
The Protective Role of Neurogenetic Components in Reducing Stress-Related Effects during Spaceflights: Evidence from the Age-Related Positive Memory Approach

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Eligibility Criteria
2.2. Information Sources
2.3. Search Strategy
2.4. Data Collection
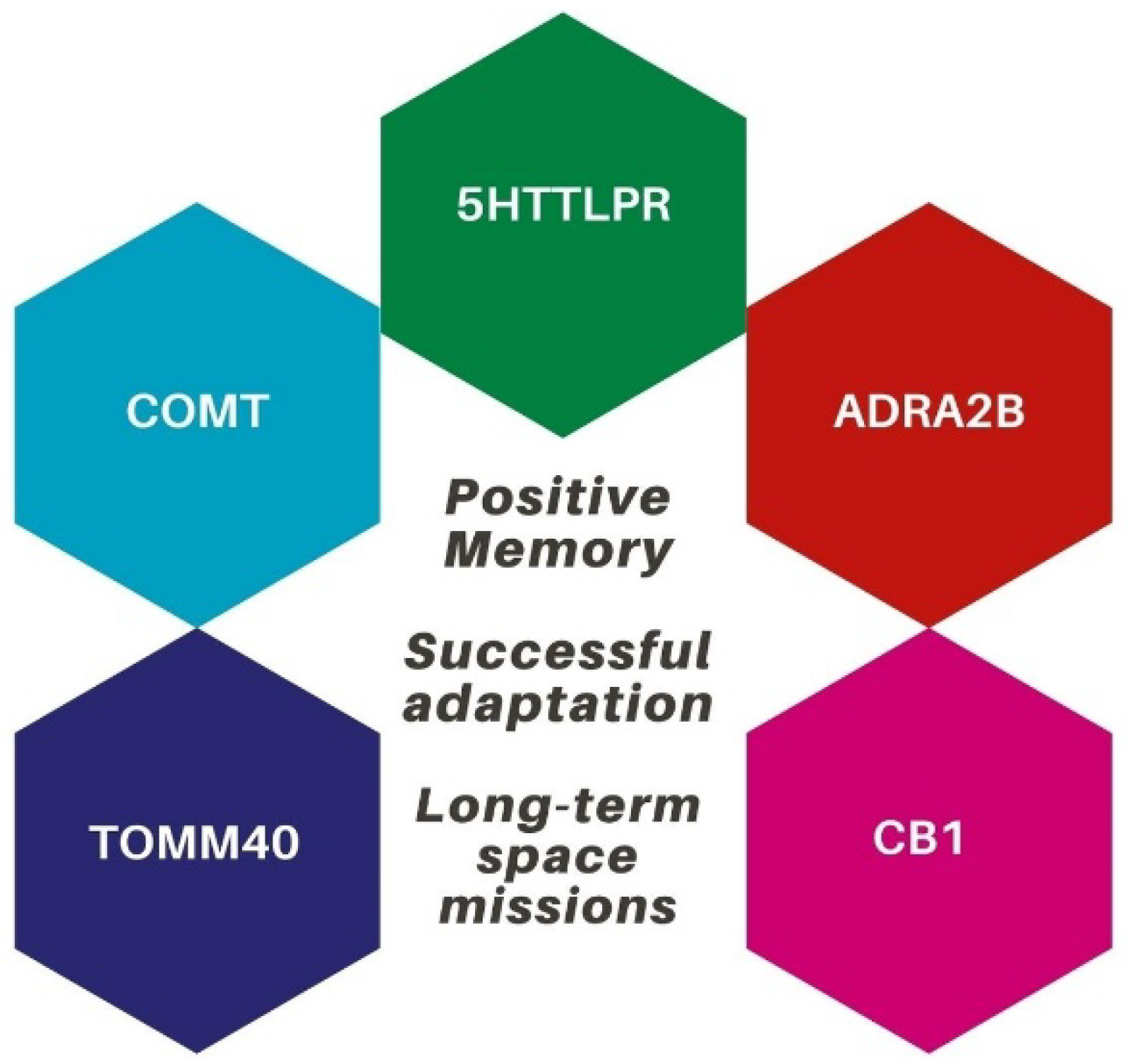
3. A Neurogenetic Profile for Sensitivity to Positive Memories: ADRA2B, COMT, 5HTTLPR, CB1, and TOMM40
- (i)
- A deletion variation of the ADRA2B gene, which affects adrenergic receptors’ activation;
- (ii)
- The Val/Met single-nucleotide polymorphism in the COMT gene, which affects dopamine metabolism in the prefrontal cortex;
- (iii)
- A short allele of the 5HTTLPR serotonin transporter gene;
- (iv)
- The endocannabinoid system, which involves cannabinoid receptors type 1 (CB1);
- (v)
- The translocase of outer mitochondria membrane 40 gene (TOMM40).
