google Scholar] [CrossRef]
- Salama, N.M. Immunohistochemical characterization of telocytes in ratuterus in different reproductive states. Egypt. J. Histol. 2013, 36, 185–194. [Google Scholar] [CrossRef]
- Tang, X.L.; Zhang, F.L.; Jiang, X.J.; Yang, X.J. Telocytes enhanced the proliferation, adhesion and motility of endometrial stromal cells as mediated by the ERK pathway in vitro. Am. J. Transl. Res. 2019, 11, 572–585. [Google Scholar]
- Banciu, A.; Banciu, D.D.; Mustaciosu, C.C.; Radu, M.; Cretoiu, D.; Xiao, J.; Cretoiu, S.M.; Suciu, N.; Radu, B.M. Beta-Estradiol Regulates Voltage-Gated Calcium Channels and Estrogen Receptors in Telocytes from Human Myometrium. Int. J. Mol. Sci. 2018, 19, 1413. [Google Scholar] [CrossRef]
- Jiang, X.J.; Cretoiu, D.; Shen, Z.J.; Yang, X.J. An in vitro investigation of telocytes-educated macrophages: Morphology, heterocellular junctions, apoptosis and invasion analysis. J. Transl. Med. 2018, 16, 85. [Google Scholar] [CrossRef] [PubMed]
- Shafik, A.; El-Sibai, O.; Shafik, I.; Shafik, A.A. Immunohistochemical identification of the pacemaker cajal cells in the normal human vagina. Arch. Gynecol. Obstet. 2005, 272, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Nizyaeva, N.V.; Sukhacheva, T.V.; Kulikova, G.V.; Nagovitsyna, M.N.; Poltavtseva, R.A.; Kan, N.E.; Tyutyunnik, V.L.; Pavlovich, S.V.; Serov, R.A.; Shchyogolev, A.I. Ultrastructural Characteristics of Placental Telocytes. Bull. Exp. Biol. Med. 2017, 162, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Gherghiceanu, M.; Popescu, L.M. Interstitial Cajal-like cells (ICLC) in human resting mammary gland stroma. Transmission electron microscope (TEM) identification. J. Cell. Mol. Med. 2005, 9, 893–910. [Google Scholar] [CrossRef]
- Petre, N.; Rusu, M.C.; Pop, F.; Jianu, A.M. Telocytes of the mammary gland stroma. Folia Morphol. (Warsz) 2016, 75, 224–231. [Google Scholar] [CrossRef]
- Sanches, B.D.A.; Leonel, E.C.R.; Maldarine, J.S.; Tamarindo, G.H.; Barquilha, C.N.; Felisbino, S.L.; Goés, R.M.; Vilamaior, P.S.L.; Taboga, S.R. Telocytes are associated with tissue remodeling and angiogenesis during the postlactational involution of the mammary gland in gerbils. Cell Biol. Int. 2020, 44, 2512–2523. [Google Scholar] [CrossRef]
- Cretoiu, S.M. Immunohistochemistry of Telocytes in the Uterus and Fallopian Tubes. Adv. Exp. Med. Biol. 2016, 913, 335–357. [Google Scholar]
- Ciontea, S.M.; Radu, E.; Regalia, T.; Ceafalan, L.; Cretoiu, D.; Gherghiceanu, M.; Braga, R.I.; Malincenco, M.; Zagrean, L.; Hinescu, M.E.; et al. C-kit immunopositive interstitial cells (Cajal-type) in human myometrium. J. Cell. Mol. Med. 2005, 9, 407–420. [Google Scholar] [CrossRef]
- Klein, M.; Urban, L.; Deckov, I.; Danisovic, L.; Polak, S.; Danihel, L.; Varga, I. Distribution of telocytes in the corpus and cervix of human uterus: An immunohistochemical study. Biologia 2017, 72, 1217–1223. [Google Scholar] [CrossRef]
- Hatta, K.; Huang, M.L.; Weisel, R.D.; Li, R.K. Culture of rat endometrial telocytes. J. Cell. Mol. Med. 2012, 16, 1392–1396. [Google Scholar] [CrossRef]
