
Brassica oleracea L. var. botrytis Leaf Extract Alleviates Gentamicin-Induced Hepatorenal Injury in Rats—Possible Modulation of IL-1β and NF-κB Activity Assisted with Computational Approach

 , , , , , , , , , and
, , , , , , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Procedures and Chemicals
2.2. Plant Material
2.3. Extraction and Isolation of Plant Constituents
2.4. Calculation of Total Phenolic Content (TPC)
2.5. Animals
2.6. Animal Experiment
2.7. Biochemical Analysis of Antioxidant Markers
2.8. Assessment of Kidney and Liver Functions
2.9. Kidney and Liver Histopathological Examinations
2.10. Morphometric Analysis of Renal and Hepatic Tissues
2.11. Methods Used to Extract and Quantify mRNA
2.12. Western Blot Analysis
2.13. Computational Analysis
2.14. Statistical Analysis
3. Results
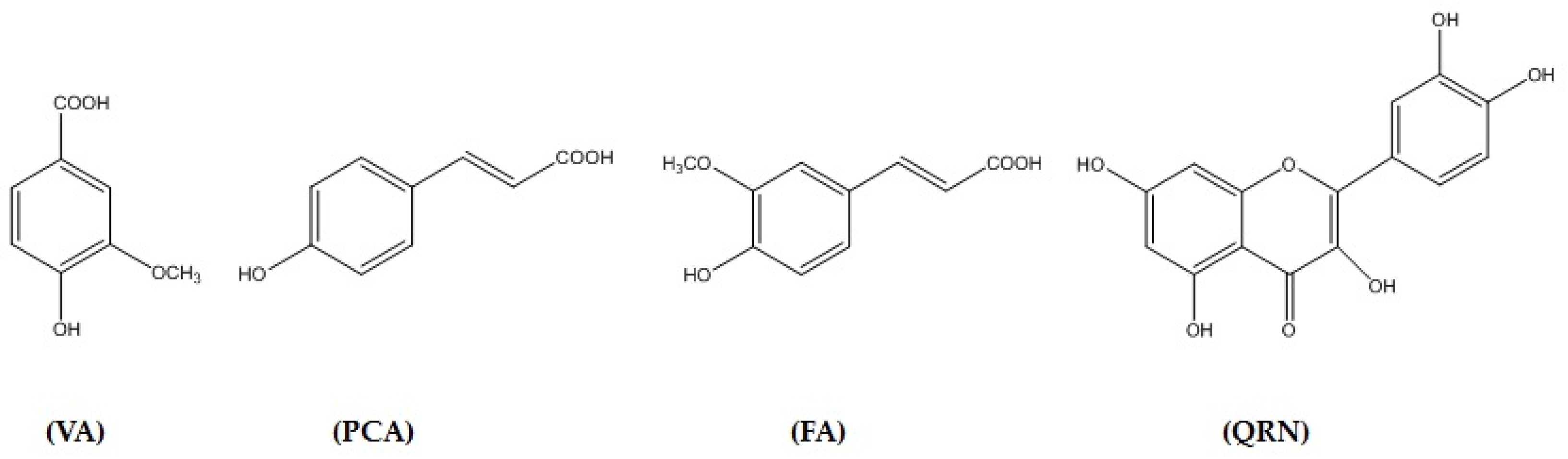
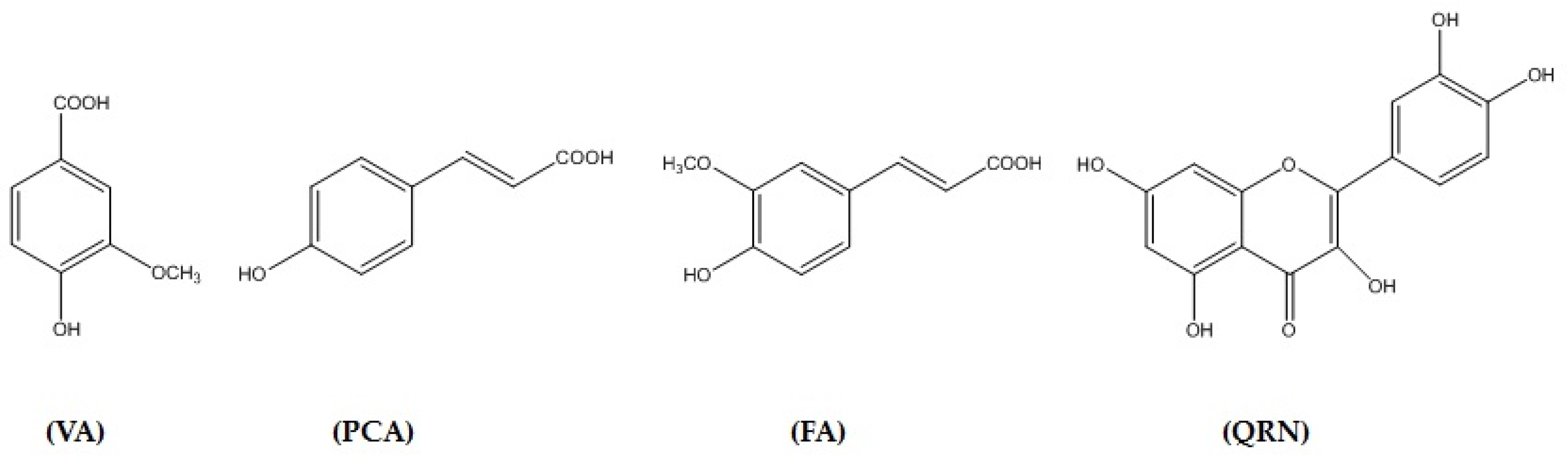
3.1. Isolation and Identification of Secondary Metabolites
3.2. TPC