3.1. ADRA2B
3.2. COMT
3.3. HTTLPR
3.4. CB1
3.5. TOMM40
4. Discussion
4.1. A Positive Memory-Based Genetic Profile for Adaptation to the Space Environment
4.2. Hampering the Expression of the Positive Memory-Based Genetic Profile: The Case of Ionizing Radiations
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Strollo, F.; Gentile, S.; Strollo, G.; Mambro, A.; Vernikos, J. Recent Progress in Space Physiology and Aging. Front. Physiol. 2018, 9, 1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, A.D.; Mroczek, D.K.; Riffin, C. The Health Significance of Positive Emotions in Adulthood and Later Life. Soc. Personal. Psychol. Compass 2011, 5, 538–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pressman, S.D.; Cohen, S. Does Positive Affect Influence Health? Psychol. Bull. 2005, 131, 925–971. [Google Scholar] [CrossRef] [PubMed]
- Fredrickson, B.L. The Broaden-and-Build Theory of Positive Emotions. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Mather, M. The Affective Neuroscience of Aging. Annu. Rev. Psychol. 2016, 67, 213–238. [Google Scholar] [CrossRef] [Green Version]
- Mather, M.; Carstensen, L.L. Aging and Motivated Cognition: The Positivity Effect in Attention and Memory. Trends Cogn. Sci. 2005, 9, 496–502. [Google Scholar] [CrossRef]
- Mammarella, N.; Di Domenico, A.; Palumbo, R.; Fairfield, B. When Green Is Positive and Red Is Negative: Aging and the Influence of Color on Emotional Memories. Psychol. Aging 2016, 31, 914–926. [Google Scholar] [CrossRef] [Green Version]
- Mammarella, N.; Di Domenico, A.; Palumbo, R.; Fairfield, B. Self-generation and positivity effects following transcranial random noise stimulation in medial prefrontal cortex: A reality monitoring task in older adults. Cortex 2017, 91, 186–196. [Google Scholar] [CrossRef]
- Alfano, C.A.; Bower, J.L.; Cowie, J.; Lau, S.; Simpson, R.J. Long-Duration Space Exploration and Emotional Health: Recommendations for Conceptualizing and Evaluating Risk. Acta Astronaut. 2018, 142, 289–299. [Google Scholar] [CrossRef]
- Mammarella, N. Towards the Affective Cognition Approach to Human Performance in Space. Aerosp. Med. Hum. Perform. 2020, 91, 532–534. [Google Scholar] [CrossRef]
- Gatti, M.; Palumbo, R.; Di Domenico, A.; Mammarella, N. Affective Health and Countermeasures in Long-Duration Space Exploration. Heliyon 2022, 8, e09414. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan—A Web and Mobile App for Systematic Reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef] [Green Version]
- Todd, R.M.; Palombo, D.J.; Levine, B.; Anderson, A.K. Genetic Differences in Emotionally Enhanced Memory. Neuropsychologia 2011, 49, 734–744. [Google Scholar] [CrossRef]
- Mammarella, N.; Di Domenico, A.; Palumbo, R.; Fairfield, B. Noradrenergic Modulation of Emotional Memory in Aging. Ageing Res. Rev. 2016, 27, 61–66. [Google Scholar] [CrossRef]
- Mammarella, N.; Di Domenico, A.; Fairfield, B. Aging and the Genetic Road towards the Positivity Effect in Memory. Exp. Gerontol. 2016, 82, 120–124. [Google Scholar] [CrossRef]
- Mammarella, N.; Fairfield, B.; Di Domenico, A.; D’Onofrio, L.; Stuppia, L.; Gatta, V. The Modulating Role of ADRA2B in Emotional Working Memory: Attending the Negative but Remembering the Positive. Neurobiol. Learn. Mem. 2016, 130, 129–134. [Google Scholar] [CrossRef]
- Rasch, B.; Spalek, K.; Buholzer, S.; Luechinger, R.; Boesiger, P.; Papassotiropoulos, A.; de Quervain, D.J.-F. A Genetic Variation of the Noradrenergic System Is Related to Differential Amygdala Activation during Encoding of Emotional Memories. Proc. Natl. Acad. Sci. USA 2009, 106, 19191–19196. [Google Scholar] [CrossRef] [Green Version]
- Mather, M.; Harley, C.W. The Locus Coeruleus: Essential for Maintaining Cognitive Function and the Aging Brain. Trends Cogn. Sci. 2016, 20, 214–226. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Cappiello, M.; Meng, M.; Rosenthal, R.; Zhang, W. ADRA2B Deletion Variant and Enhanced Cognitive Processing of Emotional Information: A Meta-Analytical Review. Neurosci. Biobehav. Rev. 2018, 92, 402–416. [Google Scholar] [CrossRef]
- Li, S.; Weerda, R.; Milde, C.; Wolf, O.T.; Thiel, C.M. ADRA2B Genotype Differentially Modulates Stress-Induced Neural Activity in the Amygdala and Hippocampus during Emotional Memory Retrieval. Psychopharmacology 2015, 232, 755–764. [Google Scholar] [CrossRef]
- Urner, M.; van Wingen, G.; Franke, B.; Rijpkema, M.; Fernández, G.; Tendolkar, I. Genetic Variation of the A2b-Adrenoceptor Affects Neural Correlates of Successful Emotional Memory Formation. Hum. Brain Mapp. 2011, 32, 2096–2103. [Google Scholar] [CrossRef] [PubMed]