- Rosenbaum, S.T.; Svalø, J.; Nielsen, K.; Larsen, T.; Jørgensen, J.C.; Bouchelouche, P. Immunolocalization and expression of small-conductance calcium-activated potassium channels in human myometrium. J. Cell. Mol. Med. 2012, 16, 3001–3008. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, D.; Ceafalan, L.C.; Popescu, L.M. Isolated human uterine telocytes: Immunocytochemistry and electrophysiology of T-type calcium channels. Histochem. Cell Biol. 2015, 143, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Suciu, L.; Popescu, L.M.; Gherghiceanu, M. Human placenta: De visu demonstration of interstitial Cajal-like cells. J. Cell. Mol. Med. 2007, 11, 590–597. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Suciu, L.; Popescu, L.M.; Gherghiceanu, M.; Regalia, T.; Nicolescu, M.I.; Hinescu, M.E.; Faussone-Pellegrini, M.S. Telocytes in human term placenta: Morphology and phenotype. Cells Tissues Organs 2010, 192, 325–339. [Google Scholar] [CrossRef]
- Mou, Y.; Wang, Y.; Li, J.; Lü, S.; Duan, C.; Du, Z.; Yang, G.; Chen, W.; Zhao, S.; Zhou, J.; et al. Immunohistochemical characterization and functional identification of mammary gland telocytes in the self-assembly of reconstituted breast cancer tissue in vitro. J. Cell. Mol. Med. 2013, 17, 65–75. [Google Scholar] [CrossRef]
- El-Tahawy, N.F.G.; Rifaai, R.A. Immunohistochemical and ultrastructural evidence for telocytes in the different physiological stages of the female rat mammary gland. Life Sci. 2019, 231, 116521. [Google Scholar] [CrossRef]
- Aleksandrovych, V.; Wrona, A.; Bereza, T.; Pityński, K.; Gil, K. Oviductal Telocytes in Patients with Uterine Myoma. Biomedicines 2021, 9, 1060. [Google Scholar] [CrossRef]
- Karasu, Y.; Önal, D.; Zırh, S.; Yersal, N.; Korkmaz, H.; Üstün, Y.; Müftüoğlu, S.; Pehlivanoğlu, B. Role of telocytes in the pathogenesis of ectopic pregnancy. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 110–119. [Google Scholar]
- Stewart, E.A.; Cookson, C.L.; Gandolfo, R.A.; Schulze-Rath, R. Epidemiology of uterine fibroids: A systematic review. BJOG 2017, 124, 1501–1512. [Google Scholar] [CrossRef]
- Aleksandrovych, V.; Białas, M.; Pasternak, A.; Bereza, T.; Sajewicz, M.; Walocha, J.; Gil, K. Identification of uterine telocytes and their architecture in leiomyoma. Folia Med. Cracov. 2018, 58, 89–102. [Google Scholar]
- Varga, I.; Klein, M.; Urban, L.; Danihel, L., Jr.; Polak, S.; Danihel, L., Sr. Recently discovered interstitial cells “telocytes” as players in the pathogenesis of uterine leiomyomas. Med. Hypotheses 2018, 110, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Padhi, S.; Nayak, H.K. Primary Extragastrointestinal Stromal Tumours in the Hepatobiliary Tree and Telocytes. Adv. Exp. Med. Biol. 2016, 913, 207–228. [Google Scholar] [PubMed]
- Fletcher, N.M.; Saed, M.G.; Abu-Soud, H.M.; Al-Hendy, A.; Diamond, M.P.; Saed, G.M. Uterine fibroids are characterized by an impaired antioxidant cellular system: Potential role of hypoxia in the pathophysiology of uterine fibroids. J. Assist. Reprod. Genet. 2013, 30, 969–974. [Google Scholar] [CrossRef]
- Aleksandrovych, V.; Bereza, T.; Ulatowska-Białas, M.; Pasternak, A.; Walocha, J.A.; Pityński, K.; Gil, K. Identification of PDGFRα + cells in uterine fibroids—Link between angiogenesis and uterine telocytes. Arch. Med. Sci. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Chi, C.; Jiang, X.J.; Su, L.; Shen, Z.J.; Yang, X.J. In vitro morphology, viability and cytokine secretion of uterine telocyte-activated mouse peritoneal macrophages. J. Cell. Mol. Med. 2015, 19, 2741–2750. [Google Scholar] [CrossRef]