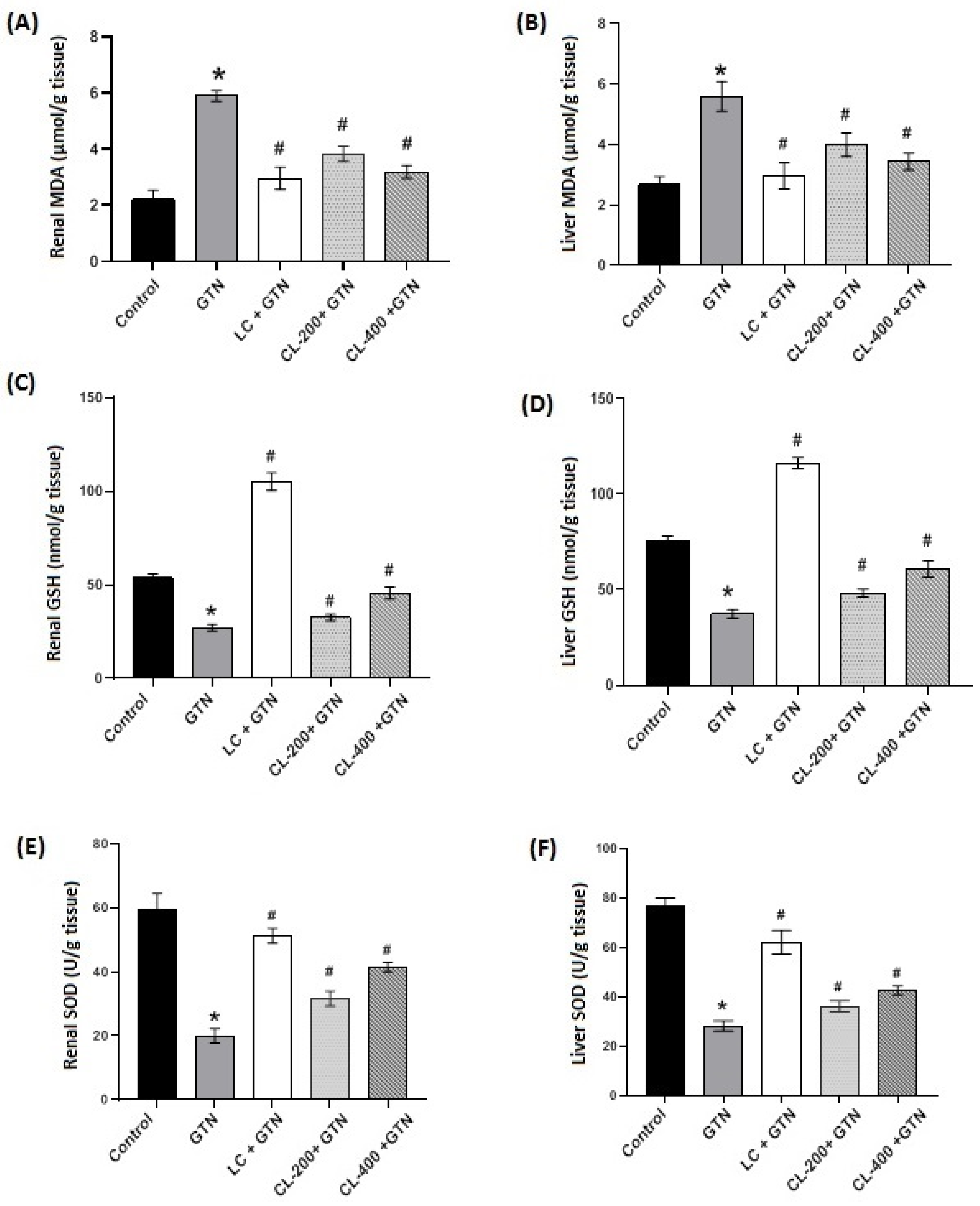
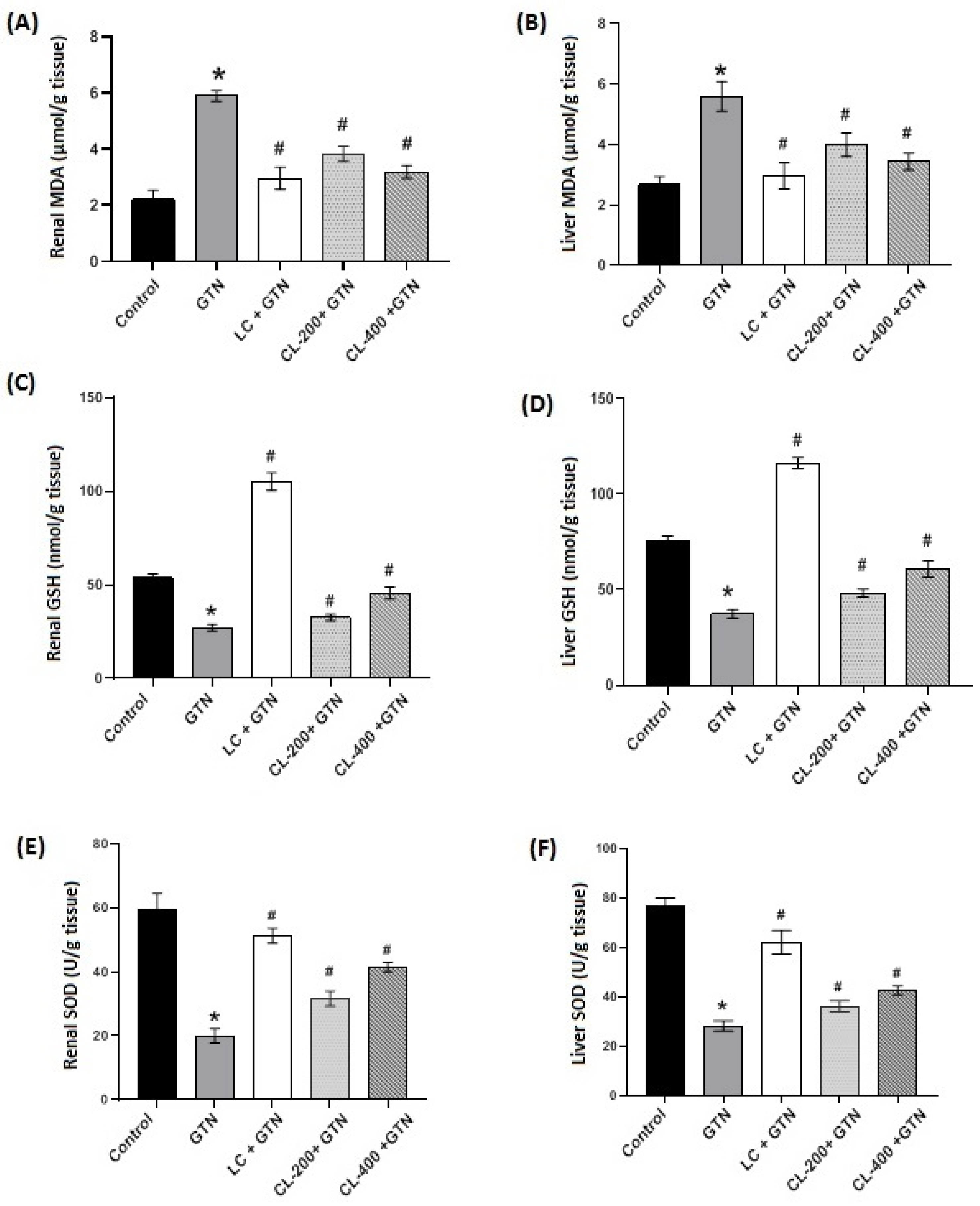
3.3. Effects of CL on the Levels/Contents of MDA, GSH and SOD
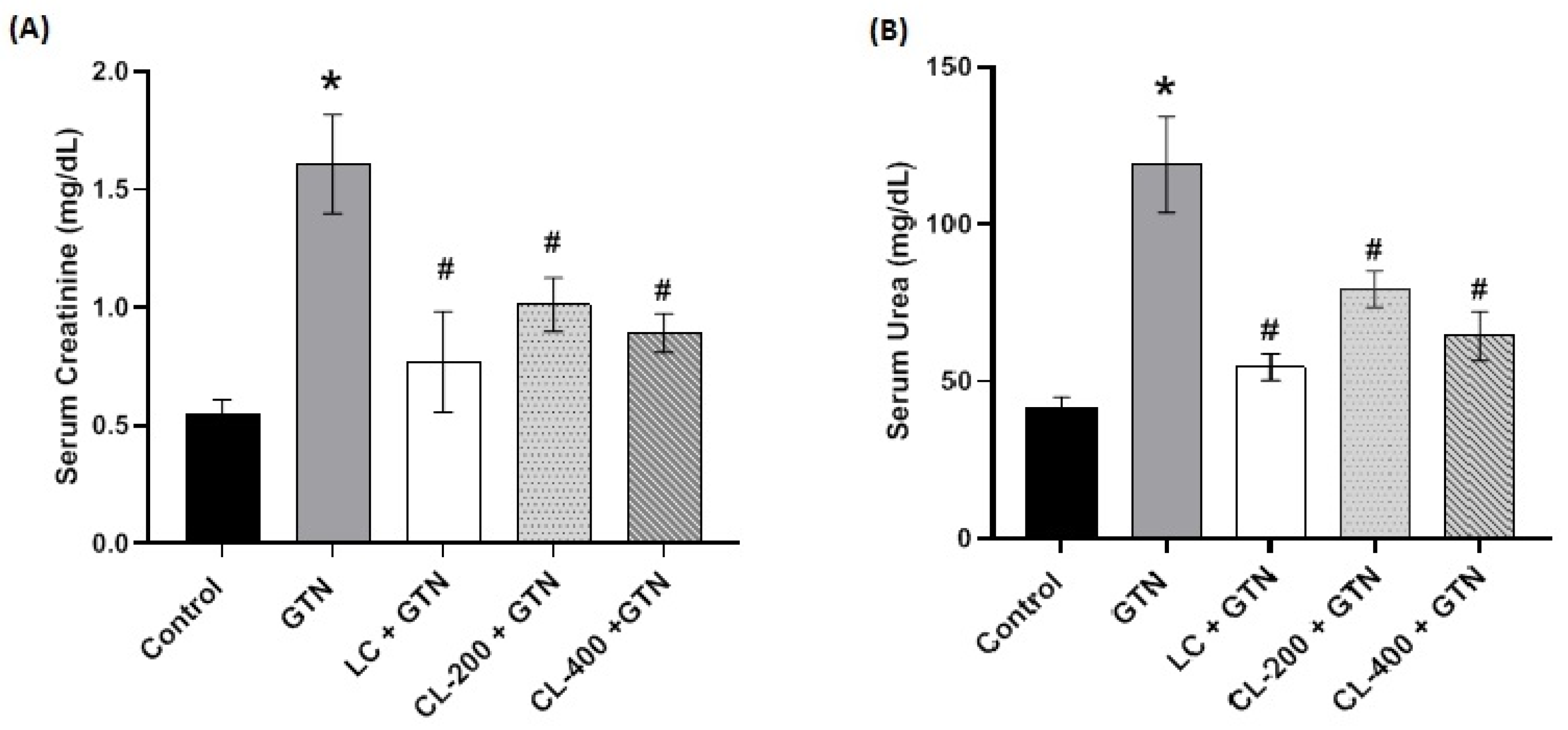
3.4. Effect of Caulifower Leaf Extract on Kidney Function Parameters