- Liberzon, I.; King, A.P.; Ressler, K.J.; Almli, L.M.; Zhang, P.; Ma, S.T.; Cohen, G.H.; Tamburrino, M.B.; Calabrese, J.R.; Galea, S. Interaction of the ADRB2 Gene Polymorphism With Childhood Trauma in Predicting Adult Symptoms of Posttraumatic Stress Disorder. JAMA Psychiatry 2014, 71, 1174–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mier, D.; Kirsch, P.; Meyer-Lindenberg, A. Neural Substrates of Pleiotropic Action of Genetic Variation in COMT: A Meta-Analysis. Mol. Psychiatry 2010, 15, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.M.; Gatt, J.M.; Grieve, S.M.; Dobson-Stone, C.; Paul, R.H.; Gordon, E.; Schofield, P.R. COMT Val(108/158)Met Polymorphism Effects on Emotional Brain Function and Negativity Bias. NeuroImage 2010, 53, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, P.M.; Lin, M.-K.; Sundararajan, R.; Fryxell, K.J.; Parasuraman, R. Healthy Aging Increases the Cognitive Effects of Two Genes That Influence Extracellular Dopamine. Psychol. Aging 2014, 29, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Papenberg, G.; Bäckman, L.; Nagel, I.E.; Nietfeld, W.; Schröder, J.; Bertram, L.; Heekeren, H.R.; Lindenberger, U.; Li, S.-C. COMT Polymorphism and Memory Dedifferentiation in Old Age. Psychol. Aging 2014, 29, 374–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, A.; Smolka, M.N. The Effects of Catechol O-Methyltransferase Genotype on Brain Activation Elicited by Affective Stimuli and Cognitive Tasks. Rev. Neurosci. 2006, 17, 359–367. [Google Scholar] [CrossRef]
- Mather, M.; Knight, M. Goal-Directed Memory: The Role of Cognitive Control in Older Adults’ Emotional Memory. Psychol. Aging 2005, 20, 554–570. [Google Scholar] [CrossRef] [Green Version]
- Ihne, J.L.; Gallagher, N.M.; Sullivan, M.; Callicott, J.H.; Green, A.E. Is Less Really More: Does a Prefrontal Efficiency Genotype Actually Confer Better Performance When Working Memory Becomes Difficult? Cortex J. Devoted Study Nerv. Syst. Behav. 2016, 74, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Swart, M.; Bruggeman, R.; Larøi, F.; Alizadeh, B.Z.; Kema, I.; Kortekaas, R.; Wiersma, D.; Aleman, A. COMT Val158Met Polymorphism, Verbalizing of Emotion and Activation of Affective Brain Systems. NeuroImage 2011, 55, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Miskowiak, K.W.; Kjaerstad, H.L.; Støttrup, M.M.; Svendsen, A.M.; Demant, K.M.; Hoeffding, L.K.; Werge, T.M.; Burdick, K.E.; Domschke, K.; Carvalho, A.F.; et al. The Catechol-O-Methyltransferase (COMT) Val158Met Genotype Modulates Working Memory-Related Dorsolateral Prefrontal Response and Performance in Bipolar Disorder. Bipolar Disord. 2017, 19, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Lelli-Chiesa, G.; Kempton, M.J.; Jogia, J.; Tatarelli, R.; Girardi, P.; Powell, J.; Collier, D.A.; Frangou, S. The Impact of the Val158Met Catechol-O-Methyltransferase Genotype on Neural Correlates of Sad Facial Affect Processing in Patients with Bipolar Disorder and Their Relatives. Psychol. Med. 2011, 41, 779–788. [Google Scholar] [CrossRef] [Green Version]
- Defrancesco, M.; Niederstätter, H.; Parson, W.; Oberacher, H.; Hinterhuber, H.; Canazei, M.; Bidner, J.; Deisenhammer, E.A.; Kemmler, G.; Weiss, E.M.; et al. Influence of Serotonin Transporter Genotype and Catechol-O-Methyltransferase Val158Met Polymorphism on Recognition of Emotional Faces. J. Int. Neuropsychol. Soc. JINS 2011, 17, 1014–1020. [Google Scholar] [CrossRef]
- Demaree, H.A.; Pu, J.; Jesberger, J.; Feeny, N.; Jeng, L.; Everhart, D.E.; Duerk, J.; Tkach, J. 5HTTLPR Predicts Left Fusiform Gyrus Activation to Positive Emotional Stimuli. Magn. Reson. Imaging 2009, 27, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Plieger, T.; Melchers, M.; Vetterlein, A.; Görtz, J.; Kuhn, S.; Ruppel, M.; Reuter, M. The Serotonin Transporter Polymorphism (5-HTTLPR) and Coping Strategies Influence Successful Emotion Regulation in an Acute Stress Situation: Physiological Evidence. Int. J. Psychophysiol. 2017, 114, 31–37. [Google Scholar] [CrossRef]
- Lang, P.J.; Bradley, M.M.; Cuthbert, B.N. International Affective Picture System (IAPS): Instruction Manual and Affective Ratings; Technical Report A-4; The Center for Research in Psychophysiology: Gainesville, FL, USA, 1999. [Google Scholar]
- Bradley, M.M.; Lang, P.J. Measuring Emotion: The Self-Assessment Manikin and the Semantic Differential—ScienceDirect. J. Behav. Ther. Exp. Psychiatry 1994, 25, 49–59. [Google Scholar] [CrossRef]
- Janke, W.; Erdmann, G.; Kallus, K.W. Der Streßverarbeitungsfragebogen (SVF 120). Kurzbeschreibung und Grundlegende Kennwerte; Hogrefe: Göttingen, Geramny, 1997. [Google Scholar]
