
SIRT1: Genetic Variants and Serum Levels in Age-Related Macular Degeneration

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Ophthalmologic Evaluation
2.3. DNA Extraction, Genotyping and SIRT1 Serum Level Determination
2.4. Statistical Analysis
3. Results
3.1. SIRT1 Haplotype Analysis
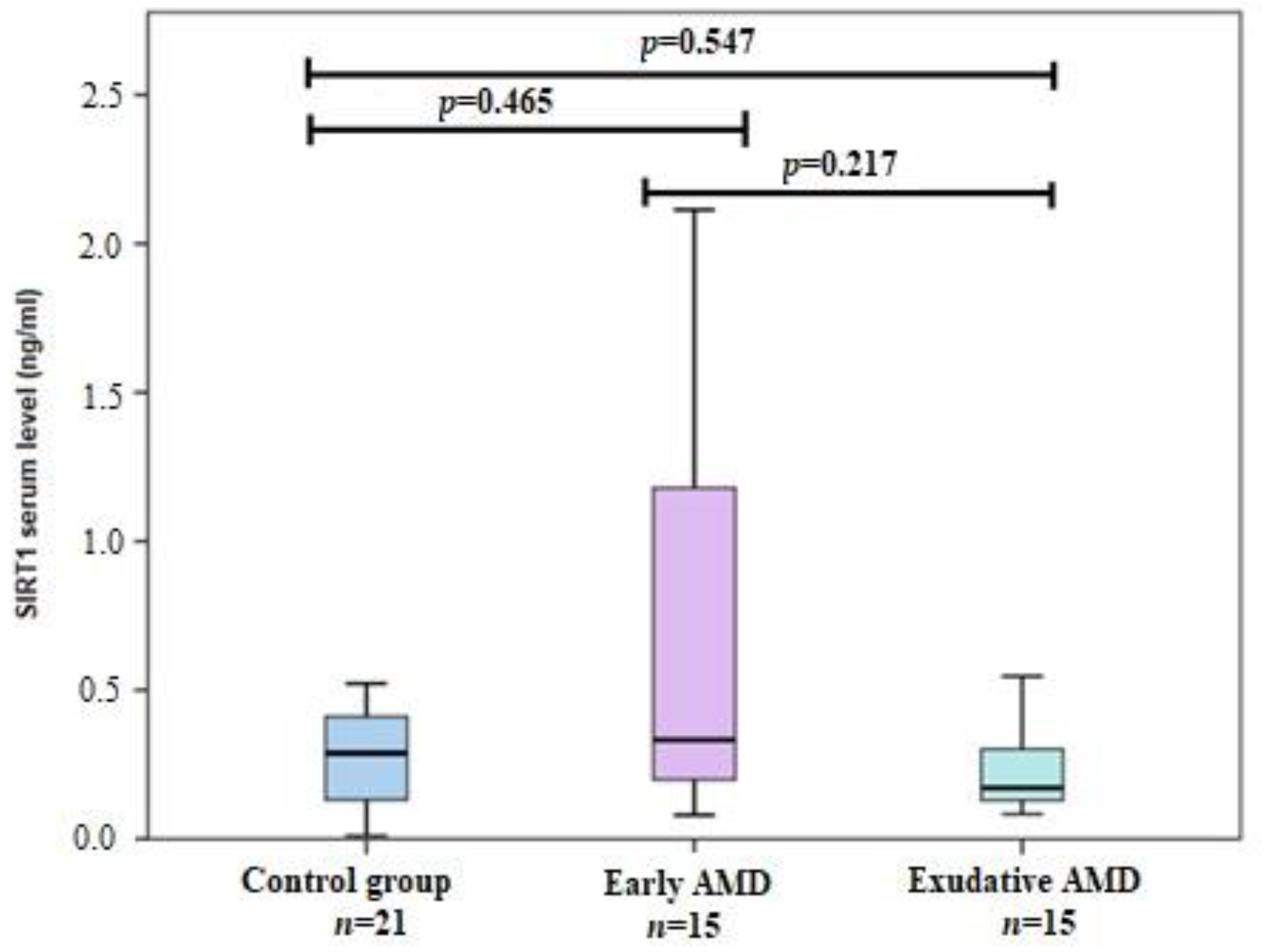
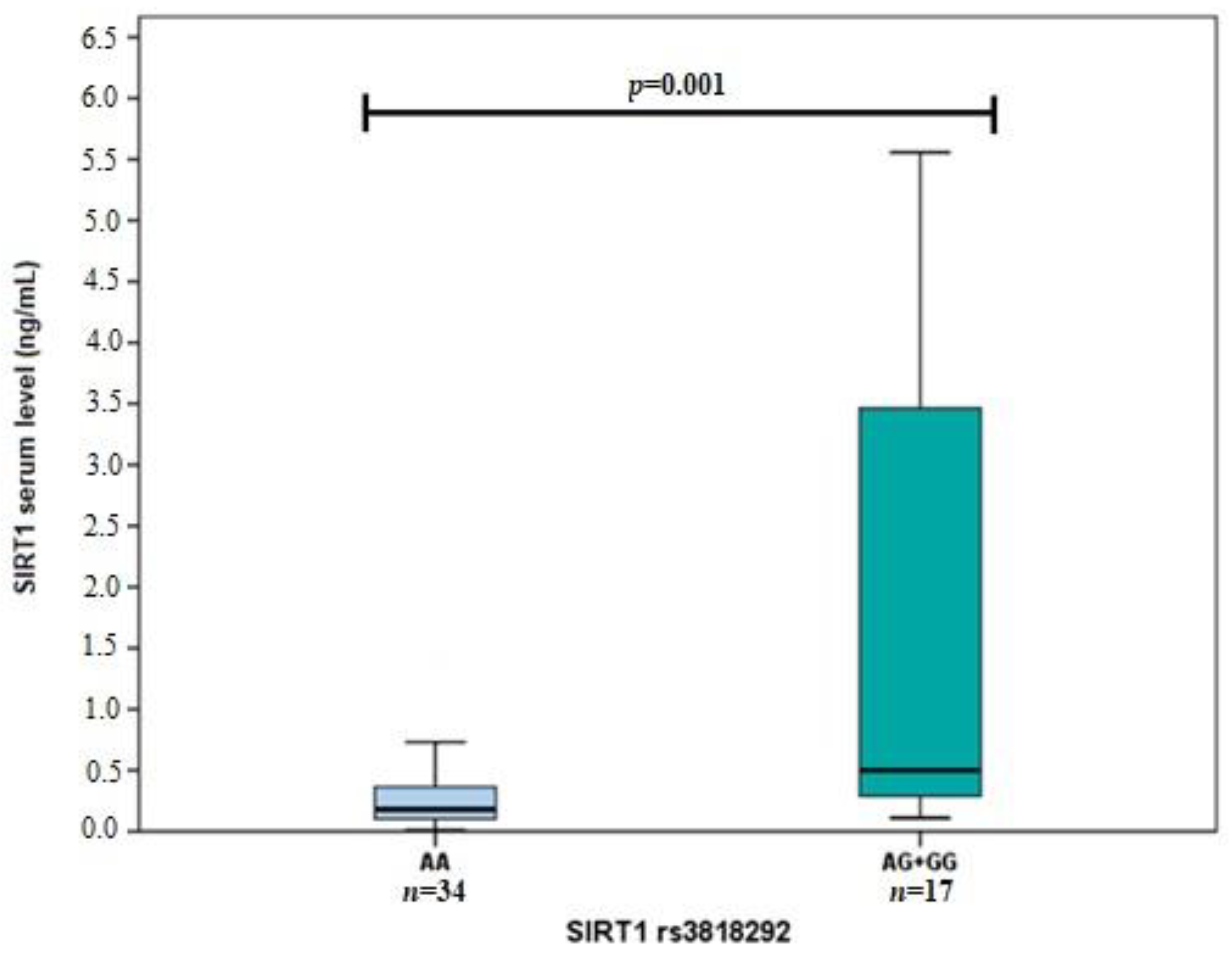
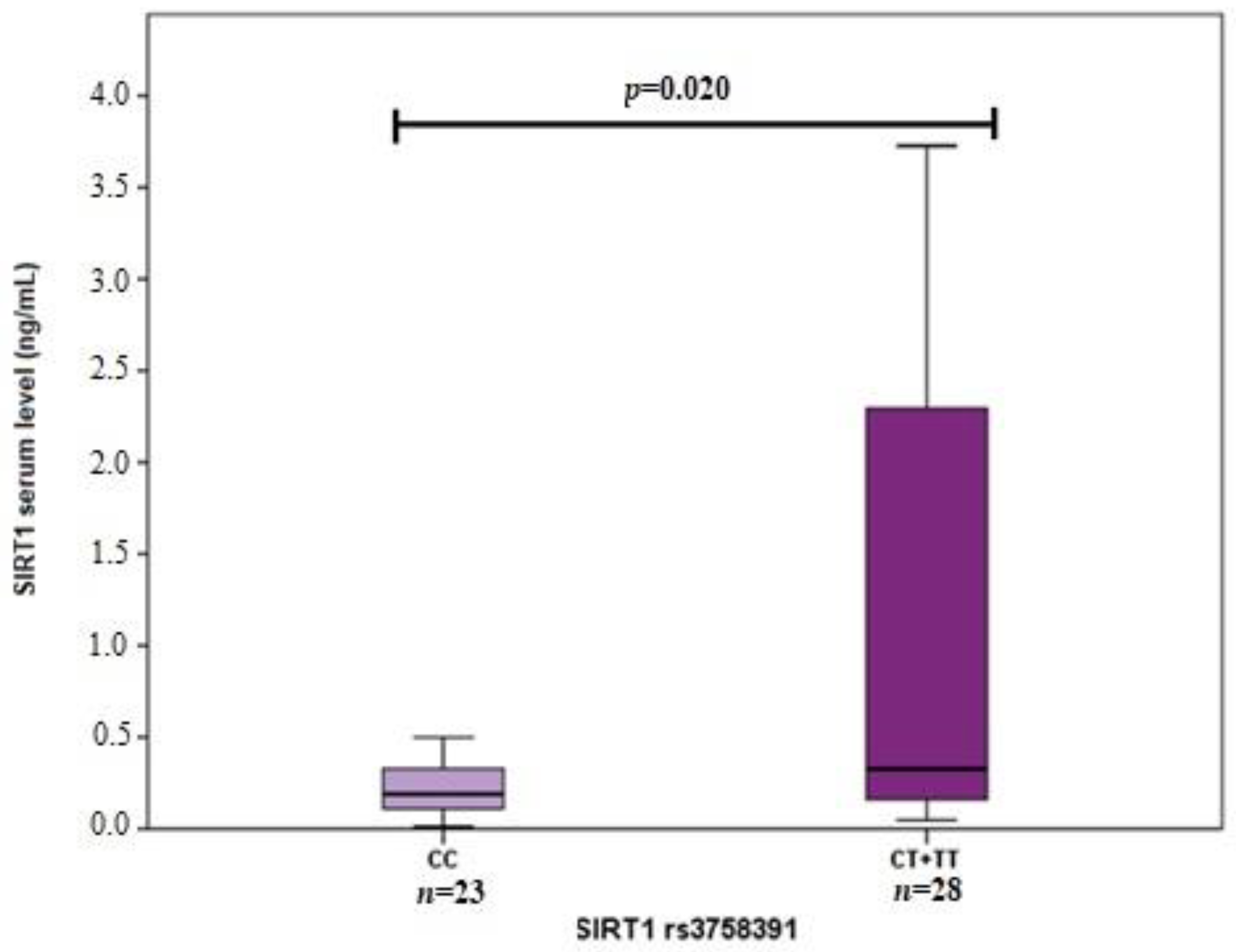
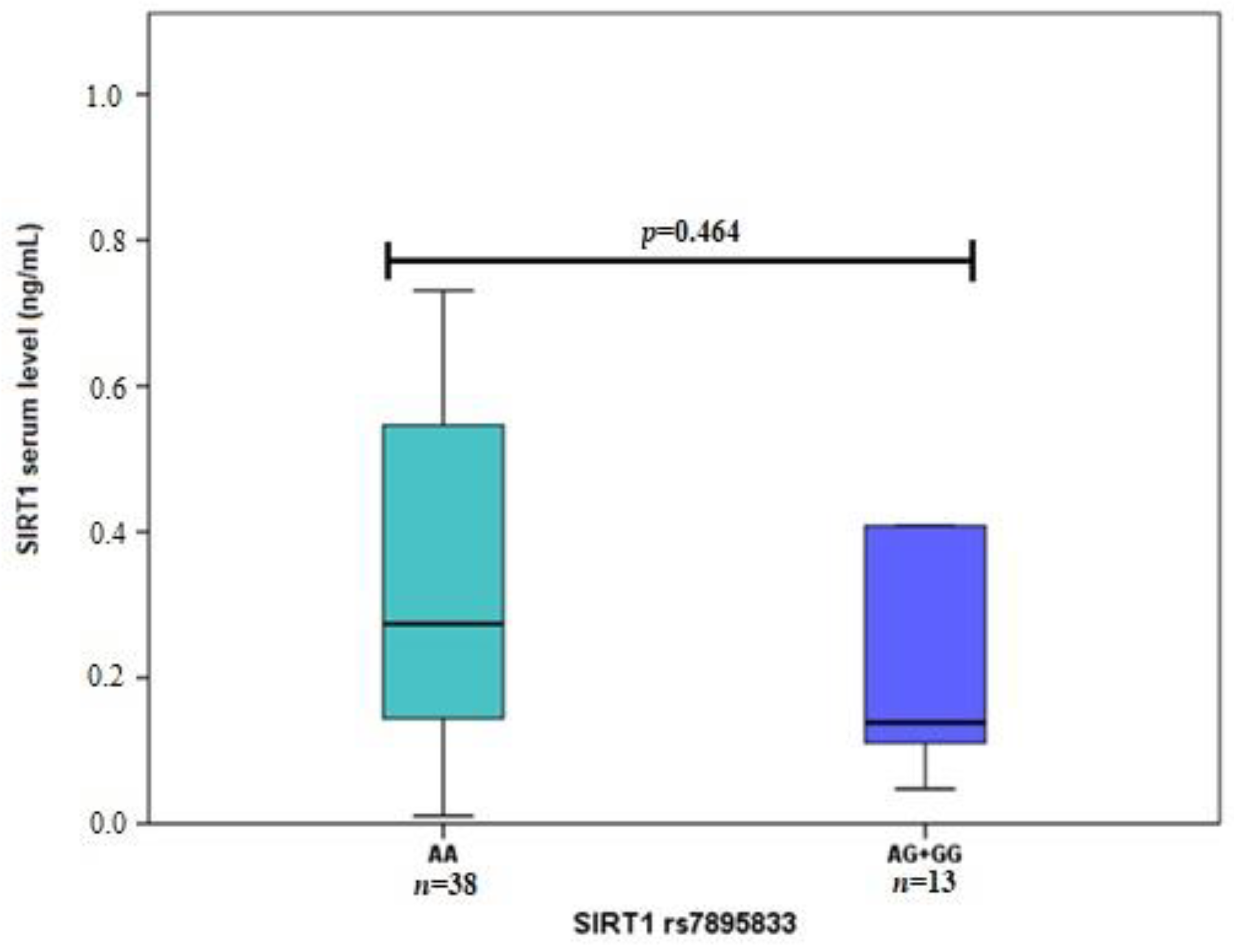
3.2. SIRT1 Serum Levels in Early and Exudative AMD and Controls
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klein, R.; Peto, T.; Bird, A.; VanNewkirk, M.R. The epidemiology of age-related macular degeneration. Am. J. Ophthalmol. 2004, 137, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Friedman, D.S.; O’Colmain, B.J.; Muñoz, B.; Tomany, S.C.; McCarty, C.; de Jong, P.T.; Barbara, N.; Paul, M.; John, K.; Congdon, N.; et al. Prevalence of age-related macular degeneration in the United States. Arch Ophthalmol. 2004, 122, 564–572. [Google Scholar] [PubMed] [Green Version]
- Neverauskiene, J.; Chaleckiene, G.; Baniuliene, D.; Kalasauskiene, A. Blindness incidence in Lithuania. Int. J. Ophthalmol. 2003, 3, 225–228. [Google Scholar]
- The Age-Related Eye Disease Study system for classifying age-related macular degeneration from stereoscopic color fundus photographs: The Age-Related Eye Disease Study Report Number 6. Am. J. Ophthalmol. 2011, 132, 668–681.