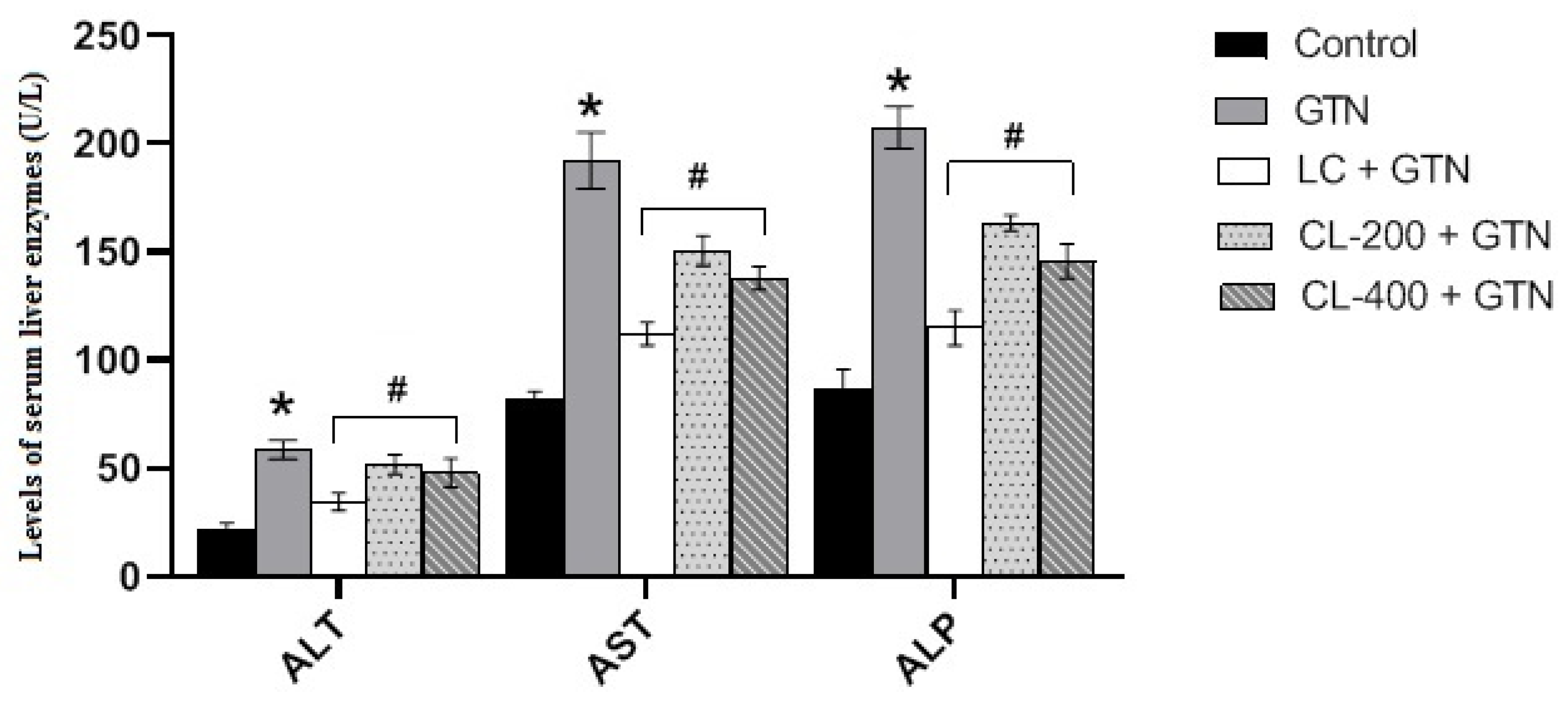
3.5. Effect of Caulifower Leaf Extract on Liver Function
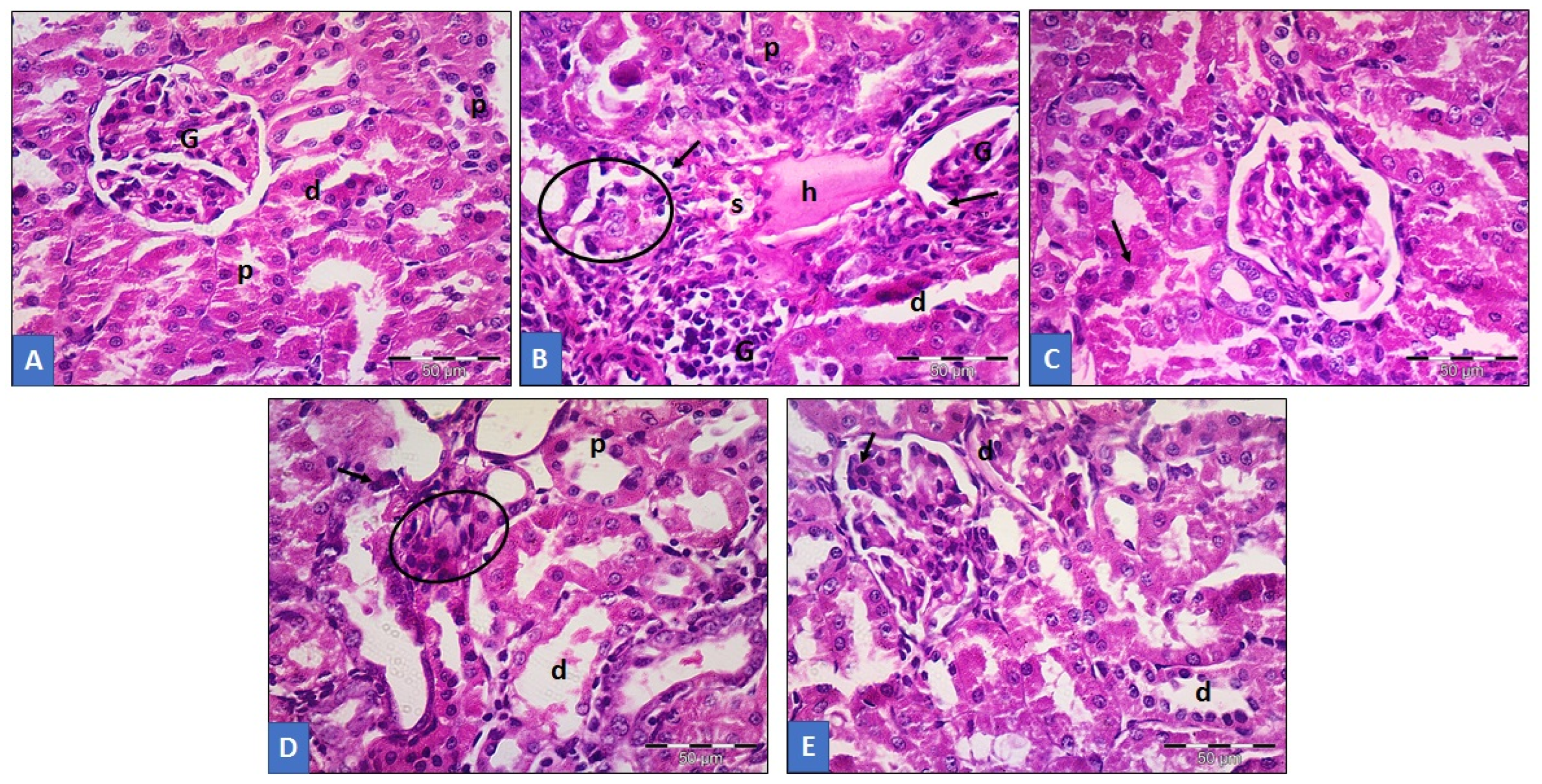
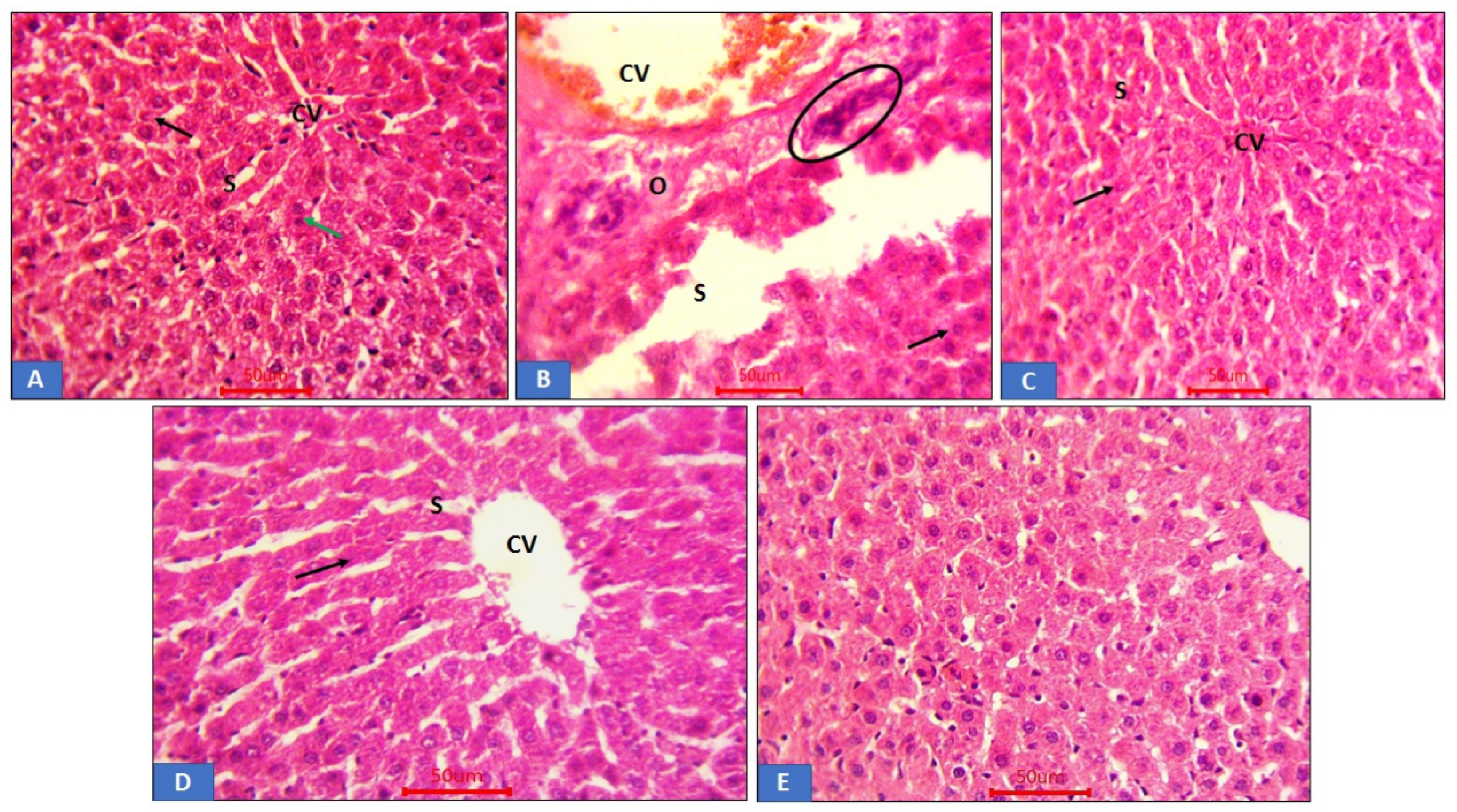
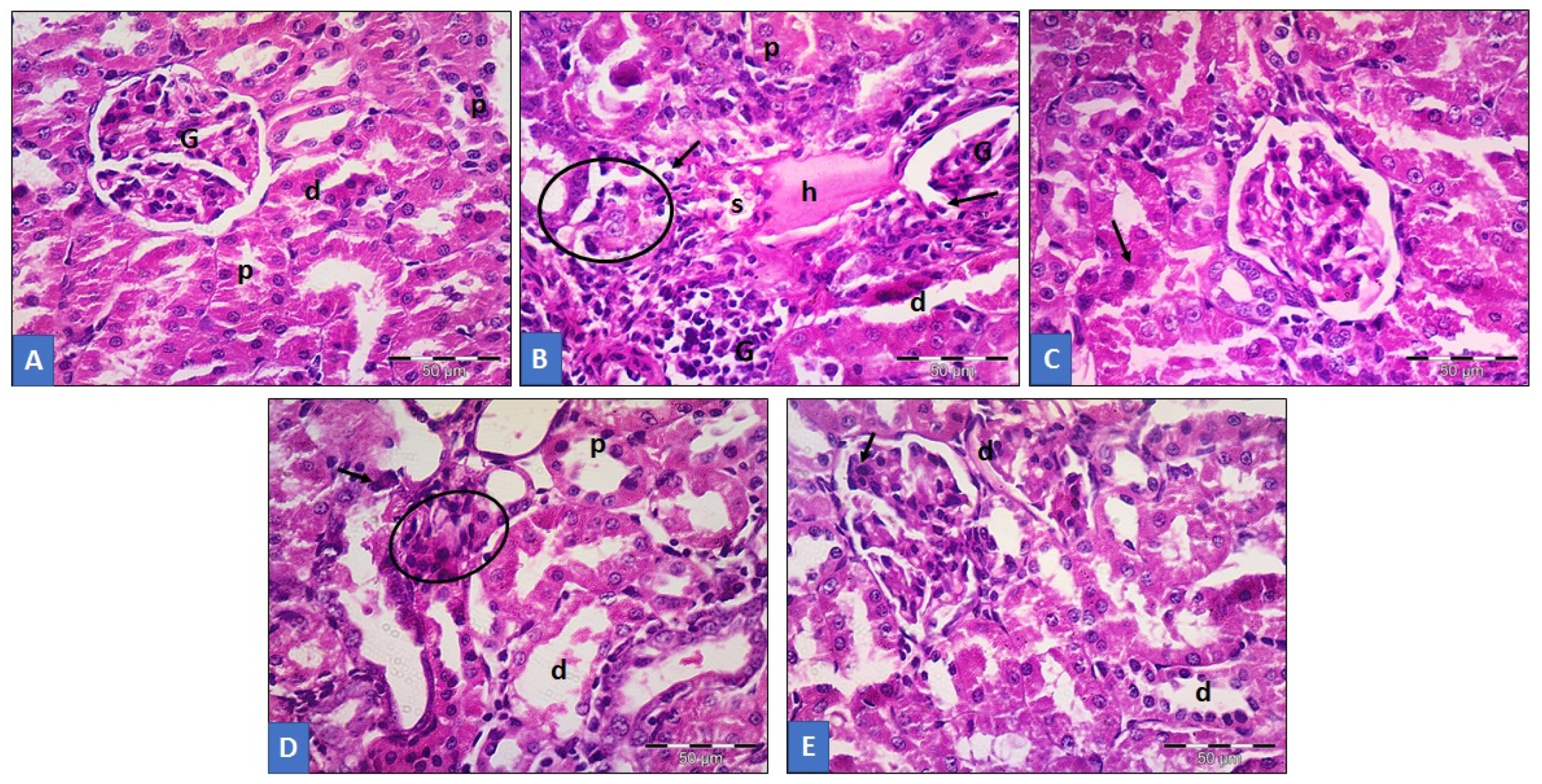