- Marini, S.; Bagnoli, S.; Bessi, V.; Tedde, A.; Bracco, L.; Sorbi, S.; Nacmias, B. Implication of Serotonin-Transporter (5-HTT) Gene Polymorphism in Subjective Memory Complaints and Mild Cognitive Impairment (MCI). Arch. Gerontol. Geriatr. 2011, 52, e71–e74. [Google Scholar] [CrossRef]
- Pacheco, J.; Beevers, C.G.; McGeary, J.E.; Schnyer, D.M. Memory Monitoring Performance and PFC Activity Are Associated with 5-HTTLPR Genotype in Older Adults. Neuropsychologia 2012, 50, 2257–2270. [Google Scholar] [CrossRef] [Green Version]
- Grünblatt, E.; Löffler, C.; Zehetmayer, S.; Jungwirth, S.; Tragl, K.-H.; Riederer, P.; Fischer, P. Association Study of the 5-HTTLPR Polymorphism and Depression in 75-Year-Old Nondemented Subjects from the Vienna Transdanube Aging (VITA) Study. J. Clin. Psychiatry 2006, 67, 1373–1378. [Google Scholar] [CrossRef]
- O’Hara, R.; Schröder, C.M.; Mahadevan, R.; Schatzberg, A.F.; Lindley, S.; Fox, S.; Weiner, M.; Kraemer, H.C.; Noda, A.; Lin, X.; et al. Serotonin Transporter Polymorphism, Memory and Hippocampal Volume in the Elderly: Association and Interaction with Cortisol. Mol. Psychiatry 2007, 12, 544–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charles, S.T. Strength and Vulnerability Integration (SAVI): A Model of Emotional Well-Being Across Adulthood. Psychol. Bull. 2010, 136, 1068–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Hillard, C.J. Adaptations in Endocannabinoid Signaling in Response to Repeated Homotypic Stress: A Novel Mechanism for Stress Habituation. Eur. J. Neurosci. 2008, 27, 2821–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirz, L.; Reuter, M.; Felten, A.; Schwabe, L. An Endocannabinoid Receptor Polymorphism Modulates Affective Processing under Stress. Soc. Cogn. Affect. Neurosci. 2018, 13, 1177–1189. [Google Scholar] [CrossRef] [Green Version]
- Fairfield, B.; Mammarella, N.; Fontanella, L.; Sarra, A.; D’Aurora, M.; Stuppia, L.; Gatta, V. Aging and the Combined Effects of ADRA2B and CB1 Deletions on Affective Working Memory. Sci. Rep. 2019, 9, 4081. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.-J.; Zheng, D.; Li, K.-X.; Yang, J.-M.; Pan, H.-Q.; Yu, X.-D.; Fu, J.-Y.; Zhu, Y.; Sun, Q.-X.; Tang, M.-Y.; et al. Cannabinoid CB1 Receptors in the Amygdalar Cholecystokinin Glutamatergic Afferents to Nucleus Accumbens Modulate Depressive-like Behavior. Nat. Med. 2019, 25, 337–349. [Google Scholar] [CrossRef]
- Cheng, A.; Hou, Y.; Mattson, M.P. Mitochondria and Neuroplasticity. ASN Neuro 2010, 2, e00045. [Google Scholar] [CrossRef]
- Markham, A.; Bains, R.; Franklin, P.; Spedding, M. Changes in Mitochondrial Function Are Pivotal in Neurodegenerative and Psychiatric Disorders: How Important Is BDNF? Br. J. Pharmacol. 2014, 171, 2206–2229. [Google Scholar] [CrossRef] [Green Version]
- Roses, A.D.; Lutz, M.W.; Crenshaw, D.G.; Grossman, I.; Saunders, A.M.; Gottschalk, W.K. TOMM40 and APOE: Requirements for Replication Studies of Association with Age of Disease Onset and Enrichment of a Clinical Trial. Alzheimers Dement. J. Alzheimers Assoc. 2013, 9, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Lutz, M.W.; Crenshaw, D.G.; Saunders, A.M.; Roses, A.D. Genetic Variation at a Single Locus and Age of Onset for Alzheimer’s Disease. Alzheimers Dement. J. Alzheimers Assoc. 2010, 6, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Carrasquillo, M.M.; Morgan, K. Commentary on Functional Analysis of APOE Locus Genetic Variation Implicates Regional Enhancers in the Regulation of Both TOMM40 and APOE. J. Hum. Genet. 2012, 57, 3–4. [Google Scholar] [CrossRef] [Green Version]
- McFarquhar, M.; Elliott, R.; McKie, S.; Thomas, E.; Downey, D.; Mekli, K.; Toth, Z.G.; Anderson, I.M.; Deakin, J.F.W.; Juhasz, G. TOMM40 Rs2075650 May Represent a New Candidate Gene for Vulnerability to Major Depressive Disorder. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2014, 39, 1743–1753. [Google Scholar] [CrossRef]
- Gottschalk, W.K.; Lutz, M.W.; He, Y.T.; Saunders, A.M.; Burns, D.K.; Roses, A.D.; Chiba-Falek, O. The Broad Impact of TOM40 on Neurodegenerative Diseases in Aging. J. Park. Dis. Alzheimers Dis. 2014, 1, 12. [Google Scholar] [CrossRef]
- Spano, M.; Signorelli, M.; Vitaliani, R.; Aguglia, E.; Giometto, B. The Possible Involvement of Mitochondrial Dysfunctions in Lewy Body Dementia: A Systematic Review. Funct. Neurol. 2015, 30, 151–158. [Google Scholar] [CrossRef]
- Čulman, J.; Kvetňansky, T.; Serova, L.V.; Tigranjan, R.A.; Macho, L. Serotonin in Individual Hypothalamic Nuclei of Rats after Space Flight on Biosatellite Cosmos 1129—ScienceDirect. Acta Astronaut. 1985, 12, 373–376. [Google Scholar] [CrossRef]
- Hyde, T.M.; Wu, L.-C.; Krasnov, I.B.; Sigworth, S.K.; Daunton, N.G.; D’Amelio, F. Quantitative Autoradiographic Analysis of Muscarinic Cholinergic and GABAA (Benzodiazepine) Receptors in the Forebrain of Rats Flown on the Soviet Biosatellite COSMOS 2044. Brain Res. 1992, 593, 291–294. [Google Scholar] [CrossRef]