- Zarbin, M.A. Current Concepts in the Pathogenesis of Age-Related Macular Degeneration. Arch. Ophthalmol. 2004, 122, 598–614. [Google Scholar] [CrossRef] [Green Version]
- Maeda, S.; Koya, D.; Araki, S.-I.; Babazono, T.; Umezono, T.; Toyoda, M.; Kawai, K.; Imanishi, M.; Uzu, T.; Suzuki, D.; et al. Association between single nucleotide polymorphisms within genes encoding sirtuin families and diabetic nephropathy in Japanese subjects with type 2 diabetes. Clin. Exp. Nephrol. 2011, 15, 381–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brochier, C.; Dennis, G.; Rivieccio, M.A.; McLaughlin, K.; Coppola, G.; Ratan, R.R.; Langley, B. Specific acetylation of p53 by HDAC inhibition prevents DNA damage-induced apoptosis in neurons. J. Neurosci. 2013, 33, 8621–8632. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; McVey, M.; Guarente, L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999, 13, 2570–2580. [Google Scholar] [CrossRef] [Green Version]
- Wojcik, M.; Mac-Marcjanek, K.; Wozniak, L.A. Physiological and pathophysiological functions of SIRT1. Mini-Rev. Med. Chem. 2009, 9, 386–394. [Google Scholar] [CrossRef]
- Han, J.; Hubbard, B.P.; Lee, J.; Montagna, C.; Lee, H.W.; Sinclair, D.A.; Suh, Y. Analysis of 41 cancer cell lines reveals excessive allelic loss and novel mutations in the SIRT1 gene. Cell Cycle 2013, 12, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, Y.; Kubota, S.; Narimatsu, T.; Yuki, K.; Koto, T.; Sasaki, M.; Tsubota, K. Retinal aging and sirtuins. Ophthalm. Res. 2010, 44, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Mimura, T.; Kaji, Y.; Noma, H.; Funatsu, H.; Okamoto, S. The role of SIRT1 in ocular aging. Exp. Eye Res. 2013, 116, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kong, X.; Martins-Santos, M.E.S.; Aleman, G.; Chaco, E.; Liu, G.E.; Wu, S.-Y.; Samols, D.; Hakimi, P.; Chiang, C.-M.; et al. Activation of SIRT1 by resveratrol represses transcription of the gene for the cytosolic form of phosphoenolpyruvate carboxykinase (GTP) by deacetylating hepatic nuclear factor 4α. J. Biol. Chem. 2009, 284, 27042–27053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidorova-Darmos, E.; Wither, R.G.; Shulyakova, N.; Fisher, C.; Ratnam, M.; Aarts, M.; Lilge, L.; Monnier, P.P.; Eubanks, J.H. Differential expression of sirtuin family members in the developing, adult, and aged rat brain. Front. Aging Neurosci. 2014, 6, 333. [Google Scholar] [CrossRef]
- Ames, A.; Li, Y.-Y. Energy requirements of glutamatergic pathways in rabbit retina. J. Neurosci. 1992, 12, 4234–4242. [Google Scholar] [CrossRef] [Green Version]
- Niven, J.E.; Laughlin, S.B. Energy limitation as a selective pressure on the evolution of sensory systems. J. Exp. Biol. 2008, 211, 1792–1804. [Google Scholar] [CrossRef] [Green Version]
- Ban, N.; Ozawa, Y.; Inaba, T.; Miyake, S.; Watanabe, M.; Shinmura, K.; Tsubota, K. Light-dark condition regulates sirtuin mRNA levels in the retina. Exp. Gerontol. 2013, 48, 1212–1217. [Google Scholar] [CrossRef] [Green Version]
- Potente, M.; Ghaeni, L.; Baldessari, D.; Mostoslavsky, R.; Rossig, L.; Dequiedt, F.; Haendeler, J.; Mione, M.; Dejana, E.; Alt, F.W.; et al. SIRT1 controls endothelial angiogenic functions during vascular growth. Genes Dev. 2007, 21, 2644–2658. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Damico, L.; Shams, N.; Lowman, H.; Kim, R. Development of ranibizumab, an anti-vascular endothelial growth factor antigen binding fragment, as therapy for neovascular age-related macular degeneration. Retina 2006, 26, 859–870. [Google Scholar] [CrossRef]
- Liutkevičienė, R.; Lesauskaitė, V.; Ašmonienė, V.; Žaliūnienė, D.; Jašinskas, V. Factors determining age-related macular degeneration: A current view. Medicina 2010, 46, 89–94. [Google Scholar] [CrossRef]
- rs3818292 RefSNP Report-dbSNP-NCBI n.d. Available online: https://www.ncbi.nlm.nih.gov/snp/rs3818292 (accessed on 22 April 2022).
- Lin, H.; Hargreaves, K.A.; Li, R.; Reiter, J.L.; Wang, Y.; Mort, M.; Cooper, D.N.; Zhou, Y.; Zhang, C.; Eadon, M.T.; et al. RegSNPs-intron: A computational framework for predicting pathogenic impact of intronic single nucleotide variants. Genome Biol. 2019, 20, 254. [Google Scholar] [CrossRef] [PubMed]
- Tao, T.T.; Lin, X.H.; Tang, S.J.; Gui, W.W.; Zhu, W.F.; Li, H. Association of genetic variants in the Sirt1 and Nrf2 genes with the risk of metabolic syndrome in a Chinese Han population. BMC Endocr. Disord. 2022, 22, 84. [Google Scholar] [CrossRef] [PubMed]
- Promoter n.d. Available online: https://www.genome.gov/genetics-glossary/Promoter (accessed on 22 April 2022).
- rs3758391 RefSNP Report-dbSNP-NCBI n.d. Available online: https://www.ncbi.nlm.nih.gov/snp/rs3758391 (accessed on 22 April 2022).
- rs7895833 RefSNP Report-dbSNP-NCBI n.d. Available online: https://www.ncbi.nlm.nih.gov/snp/rs7895833 (accessed on 22 April 2022).