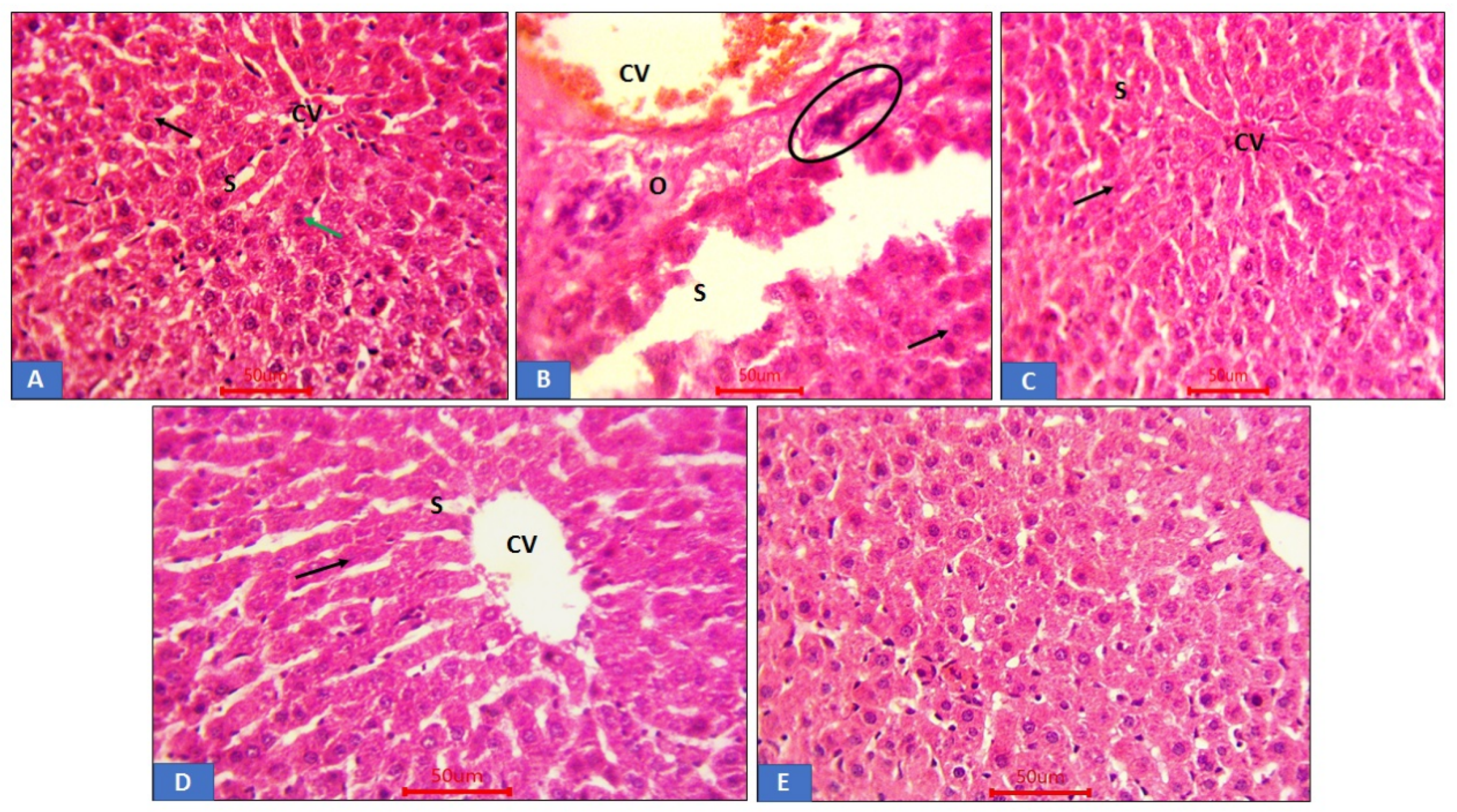
3.6. Histopathological Analysis of Renal Tissue
3.7. Histopathological Analysis of Liver Tissue
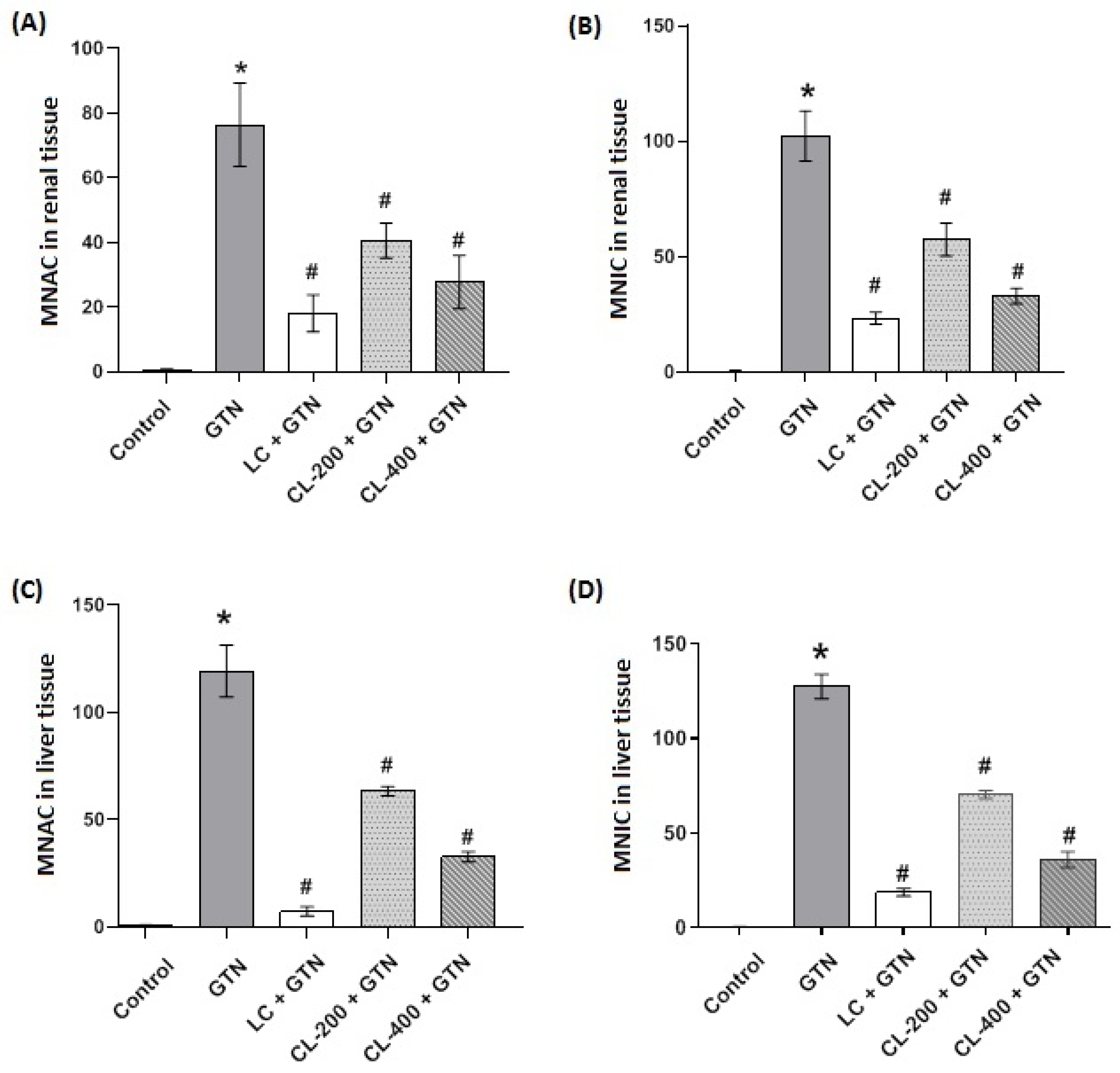
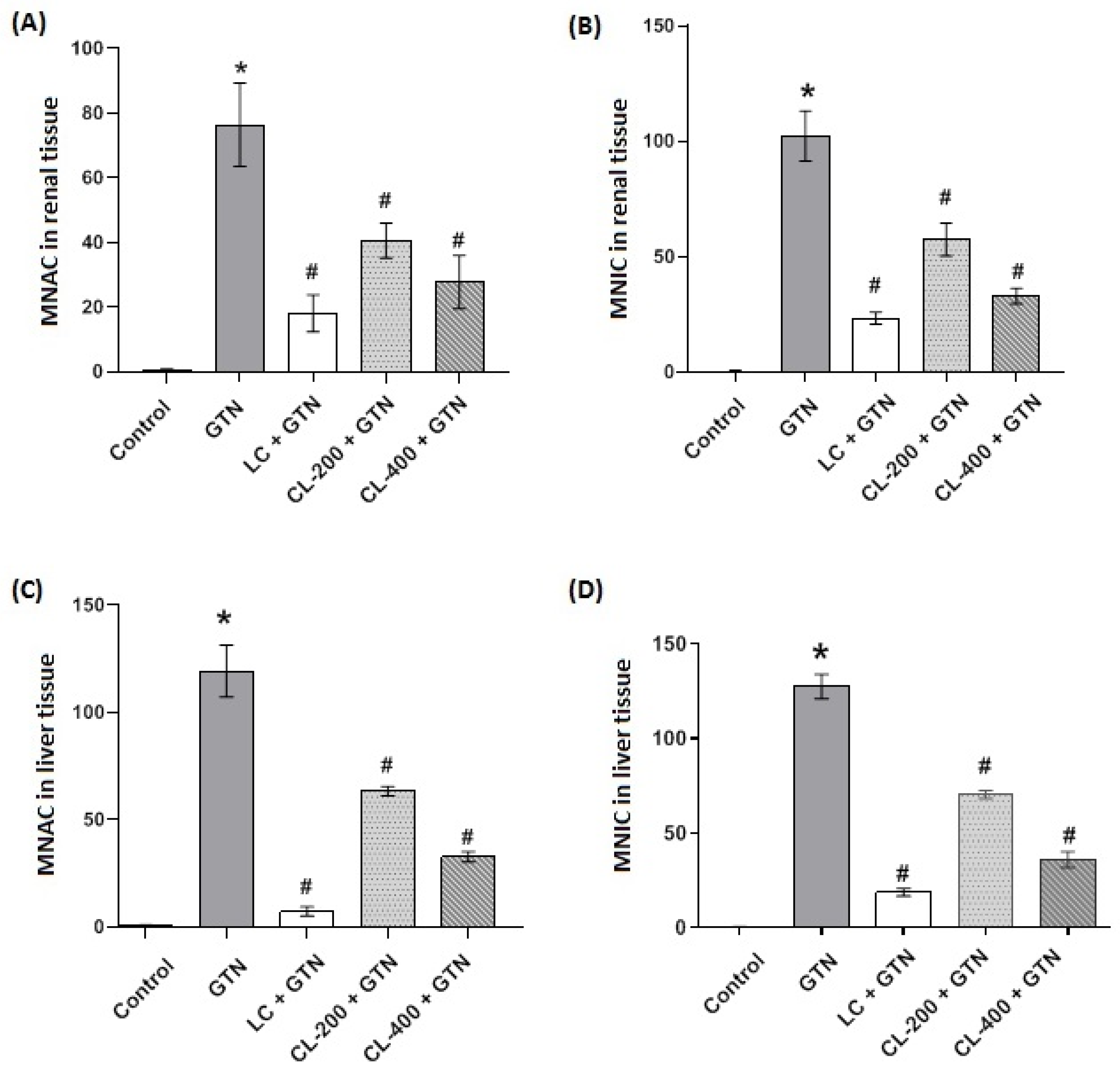