- Kvetǹanskỳ, R.; Čulman, J.; Serova, L.V.; Tigranjan, R.A.; Torda, T.; Macho, L. Catecholamines and Their Enzymes in Discrete Brain Areas of Rats after Space Flight on Biosatellites Cosmos. Acta Astronaut. 1983, 10, 295–300. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Kulikov, A.V.; Kondaurova, E.M.; Tsybko, A.S.; Kulikova, E.A.; Krasnov, I.B.; Shenkman, B.S.; Sychev, V.N.; Bazhenova, E.Y.; Sinyakova, N.A.; et al. Effect of Actual Long-Term Spaceflight on BDNF, TrkB, P75, BAX and BCL-XL Genes Expression in Mouse Brain Regions. Neuroscience 2015, 284, 730–736. [Google Scholar] [CrossRef]
- Pompeiano, O.; d’Ascanio, P.; Balaban, E.; Centini, C.; Pompeiano, M. Gene Expression in Autonomic Areas of the Medulla and the Central Nucleus of the Amygdala in Rats during and after Space Flight. Neuroscience 2004, 124, 53–69. [Google Scholar] [CrossRef]
- Popova, N.K.; Kulikov, A.V.; Kondaurova, E.M.; Tsybko, A.S.; Kulikova, E.A.; Krasnov, I.B.; Shenkman, B.S.; Bazhenova, E.Y.; Sinyakova, N.A.; Naumenko, V.S. Risk Neurogenes for Long-Term Spaceflight: Dopamine and Serotonin Brain System. Mol. Neurobiol. 2015, 51, 1443–1451. [Google Scholar] [CrossRef]
- Santucci, D.; Kawano, F.; Ohira, T.; Terada, M.; Nakai, N.; Francia, N.; Alleva, E.; Aloe, L.; Ochiai, T.; Cancedda, R.; et al. Evaluation of Gene, Protein and Neurotrophin Expression in the Brain of Mice Exposed to Space Environment for 91 Days. PLoS ONE 2012, 7, e40112. [Google Scholar] [CrossRef] [Green Version]
- Tsybko, A.; Ilchibaeva, T.; Kulikov, A.; Kulikova, E.; Krasnov, I.B.; Sychev, V.N.; Shenkman, B.S.; Popova, N.; Naumenko, V. Effect of Microgravity on Glial Cell Line-Derived Neurotrophic Factor and Cerebral Dopamine Neurotrophic Factor Gene Expression in the Mouse Brain: Effect of Microgravity on GDNF and CDNF Genes. J. Neurosci. Res. 2015, 93, 1399–1404. [Google Scholar] [CrossRef]
- Popova, N.K.; Kulikov, A.V.; Naumenko, V.S. Spaceflight and Brain Plasticity: Spaceflight Effects on Regional Expression of Neurotransmitter Systems and Neurotrophic Factors Encoding Genes. Neurosci. Biobehav. Rev. 2020, 119, 396–405. [Google Scholar] [CrossRef]
- Afshinnekoo, E.; Scott, R.T.; MacKay, M.J.; Pariset, E.; Cekanaviciute, E.; Barker, R.; Gilroy, S.; Hassane, D.; Smith, S.M.; Zwart, S.R.; et al. Fundamental Biological Features of Spaceflight: Advancing the Field to Enable Deep-Space Exploration. Cell 2020, 183, 1162–1184. [Google Scholar] [CrossRef] [PubMed]
- Pariset, E.; Bertucci, A.; Petay, M.; Malkani, S.; Macha, A.L.; Lima, I.G.P.; Gonzalez, V.G.; Tin, A.S.; Tang, J.; Plante, I.; et al. DNA Damage Baseline Predicts Space Radiation and Radio-Therapeutic Resilience. Cell Rep. 2020, 33, 108434. [Google Scholar] [CrossRef] [PubMed]
- Luxton, J.J.; McKenna, M.J.; Lewis, A.; Taylor, L.E.; George, K.A.; Dixit, S.M.; Moniz, M.; Benegas, W.; Mackay, M.J.; Mozsary, C.; et al. Telomere Length Dynamics and DNA Damage Responses Associated with Long-Duration Spaceflight. Cell Rep. 2020, 33, 108457. [Google Scholar] [CrossRef] [PubMed]
- Luxton, J.J.; McKenna, M.J.; Taylor, L.E.; George, K.A.; Zwart, S.R.; Crucian, B.E.; Drel, V.R.; Garrett-Bakelman, F.E.; Mackay, M.J.; Butler, D.; et al. Temporal Telomere and DNA Damage Responses in the Space Radiation Environment. Cell Rep. 2020, 33, 108435. [Google Scholar] [CrossRef] [PubMed]
- Kam, W.W.-Y.; Banati, R.B. Effects of Ionizing Radiation on Mitochondria. Free Radic. Biol. Med. 2013, 65, 607–619. [Google Scholar] [CrossRef]
- Gertz, M.L.; Chin, C.R.; Tomoiaga, D.; MacKay, M.; Chang, C.; Butler, D.; Afshinnekoo, E.; Bezdan, D.; Schmidt, M.A.; Mozsary, C.; et al. Multi-Omic, Single-Cell, and Biochemical Profiles of Astronauts Guide Pharmacological Strategies for Returning to Gravity. Cell Rep. 2020, 33, 108429. [Google Scholar] [CrossRef]
- Jiang, P.; Green, S.J.; Chlipala, G.E.; Turek, F.W.; Vitaterna, M.H. Reproducible Changes in the Gut Microbiome Suggest a Shift in Microbial and Host Metabolism during Spaceflight. Microbiome 2019, 7, 113. [Google Scholar] [CrossRef]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A Multidimensional Analysis of a Year-Long Human Spaceflight. Science 2019, 364, eaau8650. [Google Scholar] [CrossRef]
- Fani, L.; Hilal, S.; Sedaghat, S.; Broer, L.; Licher, S.; Arp, P.P.; van Meurs, J.B.J.; Ikram, M.K.; Ikram, M.A. Telomere Length and the Risk of Alzheimer’s Disease: The Rotterdam Study. J. Alzheimers Dis. 2020, 73, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Kanas, N. Humans in Space; Springer International Publishing: Cham, Switzerland, 2015; ISBN 978-3-319-18868-3.