- Liutkeviciene, R.; Vilkeviciute, A.; Streleckiene, G.; Kriauciuniene, L.; Chaleckis, R.; Deltuva, V.P. Associations of cholesteryl ester transfer protein (CETP) gene variants with predisposition to age-related macular degeneration. Gene 2017, 636, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaiya, S.; Murthy, R.K.; Chalam, K.V. Resveratrol inhibits proliferation of hypoxic choroidal vascular endothelial cells. Mol. Vis. 2013, 19, 2385–2392. [Google Scholar]
- Fletcher, A.E. Free radicals, antioxidants and eye diseases: Evidence from epidemiological studies on cataract and age-related macular degeneration. Ophthalm. Res. 2010, 44, 191–198. [Google Scholar] [CrossRef]
- Winkler, B.S.; Boulton, M.E.; Gottsch, J.D.; Sternberg, P. Oxidative damage and age-related macular degeneration. Mol. Vis. 1999, 5, 32. [Google Scholar]
- Zuo, L.; Khan, R.S.; Lee, V.; Dine, K.; Wu, W.; Shindler, K.S. SIRT1 promotes RGC survival and delays loss of function following optic nerve crush. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5097–5102. [Google Scholar] [CrossRef]
- Lee, S.H.; Schloss, D.J.; Swain, J.L. Maintenance of Vascular Integrity in the Embryo Requires Signaling through the Fibroblast Growth Factor Receptor. J. Biol. Chem. 2000, 275, 33679–33687. [Google Scholar] [CrossRef] [Green Version]
- Lesauskaitė, V.; Šinkūnaitė, G.; Benetis, R.; Grabauskas, V.; Vaškelytė, J.; Smalinskienė, A.; Šimonytė, S.; Jarienė, G.; Tatarūnas, V.; Klumbienė, J.; et al. Matrix metalloproteinase-3 gene polymorphism and dilatative pathology of ascending thoracic aorta. Medicina 2008, 44, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Balaiya, S.; Khetpal, V.; Chalam, K.V. Hypoxia initiates sirtuin1-mediated vascular endothelial growth factor activation in choroidal endothelial cells through hypoxia inducible factor-2α. Mol. Vis. 2012, 18, 114–120. [Google Scholar] [PubMed]
- Bhattacharya, S.; Chaum, E.; Johnson, D.A.; Johnson, L.R. Age-Related Susceptibility to Apoptosis in Human Retinal Pigment Epithelial Cells Is Triggered by Disruption of p53–Mdm2 Association. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8350–8366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloney, S.C.; Antecka, E.; Granner, T.; Fernandes, B.; Lim, L.-A.; Orellana, M.E.; Burnier, M.N. Expression of sirt1 in choroidal neovascular membranes. Retina 2013, 33, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-L.; Mostoslavsky, R.; Saito, S.; Manis, J.P.; Gu, Y.; Patel, P.; Bronson, R.; Appella, E.; Alt, F.W.; Chua, K.F. Developmental defects and p53 hyperacetylation in Sir2 homolog (SIRT1)-deficient mice. Proc. Natl. Acad. Sci. USA 2003, 100, 10794–10799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaliffa, C.; Ameqrane, I.; Dansault, A.; Leemput, J.; Vieira, V.; Lacassagne, E.; Provost, A.; Bigot, K.; Masson, C.; Menasche, M.; et al. Sirt1 involvement in rd10 mouse retinal degeneration. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3562–3572. [Google Scholar] [CrossRef] [Green Version]
- McBurney, M.W.; Yang, X.; Jardine, K.; Hixon, M.; Boekelheide, K.; Webb, J.R.; Lansdorp, P.M.; Lemieux, M. The mammalian SIR2α protein has a role in embryogenesis and gametogenesis. Mol. Cell. Biol. 2003, 23, 38–54. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Lu, Y. Changes in SIRT1 expression and its downstream pathways in age-related cataract in humans. Curr. Eye Res. 2011, 36, 449–455. [Google Scholar] [CrossRef]
- Lin, T.-J.; Peng, C.-H.; Chiou, S.-H.; Liu, J.H.; Tsai, C.Y.; Chuang, J.H.; Chen, S.J. Severity of lens opacity, age, and correlation of the level of silent information regulator T1 expression in age-related cataract. J. Cataract. Refract. Surg. 2011, 37, 1270–1274. [Google Scholar] [CrossRef]
- Anekonda, T.S.; Adamus, G. Resveratrol prevents antibody-induced apoptotic death of retinal cells through upregulation of Sirt1 and Ku70. BMC Res. Notes 2008, 1, 122. [Google Scholar] [CrossRef] [Green Version]
- Kurihara, T.; Kubota, Y.; Ozawa, Y.; Takubo, K.; Noda, K.; Simon, M.C.; Johnson, R.S.; Suematsu, M.; Tsubota, K.; Ishida, S.; et al. Von Hippel-Lindau protein regulates transition from the fetal to the adult circulatory system in retina. Development 2010, 137, 1563–1571. [Google Scholar] [CrossRef] [Green Version]