- Skowron, K.; Aleksandrovych, V.; Kurnik-Łucka, M.; Stach, P.; Baranowska, A.; Skowron, B.; Gil, K. Aberrations in the female reproductive organs and a role of telocytes in a rat model of anorexia nervosa. Folia Med. Cracov. 2018, 58, 115–125. [Google Scholar] [PubMed]
- Nirupama, R.; Divyashree, S.; Janhavi, P.; Muthukumar, S.P.; Ravindra, P.V. Preeclampsia: Pathophysiology and management. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 101975. [Google Scholar] [CrossRef]
- Bosco, C.B.; Díaz, E.G.; Gutierrez, R.R.; González, J.M.; Parra-Cordero, M.; Rodrigo, R.S.; Barja, P.Y. Placental Hypoxia Developed During Preeclampsia Induces Telocytes Apoptosis in Chorionic Villi Affecting The Maternal-Fetus Metabolic Exchange. Curr. Stem Cell Res. Ther. 2016, 11, 420–425. [Google Scholar] [CrossRef]
- Bosco, C.; Díaz, E.; Gutiérrez, R.; González, J.; Parra-Cordero, M.; Rodrigo, R.; Barja, P. A putative role for telocytes in placental barrier impairment during preeclampsia. Med. Hypotheses 2015, 84, 72–77. [Google Scholar] [CrossRef]
- Abu-Dief, E.E.; Elsayed, H.M.; Atia, E.W.; Abdel-Rahman, M.; Fawzy, M. Modulation of Telocytes in Women with Preeclampsia: A Prospective Comparative Study. J. Microsc. Ultrastruct. 2021, 9, 158–163. [Google Scholar] [CrossRef]
- Momenimovahed, Z.; Salehiniya, H. Epidemiological characteristics of and risk factors for breast cancer in the world. Breast Cancer (Dove Med. Press) 2019, 11, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, M.P.; Díaz-Flores, L., Jr.; Carrasco, J.L.; Martín-Vasallo, P. CD34+ Stromal Cells/Telocytes as a Source of Cancer-Associated Fibroblasts (CAFs) in Invasive Lobular Carcinoma of the Breast. Int. J. Mol. Sci. 2021, 22, 3686. [Google Scholar] [CrossRef] [PubMed]
- Ping, Q.; Yan, R.; Cheng, X.; Wang, W.; Zhong, Y.; Hou, Z.; Shi, Y.; Wang, C.; Li, R. Cancer-associated fibroblasts: Overview, progress, challenges, and directions. Cancer Gene Ther. 2021, 28, 984–999. [Google Scholar] [CrossRef]
- Yonghong, S.; Ruizhi, W.; Yue, Z.; Xuebing, B.; Tarique, I.; Chunhua, L.; Ping, Y.; Qiusheng, C. Telocytes in Different Organs of Vertebrates: Potential Essence Cells of the Meridian in Chinese Traditional Medicine. Microsc. Microanal. 2020, 26, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Polák, Š.; Kyselovič, J.; Kachlík, D.; Danišovič, Ľ.; Klein, M. Recently Discovered Interstitial Cell Population of Telocytes: Distinguishing Facts from Fiction Regarding Their Role in the Pathogenesis of Diverse Diseases Called “Telocytopathies”. Medicina 2019, 55, 56. [Google Scholar] [CrossRef]
- Magalhaes, R.S.; Atala, A. Chapter 70—Regenerative Medicine for the Female Reproductive System. In Principles of Regenerative Medicine, 3rd ed.; Atala, A., Lanza, R., Mikos, A.G., Nerem, R., Eds.; Academic Press: Boston, MA, USA, 2019; pp. 1237–1250. [Google Scholar] [CrossRef]
- Sysak, R.; Bluska, P.; Stencl, P.; Klein, M.; Varga, I. Agenesis of female internal reproductive organs, the Mayer- Rokitansky-Küster-Hauser syndrome. Bratisl. Lek. Listy 2021, 122, 839–845. [Google Scholar] [CrossRef]