- Cucinotta, F.A.; Durante, M. Cancer Risk from Exposure to Galactic Cosmic Rays: Implications for Space Exploration by Human Beings. Lancet Oncol. 2006, 7, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Cucinotta, F.A.; Hu, S.; Schwadron, N.A.; Kozarev, K.; Townsend, L.W.; Kim, M.-H.Y. Space Radiation Risk Limits and Earth-Moon-Mars Environmental Models: SPACE RADIATION RISK LIMITS. Space Weather 2010, 8, S00E09. [Google Scholar] [CrossRef]
- Zeitlin, C.; Hassler, D.M.; Cucinotta, F.A.; Ehresmann, B.; Wimmer-Schweingruber, R.F.; Brinza, D.E.; Kang, S.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Measurements of Energetic Particle Radiation in Transit to Mars on the Mars Science Laboratory. Science 2013, 340, 1080–1084. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Pei, W.; Hu, W. A Current Overview of the Biological Effects of Combined Space Environmental Factors in Mammals. Front. Cell Dev. Biol. 2022, 10, 861006. [Google Scholar] [CrossRef]
- Hamilton, S.A.; Pecaut, M.J.; Gridley, D.S.; Travis, N.D.; Bandstra, E.R.; Willey, J.S.; Nelson, G.A.; Bateman, T.A. A Murine Model for Bone Loss from Therapeutic and Space-Relevant Sources of Radiation. J. Appl. Physiol. 2006, 101, 789–793. [Google Scholar] [CrossRef]
- Kondo, H.; Searby, N.D.; Mojarrab, R.; Phillips, J.; Alwood, J.; Yumoto, K.; Almeida, E.A.C.; Limoli, C.L.; Globus, R.K. Total-Body Irradiation of Postpubertal Mice with (137)Cs Acutely Compromises the Microarchitecture of Cancellous Bone and Increases Osteoclasts. Radiat. Res. 2009, 171, 283–289. [Google Scholar] [CrossRef]
- Bokhari, R.S.; Metzger, C.E.; Black, J.M.; Franklin, K.A.; Boudreaux, R.D.; Allen, M.R.; Macias, B.R.; Hogan, H.A.; Braby, L.A.; Bloomfield, S.A. Positive Impact of Low-Dose, High-Energy Radiation on Bone in Partial-and/or Full-Weightbearing Mice. NPJ Microgravity 2019, 5, 13. [Google Scholar] [CrossRef]
- Juhl, O.J.; Buettmann, E.G.; Friedman, M.A.; DeNapoli, R.C.; Hoppock, G.A.; Donahue, H.J. Update on the Effects of Microgravity on the Musculoskeletal System. Npj Microgravity 2021, 7, 1–15. [Google Scholar] [CrossRef]
- Willey, J.S.; Britten, R.A.; Blaber, E.; Tahimic, C.G.T.; Chancellor, J.; Mortreux, M.; Sanford, L.D.; Kubik, A.J.; Delp, M.D.; Mao, X.W. The Individual and Combined Effects of Spaceflight Radiation and Microgravity on Biologic Systems and Functional Outcomes. J. Environ. Sci. Health Part C Toxicol. Carcinog. 2021, 39, 129–179. [Google Scholar] [CrossRef]
- Zhang, L.-F.; Hargens, A.R. Spaceflight-Induced Intracranial Hypertension and Visual Impairment: Pathophysiology and Countermeasures. Physiol. Rev. 2018, 98, 59–87. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.W.; Archambeau, J.O.; Kubínová, L.; Boyle, S.; Petersen, G.; Grove, R. Quantification of Rat Retinal Growth and Vascular Population Changes after Single and Split Doses of Proton Irradiation: Translational Study Using Stereology Methods. Radiat. Res. 2003, 160, 5–13. [Google Scholar] [CrossRef]
- Taylor, C.R.; Hanna, M.; Behnke, B.J.; Stabley, J.N.; McCullough, D.J.; Davis, R.T.; Ghosh, P.; Papadopoulos, A.; Muller-Delp, J.M.; Delp, M.D. Spaceflight-Induced Alterations in Cerebral Artery Vasoconstrictor, Mechanical, and Structural Properties: Implications for Elevated Cerebral Perfusion and Intracranial Pressure. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 2282–2292. [Google Scholar] [CrossRef] [Green Version]
- Morgan, W.H.; Balaratnasingam, C.; Lind, C.R.P.; Colley, S.; Kang, M.H.; House, P.H.; Yu, D.-Y. Cerebrospinal Fluid Pressure and the Eye. Br. J. Ophthalmol. 2016, 100, 71–77. [Google Scholar] [CrossRef]
- Mao, X.W.; Boerma, M.; Rodriguez, D.; Campbell-Beachler, M.; Jones, T.; Stanbouly, S.; Sridharan, V.; Nishiyama, N.C.; Wroe, A.; Nelson, G.A. Combined Effects of Low-Dose Proton Radiation and Simulated Microgravity on the Mouse Retina and the Hematopoietic System. Radiat. Res. 2019, 192, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.R.; Albrecht, M.H.; Collins, H.R.; Asemani, D.; Chatterjee, A.R.; Spampinato, M.V.; Zhu, X.; Chimowitz, M.I.; Antonucci, M.U. Effects of Spaceflight on Astronaut Brain Structure as Indicated on MRI. N. Engl. J. Med. 2017, 377, 1746–1753. [Google Scholar] [CrossRef] [PubMed]