- Kubota, S.; Ozawa, Y.; Kurihara, T.; Sasaki, M.; Yuki, K.; Miyake, S.; Noda, K.; Ishida, S.; Tsubota, K. Roles of AMP-Activated Protein Kinase in Diabetes-Induced Retinal Inflammation. Investig. Ophthalmol. Vis. Sci. 2011, 52, 9142–9148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, S.; Kurihara, T.; Ebinuma, M.; Kubota, M.; Yuki, K.; Sasaki, M.; Noda, K.; Ozawa, Y.; Oike, Y.; Ishida, S.; et al. Resveratrol Prevents Light-Induced Retinal Degeneration via Suppressing Activator Protein-1 Activation. Am. J. Pathol. 2010, 177, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhai, Y.; Zhang, W.; Teng, Y.; Yao, K. Single Nucleotide Polymorphisms of the Sirtuin 1 (SIRT1) Gene are Associated With age-Related Macular Degeneration in Chinese Han Individuals. Medicine 2015, 94, e2238. [Google Scholar] [CrossRef] [PubMed]
- Razi, S.; Cogger, V.C.; Kennerson, M.; Benson, V.L.; McMahon, A.C.; Blyth, F.M.; Handelsman, D.J.; Seibel, M.J.; Hirani, V.; Naganathan, V.; et al. SIRT1 Polymorphisms and Serum-Induced SIRT1 Protein Expression in Aging and Frailty: The CHAMP Study. J. Gerontol. Ser. A 2017, 72, 870–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liutkeviciene, R.; Vilkeviciute, A.; Kriauciuniene, L.; Deltuva, V.P. SIRT1 rs12778366, FGFR2 rs2981582, STAT3 rs744166, LIPC rs10468017, rs493258 and LPL rs12678919 genotypes and haplotype evaluation in patients with age-related macular degeneration. Gene 2018, 686, 8–15. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Polymorphisms | Position | Chromosome’s Location | Molecular Significance |
|---|---|---|---|
| SIRT1 rs3818292 | Chr10:67907144 [21] | Intragenic region [21] | Alteration of gene-splicing processes [22] |
| SIRT1 rs3758391 | Chr10:67883584 [25] | Promoter region [23] | Initiation of gene transcription [24] |
| SIRT1 rs7895833 | Chr10:67863299 [26] | Promoter region [23] | Initiation of a gene transcription [24] |
| Characteristic | Group | p-Value | ||
|---|---|---|---|---|
| Early AMD n = 308 n (%) | Exudative AMD n = 411 n (%) | Control Group n = 225 n (%) | ||
| Gender | ||||
| Males | 97 (31.5) | 145 (35.3) | 89 (39.6) | 0.054 * |
| Females | 211 (68.5) | 266 (64.7) | 136 (60.4) | 0.285 ** |
| Age, (years), median (IQR) | 74 (12) | 77 (10) | 74 (7) | 0.052 * 0.008 ** |
| Genotype/Allele | Early AMD n = 308 n (%) | Exudative AMD n = 411 n (%) | Control Group n = 225 n (%) | HWE p-Value | p-Value |
|---|---|---|---|---|---|
| SIRT1 rs3818292 | |||||
| AA | 263 (85.4) | 336 (81.8) | 200 (88.9) | 0.378 | 0.291 * |
| AG | 43 (14) | 72 (17.5) | 25 (11.1) | 0.040 ** | |
| GG | 2 (0.6) | 3 (0.7) | 0 (0) | 0.430 *** | |
| Total | 308 (100) | 411 (100) | 225 (100) | 0.183 * | |
| A | 569 (92.4) | 744 (90.5) | 425 (94.4) | 0.014 ** | |
| G | 47 (7.6) | 78 (9.5) | 25 (5.6) | 0.216 *** | |
| SIRT1 rs3758391 | |||||
| CC | 163 (52.9) | 202 (49.1) | 126 (56.0) | 0.148 | 0.631 * |
| CT | 128 (41.6) | 178 (43.3) | 90 (40.0) | 0.100 ** | |
| CC | 17 (5.5) | 31 (7.5) | 9 (4.0) | 0.427 *** | |
| Total | 308 (100) | 411 (100) | 225 (100) | 0.394 * | |
| C | 454 (73.7) | 582 (70.8) | 342 (76) | 0.047 ** | |
| T | 162 (26.3) | 240 (29.2) | 108 (24) | 0.226 *** | |
| SIRT1 rs7895833 | |||||
| AA | 231 (75.0) | 275 (66.9) | 169 (75.1) | 0.302 | 0.905 * |
| AG | 73 (23.7) | 120 (29.2) | 54 (24.0) | 0.024 ** | |
| GG | 4 (1.3) | 16 (3.9) | 2 (0.9) | 0.019 *** | |
| Total | 308 (100) | 411 (100) | 225 (100) | 0.007 * | |
| A | 535 (86.9) | 670 (81.5) | 392 (87.1) | 0.010 ** | |
| G | 81 (13.1) | 152 (18.5) | 58 (12.9) | 0.007 *** | |
| Model | Genotype/Allele | OR (95% CI) * | p-Value | AIC |
|---|---|---|---|---|
| Exudative AMD | ||||
| SIRT1 rs3818292 | ||||
| Codominant | AG vs. AA GG vs. AA | 1.750 (1.073–2.855) - | 0.025 - | 820.322 |
| Dominant | AG+GG vs. AA | 1.823 (1.120–2.966) | 0.016 | 820.047 |
| Recessive | GG vs. AG+AA | - | - | - |
| Overdominant | AG vs. AA+GG | 1.735 (1.064–2.829) | 0.027 | 821.113 |
| Additive | G | 1.845 (1.148–2.966) | 0.011 | 819.337 |
| SIRT1 rs3758391 | ||||
| Co-dominant | CT vs. CC TT vs. CC | 1.270 (0.904–1.783) 2.226 (1.039–4.942) | 0.169 0.040 | 822.709 |
| Dominant | CT+TT vs. CC | 1.358 (0.977–1.888) | 0.069 | 822.956 |
| Recessive | TT vs. CT+CC | 2.031 (0.946–4.361) | 0.069 | 822.614 |
| Overdominant | CT vs. CC+TT | 1.170 (0.839–1.631) | 0.355 | 825.424 |
| Additive | T | 1.367 (1.037–1.800) | 0.026 | 821.238 |
| SIRT1 rs7895833 | ||||
| Co-dominant | AG vs. AA GG vs. AA | 1.405 (0.964–2.047) 5.246 (1.187–23.196) | 0.077 0.029 | 818.971 |