- Campeanu, R.A.; Radu, B.M.; Cretoiu, S.M.; Banciu, D.D.; Banciu, A.; Cretoiu, D.; Popescu, L.M. Near-infrared low-level laser stimulation of telocytes from human myometrium. Lasers Med. Sci. 2014, 29, 1867–1874. [Google Scholar] [CrossRef]
- Popescu, L.M.; Gherghiceanu, M.; Manole, C.G.; Faussone-Pellegrini, M.S. Cardiac renewing: Interstitial Cajal-like cells nurse cardiomyocyte progenitors in epicardial stem cell niches. J. Cell. Mol. Med. 2009, 13, 866–886. [Google Scholar] [CrossRef]
- Popescu, L.M.; Manole, E.; Serboiu, C.S.; Manole, C.G.; Suciu, L.C.; Gherghiceanu, M.; Popescu, B.O. Identification of telocytes in skeletal muscle interstitium: Implication for muscle regeneration. J. Cell. Mol. Med. 2011, 15, 1379–1392. [Google Scholar] [CrossRef]
- Ceafalan, L.; Gherghiceanu, M.; Popescu, L.M.; Simionescu, O. Telocytes in human skin--are they involved in skin regeneration? J. Cell. Mol. Med. 2012, 16, 1405–1420. [Google Scholar] [CrossRef]
- Popescu, B.O.; Gherghiceanu, M.; Kostin, S.; Ceafalan, L.; Popescu, L.M. Telocytes in meninges and choroid plexus. Neurosci. Lett. 2012, 516, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Luesma, M.J.; Gherghiceanu, M.; Popescu, L.M. Telocytes and stem cells in limbus and uvea of mouse eye. J. Cell. Mol. Med. 2013, 17, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Song, Y.; Bei, Y.; Zhao, Y.; Xiao, J.; Yang, C. Telocytes in liver regeneration: Possible roles. J. Cell. Mol. Med. 2014, 18, 1720–1726. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lin, M.; Li, L.; Wang, R.; Zhang, C.; Qi, G.; Xu, M.; Rong, R.; Zhu, T. Renal telocytes contribute to the repair of ischemically injured renal tubules. J. Cell. Mol. Med. 2014, 18, 1144–1156. [Google Scholar] [CrossRef]
- Grounds, M.D. Obstacles and challenges for tissue engineering and regenerative medicine: Australian nuances. Clin. Exp. Pharmacol. Physiol. 2018, 45, 390–400. [Google Scholar] [CrossRef]
- Varga, I.; Kyselovič, J.; Danišovič, Ľ.; Gálfiová, P.; Kachlík, D.; Polák, Š.; Klein, M. Recently discovered interstitial cells termed telocytes: Distinguishing cell-biological and histological facts from fictions. Biologia 2019, 74, 195–203. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klein, M.; Csöbönyeiová, M.; Danišovič, Ľ.; Lapides, L.; Varga, I. Telocytes in the Female Reproductive System: Up-to-Date Knowledge, Challenges and Possible Clinical Applications. Life 2022, 12, 267. https://doi.org/10.3390/life12020267
Klein M, Csöbönyeiová M, Danišovič Ľ, Lapides L, Varga I. Telocytes in the Female Reproductive System: Up-to-Date Knowledge, Challenges and Possible Clinical Applications. Life. 2022; 12(2):267. https://doi.org/10.3390/life12020267
Chicago/Turabian StyleKlein, Martin, Mária Csöbönyeiová, Ľuboš Danišovič, Lenka Lapides, and Ivan Varga. 2022. "Telocytes in the Female Reproductive System: Up-to-Date Knowledge, Challenges and Possible Clinical Applications" Life 12, no. 2: 267. https://doi.org/10.3390/life12020267
APA StyleKlein, M., Csöbönyeiová, M., Danišovič, Ľ., Lapides, L., & Varga, I. (2022). Telocytes in the Female Reproductive System: Up-to-Date Knowledge, Challenges and Possible Clinical Applications. Life, 12(2), 267. https://doi.org/10.3390/life12020267