- Carriot, J.; Mackrous, I.; Cullen, K.E. Challenges to the Vestibular System in Space: How the Brain Responds and Adapts to Microgravity. Front. Neural Circuits 2021, 15, 760313. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Jeffrey, A.; Nguyen, K.; Kumar, S.; Toshimasa, O.; Hirose, R.; Reue, K.; Vergnes, L.; Kinchen, J.; de Vellis, J. Simulated Microgravity Enhances Oligodendrocyte Mitochondrial Function and Lipid Metabolism. J. Neurosci. Res. 2016, 94, 1434–1450. [Google Scholar] [CrossRef] [PubMed]
- Bellone, J.A.; Gifford, P.S.; Nishiyama, N.C.; Hartman, R.E.; Mao, X.W. Long-Term Effects of Simulated Microgravity and/or Chronic Exposure to Low-Dose Gamma Radiation on Behavior and Blood-Brain Barrier Integrity. NPJ Microgravity 2016, 2, 16019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashcheev, V.V.; Chekin, S.Y.; Karpenko, S.V.; Maksioutov, M.A.; Menyaylo, A.N.; Tumanov, K.A.; Kochergina, E.V.; Kashcheeva, P.V.; Gorsky, A.I.; Shchukina, N.V.; et al. Radiation Risk of Cardiovascular Diseases in the Cohort of Russian Emergency Workers of the Chernobyl Accident. Health Phys. 2017, 113, 23–29. [Google Scholar] [CrossRef]
- Little, M.P.; Azizova, T.V.; Bazyka, D.; Bouffler, S.D.; Cardis, E.; Chekin, S.; Chumak, V.V.; Cucinotta, F.A.; de Vathaire, F.; Hall, P.; et al. Systematic Review and Meta-Analysis of Circulatory Disease from Exposure to Low-Level Ionizing Radiation and Estimates of Potential Population Mortality Risks. Environ. Health Perspect. 2012, 120, 1503–1511. [Google Scholar] [CrossRef] [Green Version]
- Soucy, K.G.; Lim, H.K.; Kim, J.H.; Oh, Y.; Attarzadeh, D.O.; Sevinc, B.; Kuo, M.M.; Shoukas, A.A.; Vazquez, M.E.; Berkowitz, D.E. HZE 56Fe-Ion Irradiation Induces Endothelial Dysfunction in Rat Aorta: Role of Xanthine Oxidase. Radiat. Res. 2011, 176, 474–485. [Google Scholar] [CrossRef]
- Ade, C.J.; Broxterman, R.M.; Charvat, J.M.; Barstow, T.J. Incidence Rate of Cardiovascular Disease End Points in the National Aeronautics and Space Administration Astronaut Corps. J. Am. Heart Assoc. 2017, 6, e005564. [Google Scholar] [CrossRef]
- Hughson, R.L.; Helm, A.; Durante, M. Heart in Space: Effect of the Extraterrestrial Environment on the Cardiovascular System. Nat. Rev. Cardiol. 2018, 15, 167–180. [Google Scholar] [CrossRef]
- Ghosh, P.; Behnke, B.J.; Stabley, J.N.; Kilar, C.R.; Park, Y.; Narayanan, A.; Alwood, J.S.; Shirazi-Fard, Y.; Schreurs, A.-S.; Globus, R.K.; et al. Effects of High-LET Radiation Exposure and Hindlimb Unloading on Skeletal Muscle Resistance Artery Vasomotor Properties and Cancellous Bone Microarchitecture in Mice. Radiat. Res. 2016, 185, 257–266. [Google Scholar] [CrossRef]
- Delp, M.D.; Charvat, J.M.; Limoli, C.L.; Globus, R.K.; Ghosh, P. Apollo Lunar Astronauts Show Higher Cardiovascular Disease Mortality: Possible Deep Space Radiation Effects on the Vascular Endothelium. Sci. Rep. 2016, 6, 29901. [Google Scholar] [CrossRef] [Green Version]
- Rodman, C.; Almeida-Porada, G.; George, S.K.; Moon, J.; Soker, S.; Pardee, T.; Beaty, M.; Guida, P.; Sajuthi, S.P.; Langefeld, C.D.; et al. In Vitro and in Vivo Assessment of Direct Effects of Simulated Solar and Galactic Cosmic Radiation on Human Hematopoietic Stem/Progenitor Cells. Leukemia 2017, 31, 1398–1407. [Google Scholar] [CrossRef]
- Blaber, E.; Sato, K.; Almeida, E.A.C. Stem Cell Health and Tissue Regeneration in Microgravity. Stem Cells Dev. 2014, 23, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, N.; Khandani, A.; Camirand, A.; Harrison, R.E. Effects of Microgravity on Osteoclast Bone Resorption and Osteoblast Cytoskeletal Organization and Adhesion. Bone 2011, 49, 965–974. [Google Scholar] [CrossRef]
- Hughes-Fulford, M.; Lewis, M.L. Effects of Microgravity on Osteoblast Growth Activation. Exp. Cell Res. 1996, 224, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Fitts, R.H.; Trappe, S.W.; Costill, D.L.; Gallagher, P.M.; Creer, A.C.; Colloton, P.A.; Peters, J.R.; Romatowski, J.G.; Bain, J.L.; Riley, D.A. Prolonged Space Flight-Induced Alterations in the Structure and Function of Human Skeletal Muscle Fibres. J. Physiol. 2010, 588, 3567–3592. [Google Scholar] [CrossRef]
- Buravkova, L.B.; Grigor’eva, O.V.; Rykova, M.P.; Grigor’ev, A.I. Cytotoxic Activity of Natural Killer Cells in Vitro under Microgravity. Dokl. Biol. Sci. Proc. Acad. Sci. USSR Biol. Sci. Sect. 2008, 421, 275–277. [Google Scholar] [CrossRef]