| Dominant | AG+GG vs. AA | 1.538 (1.064–2.223) | 0.022 | 820.890 |
| Recessive | GG vs. AG+AA | 4.756 (1.080–20.939) | 0.039 | 820.161 |
| Overdominant | AG vs. AA+GG | 1.336 (0.918–1.943) | 0.130 | 823.946 |
| Additive | G | 1.577 (1.133–2.196) | 0.007 | 818.614 |
| Genotype/Allele | Early AMD n = 308 n (%) | Exudative AMD n = 411 n (%) | Control Group n = 225 n (%) | p-Value |
|---|---|---|---|---|
| Females | ||||
| SIRT1 rs3818292 | ||||
| AA | 186 (88.2) | 221 (83.1) | 119 (87.5) | 0.692 * |
| AG | 24 (11.4) | 42 (15.8) | 17 (12.5) | 0.300 ** |
| GG | 1 (0.5) | 3 (1.1) | 0 (0.0) | 0.271 *** |
| Total | 211 (100) | 266 (100) | 136 (100) | 0.629 * |
| A | 402 (93.9) | 484 (91) | 225 (93) | 0.353 ** |
| G | 26 (6.1) | 48 (9) | 17 (7) | 0.089 *** |
| SIRT1 rs3758391 | ||||
| CC | 112 (53.1) | 128 (48.1) | 75 (55.1) | 0.677 * |
| CT | 90 (42.7) | 115 (43,2) | 53 (39.0) | 0.339 ** |
| CC | 9 (4.3) | 23 (8,6) | 8 (5.9) | 0.139 *** |
| Total | 211 (100) | 411 (100) | 136 (100) | 0.947 * |
| 314 (74.4) | 371 (69.7) | 203 (74.6) | 0.146 ** | |
| C | 108 (25.6) | 161 (30.3) | 69 (25.4) | <0.001 *** |
| SIRT1 rs7895833 | ||||
| AA | 162 (76.8) | 177 (66.5) | 105 (77.2) | 0.977 * |
| AG | 47 (22.3) | 78 (29.3) | 30 (22.1) | 0.977 ** |
| GG | 2 (0.9) | 11 (4.1) | 1 (0.7) | 0.015 *** |
| Total | 211 (100) | 411 (100) | 136 (100) | 0.899 * |
| A | 371 (87.9) | 432 (81.2) | 240 (88.2) | 0.011 ** |
| G | 51 (12.1) | 100 (18.8) | 32 (11.8) | 0.005 *** |
| Males | ||||
| SIRT1 rs3818292 | ||||
| AA | 77 (79.4) | 115 (79.3) | 81 (91.0) | 0.072 * |
| AG | 19 (19.6) | 30 (20.7) | 8 (9.0) | 0.072 ** |
| GG | 1 (1.0) | 1 (1.0) | 0 (0.0) | 0.465 *** |
| Total | 97 (100) | 146 (100) | 89 (100) | 0.023 * |
| A | 173 (89.2) | 230 (87.8) | 170 (95.5) | 0.006 ** |
| G | 21 (10.8) | 32 (12.2) | 8 (4.5) | 0.647 *** |
| SIRT1 rs3758391 | ||||
| CC | 51 (57.3) | 74 (51.0) | 51 (57.3) | 0.077 * |
| CT | 37 (41.6) | 63 (43.4) | 37 (41.6) | 0.077 ** |
| CC | 1 (1.1) | 8 (5.5) | 1 (1.1) | 0.626 *** |
| Total | 97 (100) | 146 (100) | 89 (100) | 1.000 * |
| C | 139 (78.1) | 211 (72.8) | 139 (78.1) | 0.197 ** |
| T | 39 (21.9) | 79 (27.2) | 39 (21.9) | 0.197 *** |
| SIRT1 rs7895833 | ||||
| AA | 64 (71.9) | 98 (67.7) | 64 (71.9) | 0.879 * |
| AG | 24 (27.0) | 42 (29.0) | 24 (27.0) | 0.879 ** |
| GG | 1 (1.1) | 5 (3.4) | 1 (1.1) | 0.745 *** |
| Total | 97 (100) | 146 (100) | 89 (100) | 1.000 * |
| A | 152 (85.4) | 238 (82.1) | 152 (85.4) | 0.349 ** |
| G | 26 (14.6) | 52 (17.9) | 26 (14.6) | 0.349 *** |
| Model | Genotype/Allele | OR (95% CI) * | p-Value | AIC |
|---|---|---|---|---|
| Exudative AMD | ||||
| SIRT1 rs3818292 | ||||
| Codominant | AG vs. AA GG vs. AA | 1.349 (0.735–2.479) - | 0.334 - | 514.698 |
| Dominant | AG+GG vs. AA | 1.446 (0.791–2.642) | 0.231 | 514.665 |
| Recessive | GG vs. AG+AA | - | - | - |
| Overdominant | AG vs. AA+GG | 1.331 (0.725–2.444) | 0.356 | 515.282 |
| Additive | G | 1.502 (0.846–2.667) | 0.165 | 514.121 |
| SIRT1 rs3758391 | ||||
| Co-dominant | CT vs. CC TT vs. CC | 1.282 (0.831–1.977) 1.742 (0.738–4.114) | 0.262 0.206 | 515.754 |
| Dominant | CT+TT vs. CC | 1.340 (0.884–2.033) | 0.168 | 514.247 |
| Recessive | TT vs. CT+CC | 1.556 (0.674–3.595) | 0.300 | 515.021 |
| Overdominant | CT vs. CC+TT | 1.196 (0.785–1.824) | 0.405 | 515.460 |
| Additive | T | 1.301 (0.929–1.822) | 0.126 | 513.766 |
| SIRT1 rs7895833 | ||||
| Co-dominant | AG vs. AA GG vs. AA | 1.563 (0.964–2.047) 6.878 (0.872–547.279) | 0.072 0.067 | 510.122 |
| Dominant | AG+GG vs. AA | 1.729 (1.073–2.786) | 0.024 | 510.884 |
| Recessive | GG vs. AG+AA | 6.072 (0.773–47.709) | 0.086 | 511.457 |
| Overdominant | AG vs. AA+GG | 1.478 (0.910–2.400) | 0.114 | 513.589 |
| Additive | G | 1.753 (1.140–2.697) | 0.011 | 509.123 |
| Model | Genotype/Allele | OR (95% CI) * | p-Value | AIC |
|---|---|---|---|---|
| Early AMD | ||||
| SIRT1 rs3818292 | ||||
| Codominant | AG vs. AA GG vs. AA | 2.498 (1.033–6.041) - | 0.042 - | 255.749 |
| Dominant | AG+GG vs. AA | 2.630 (1.094–6.323) | 0.031 | 254.436 |
| Recessive | GG vs. AG+AA | - | - | - |
| Overdominant | AG vs. AA+GG | 1.298 (0.767–2.197) | 0.331 | 255.180 |
| Additive | G | 1.411 (0.851–2.338) | 0.182 | 254.046 |
| SIRT1 rs3758391 | ||||
| Co-dominant | CT vs. CC TT vs. CC | 1.027 (0.566–1.864) 8.000 (0.965–66.302) | 0.350 0.054 | 255.640 |