- Freed, L.E.; Langer, R.; Martin, I.; Pellis, N.R.; Vunjak-Novakovic, G. Tissue Engineering of Cartilage in Space. Proc. Natl. Acad. Sci. USA 1997, 94, 13885–13890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, M.L.; Reynolds, J.L.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight Alters Microtubules and Increases Apoptosis in Human Lymphocytes (Jurkat). FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1998, 12, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Algeda, F.R.; Eltahawy, N.A.; Shedid, S.M.; Saada, H.N. The Impact of Gamma-Radiation on the Cerebral- and Cerebellar- Cortex of Male Rats’ Brain. Brain Res. Bull. 2022, 186, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Sudevan, S.; Muto, K.; Higashitani, N.; Hashizume, T.; Higashibata, A.; Ellwood, R.A.; Deane, C.S.; Rahman, M.; Vanapalli, S.A.; Etheridge, T.; et al. Loss of Physical Contact in Space Alters the Dopamine System in C. elegans. iScience 2022, 25, 103762. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Kudrin, V.S.; Shtemberg, A.S. Serotonin and Noradrenaline Metabolism in the Brain of Rats under the Combined Action of Radiation and Hypogravity in a Ground-Based Experiment. Neurochem. J. 2019, 13, 57–61. [Google Scholar] [CrossRef]
- Strewe, C.; Feuerecker, M.; Nichiporuk, I.; Kaufmann, I.; Hauer, D.; Morukov, B.; Schelling, G.; Choukèr, A. Effects of Parabolic Flight and Spaceflight on the Endocannabinoid System in Humans. Rev. Neurosci. 2012, 23, 673–680. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Candidate Genes | Main Role | Relationships with Positive Memory |
|---|---|---|
| ADRA2B (209 bp/200 bp) | Regulation of neurotransmitter release from sympathetic nerves and adrenergic neurons in the CNS | Deletion (200 bp) carriers show enhanced emotional stimuli processing and memory |
| Deletion carriers have higher cognitive–affective flexibility compared with non-carriers | ||
| Deletion carriers preserve emotional faces’ recognition abilities under acute stress conditions | ||
| COMT (Val158/Met158) | Metabolism of catecholamines and l-dopa | Met158 polymorphism is linked to increased neural activation for negative stimuli Met158 allele carriers have reduced performance in emotion-related tasks |
| Met carriers have better WM performance, while Val carriers are better in processing negative stimuli | ||
| 5HTTLPR (S/S; L/L; S/L genotypes) | The serotonin transporter (5-HTT) gene mediates the serotonin reuptake from the intersynaptic space | S-allele carriers show decreased left fusiform gyrus activation to positive stimuli |
| S-allele carriers experience greater emotional responses to negative stimuli than l-carriers | ||
| L-allele carriers are more efficient in emotion regulation than s-carriers | ||
| CB1 (AA/AG/GG genotypes) | Modulate neurotransmission by inhibiting presynaptic Ca | Carriers of at least one copy of the minor A allele show improved consolidation for emotional events into autobiographical memory than GG genotype’s subjects |
| Carriers of the combination of CB1 and ADRA2B polymorphisms show enhanced memory and a positive effect bias | ||
| One copy of the minor A allele could be a protective factor against depression after stressful events | ||
| TOMM40 (intron 6: S/L/VL; intron 2: AA/GA/GG) | Allows protein import from the cytoplasm into mitochondria | S variant of intron6 is associated with enhanced memory performance and executive functions |
| AA genotype shows a positive memory bias compared with G-allele carriers |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mammarella, N.; Gatti, M.; Ceccato, I.; Di Crosta, A.; Di Domenico, A.; Palumbo, R. The Protective Role of Neurogenetic Components in Reducing Stress-Related Effects during Spaceflights: Evidence from the Age-Related Positive Memory Approach. Life 2022, 12, 1176. https://doi.org/10.3390/life12081176
Mammarella N, Gatti M, Ceccato I, Di Crosta A, Di Domenico A, Palumbo R. The Protective Role of Neurogenetic Components in Reducing Stress-Related Effects during Spaceflights: Evidence from the Age-Related Positive Memory Approach. Life. 2022; 12(8):1176. https://doi.org/10.3390/life12081176
Chicago/Turabian StyleMammarella, Nicola, Matteo Gatti, Irene Ceccato, Adolfo Di Crosta, Alberto Di Domenico, and Rocco Palumbo. 2022. "The Protective Role of Neurogenetic Components in Reducing Stress-Related Effects during Spaceflights: Evidence from the Age-Related Positive Memory Approach" Life 12, no. 8: 1176. https://doi.org/10.3390/life12081176
APA StyleMammarella, N., Gatti, M., Ceccato, I., Di Crosta, A., Di Domenico, A., & Palumbo, R. (2022). The Protective Role of Neurogenetic Components in Reducing Stress-Related Effects during Spaceflights: Evidence from the Age-Related Positive Memory Approach. Life, 12(8), 1176. https://doi.org/10.3390/life12081176