| Dominant | CT+TT vs. CC | 1.211 (0.678–2.160) | 0.518 | 259.088 |
| Recessive | TT vs. CT+CC | 7.910 (0.969–64.572) | 0.054 | 253.647 |
| Overdominant | CT vs. CC+TT | 0.905 (0.504–1.627) | 0.739 | 259.396 |
| Additive | T | 1.413 (0.860–2.321) | 0.172 | 257.615 |
| SIRT1 rs7895833 | ||||
| Co-dominant | AG vs. AA GG vs. AA | 1.005 (0.524–1.926) 1.855 (0.164–20.955) | 0.988 0.617 | 261.243 |
| Dominant | AG+GG vs. AA | 1.039 (0.549–1.965) | 0.907 | 259.493 |
| Recessive | GG vs. AG+AA | 1.853 (0.165–20.791) | 0.617 | 259.243 |
| Overdominant | AG vs. AA+GG | 0.992 (0.518–1.898) | 0.980 | 259.506 |
| Additive | G | 1.073 (0.598–1.926) | 0.813 | 259.450 |
| Exudative AMD | ||||
| SIRT1 rs3818292 | ||||
| Codominant | AG vs. AA GG vs. AA | 2.667 (1.159–6.142) - | 0.021 - | 306.335 |
| Dominant | AG+GG vs. AA | 2.667 (1.159–6.142) | 0.021 | 304.335 |
| Recessive | GG vs. AG+AA | - | - | - |
| Overdominant | AG vs. AA+GG | 2.667 (1.159–6.142) | 0.021 | 304.335 |
| Additive | G | 2.667 (1.159–6.142) | 0.021 | 304.335 |
| SIRT1 rs3758391 | ||||
| Co-dominant | CT vs. CC TT vs. CC | 1.283 (0.737–2.234) 5.771 (0.694–47.986) | 0.378 0.105 | 308.174 |
| Dominant | CT+TT vs. CC | 1.407 (0.816–2.426) | 0.220 | 308.841 |
| Recessive | TT vs. CT+CC | 5.158 (0.630–42.206) | 0.126 | 306.956 |
| Overdominant | CT vs. CC+TT | 1.173 (0.679–2.028) | 0.567 | 310.030 |
| Additive | T | 1.501 (0.923–2.441) | 0.102 | 307.606 |
| SIRT1 rs7895833 | ||||
| Co-dominant | AG vs. AA GG vs. AA | 1.210 (0.664–2.205) 3.382 (0.382–29.988) | 0.533 0.274 | 310.600 |
| Dominant | AG+GG vs. AA | 1.298 (0.722–2.333) | 0.383 | 309.590 |
| Recessive | GG vs. AG+AA | 3.197 (0.364–28.116) | 0.295 | 308.991 |
| Overdominant | AG vs. AA+GG | 1.167 (0.642–2.121) | 0.612 | 310.101 |
| Additive | G | 1.344 (0.794–2.275) | 0.271 | 309.117 |
| SNPs | Early AMD vs. Controls | Exudative AMD vs. Controls | ||||
|---|---|---|---|---|---|---|
| D’ | r2 | p-Value | D’ | r2 | p-Value | |
| rs3818292–rs3758391 | 0.9302 | 0.1848 | 0.0 | 0.9321 | 0.2032 | 0.0 |
| rs3818292–rs7895833 | 0.9142 | 0.4037 | 0.0 | 0.9235 | 0.3799 | 0.0 |
| rs3758391–rs7895833 | 0.9329 | 0.3848 | 0.0 | 0.6978 | 0.4869 | 0.0 |
| Haplotype | rs3818292 | rs3758391 | rs7895833 | Frequency (%) | OR (95% CI) | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Early AMD | Controls | Total | ||||||
| 1 | A | C | A | 73.17 | 74.44 | 73.65 | 1.00 | --- |
| 2 | A | T | A | 13.34 | 11.96 | 12.8 | 1.14 (0.77–1.69) | 0.51 |
| 3 | G | T | G | 7.29 | 4.8 | 6.25 | 1.51 (0.78–1.50) | 0.13 |
| 4 | A | T | G | 5.67 | 6.78 | 6.13 | 0.85 (0.51–1.42) | 0.53 |
| rare | * | * | * | NA | NA | 1.15 | 0.28 (0.07–1.09) | 0.066 |
| Haplotype | rs3818292 | rs3758391 | rs7895833 | Frequency (%) | OR (95% CI) | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Exudative AMD | Controls | Total | ||||||
| 1 | A | C | A | 70.4 | 74.44 | 71.79 | 1.00 | --- |
| 2 | A | T | A | 10.71 | 11.96 | 11.19 | 0.98 (0.66–1.45) | 0.92 |
| 3 | A | T | G | 9.41 | 16.78 | 8.49 | 1.45 (0.93–2.27) | 0.1 |
| 4 | G | T | G | 9.08 | 4.84 | 7.58 | 2.05 (1.23–3.43) | 0.0062 |
| rare | * | * | * | NA | NA | 0.96 | 0.23 (0.06–0.89) | 0.033 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaikaryte, K.; Gedvilaite, G.; Vilkeviciute, A.; Kriauciuniene, L.; Mockute, R.; Cebatoriene, D.; Zemaitiene, R.; Balciuniene, V.J.; Liutkeviciene, R. SIRT1: Genetic Variants and Serum Levels in Age-Related Macular Degeneration. Life 2022, 12, 753. https://doi.org/10.3390/life12050753
Kaikaryte K, Gedvilaite G, Vilkeviciute A, Kriauciuniene L, Mockute R, Cebatoriene D, Zemaitiene R, Balciuniene VJ, Liutkeviciene R. SIRT1: Genetic Variants and Serum Levels in Age-Related Macular Degeneration. Life. 2022; 12(5):753. https://doi.org/10.3390/life12050753
Chicago/Turabian StyleKaikaryte, Kriste, Greta Gedvilaite, Alvita Vilkeviciute, Loresa Kriauciuniene, Ruta Mockute, Dzastina Cebatoriene, Reda Zemaitiene, Vilma Jurate Balciuniene, and Rasa Liutkeviciene. 2022. "SIRT1: Genetic Variants and Serum Levels in Age-Related Macular Degeneration" Life 12, no. 5: 753. https://doi.org/10.3390/life12050753
APA StyleKaikaryte, K., Gedvilaite, G., Vilkeviciute, A., Kriauciuniene, L., Mockute, R., Cebatoriene, D., Zemaitiene, R., Balciuniene, V. J., & Liutkeviciene, R. (2022). SIRT1: Genetic Variants and Serum Levels in Age-Related Macular Degeneration. Life, 12(5), 753. https://doi.org/10.3390/life12050753