3.8. Morphometric Analysis of Renal and Liver Tissues
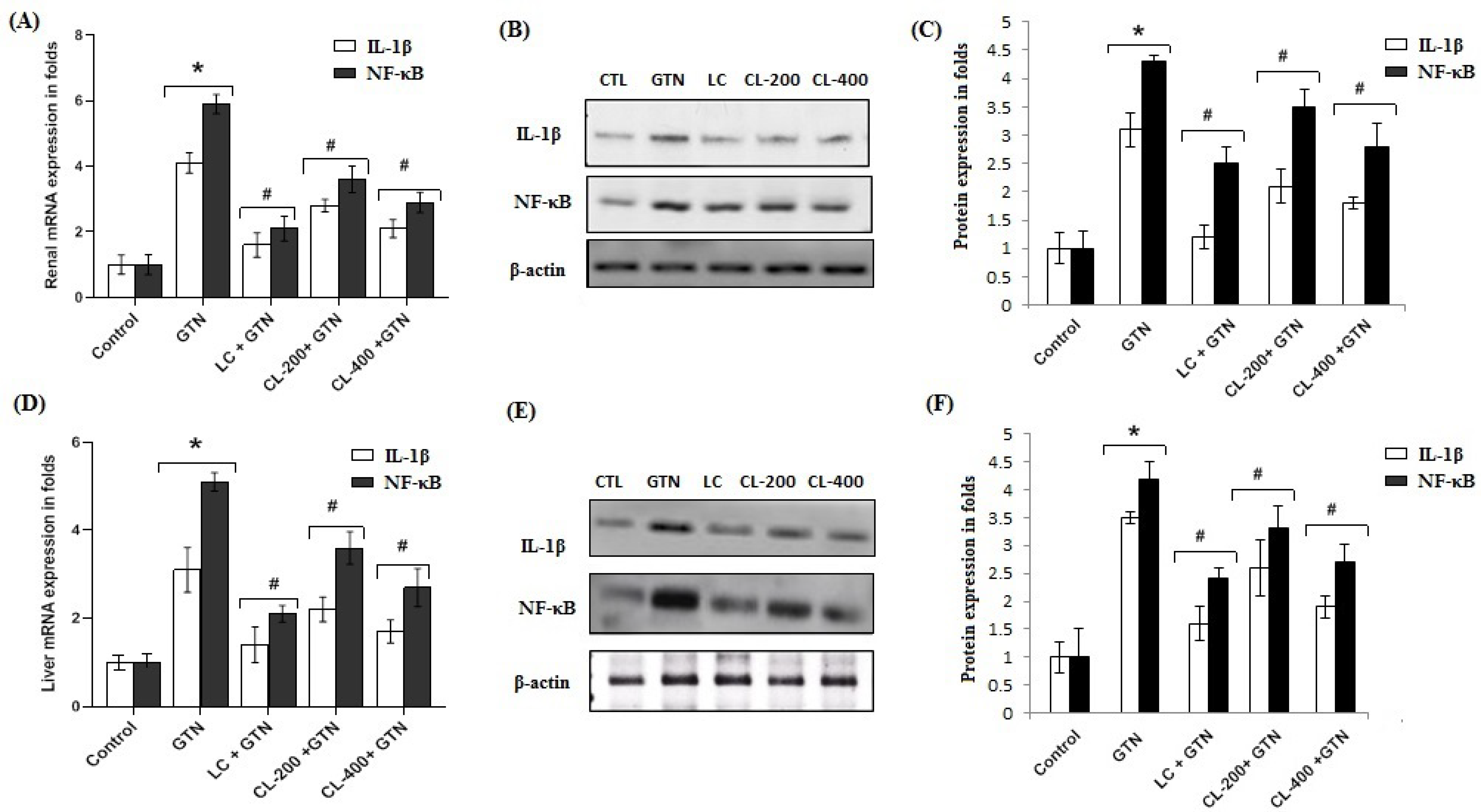
3.9. Effect of Caulifower Leaf Extract on mRNA and Protein Expressions of IL-1β and NF-κB Markers in Renal and Liver Tissues
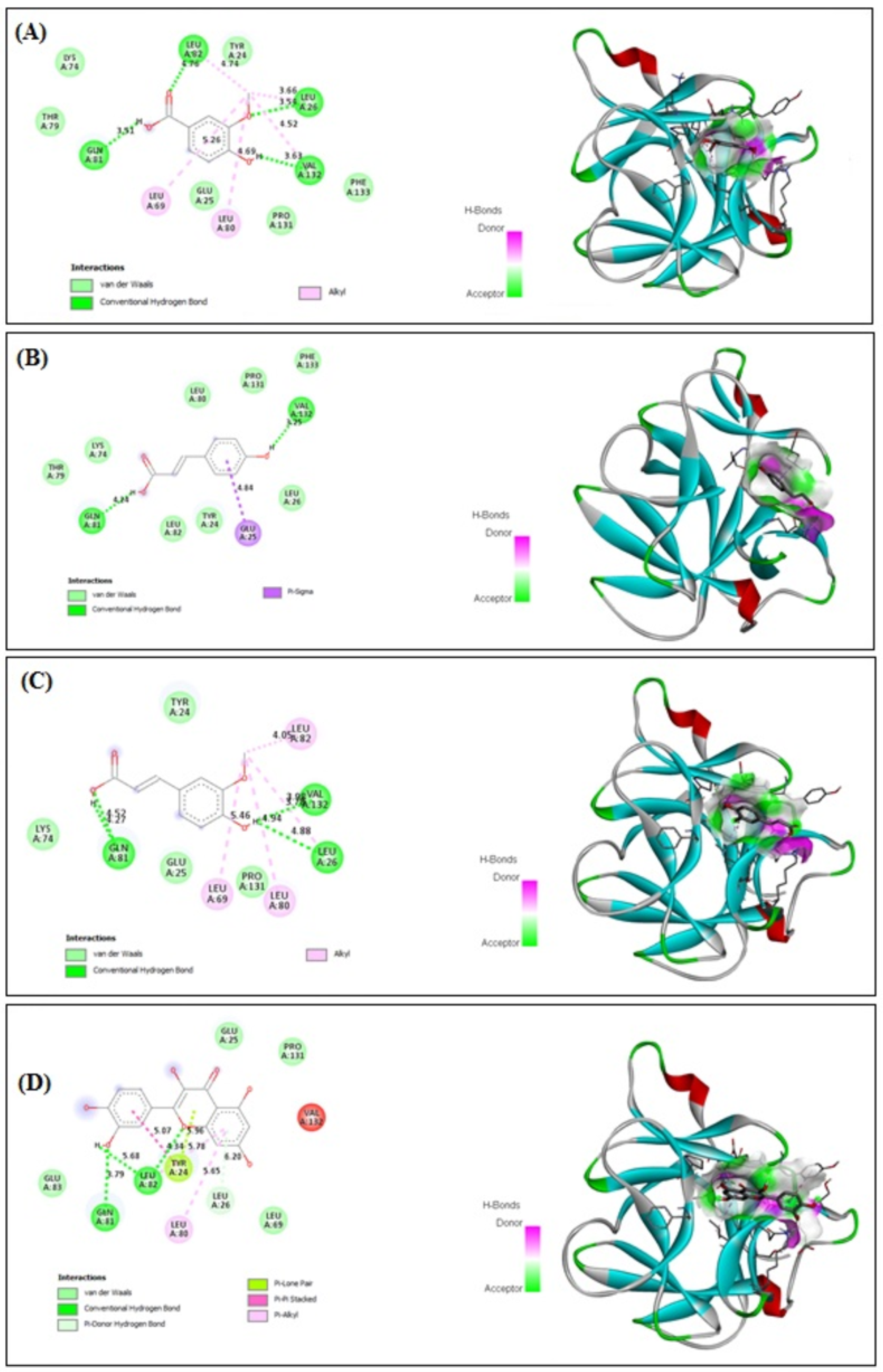
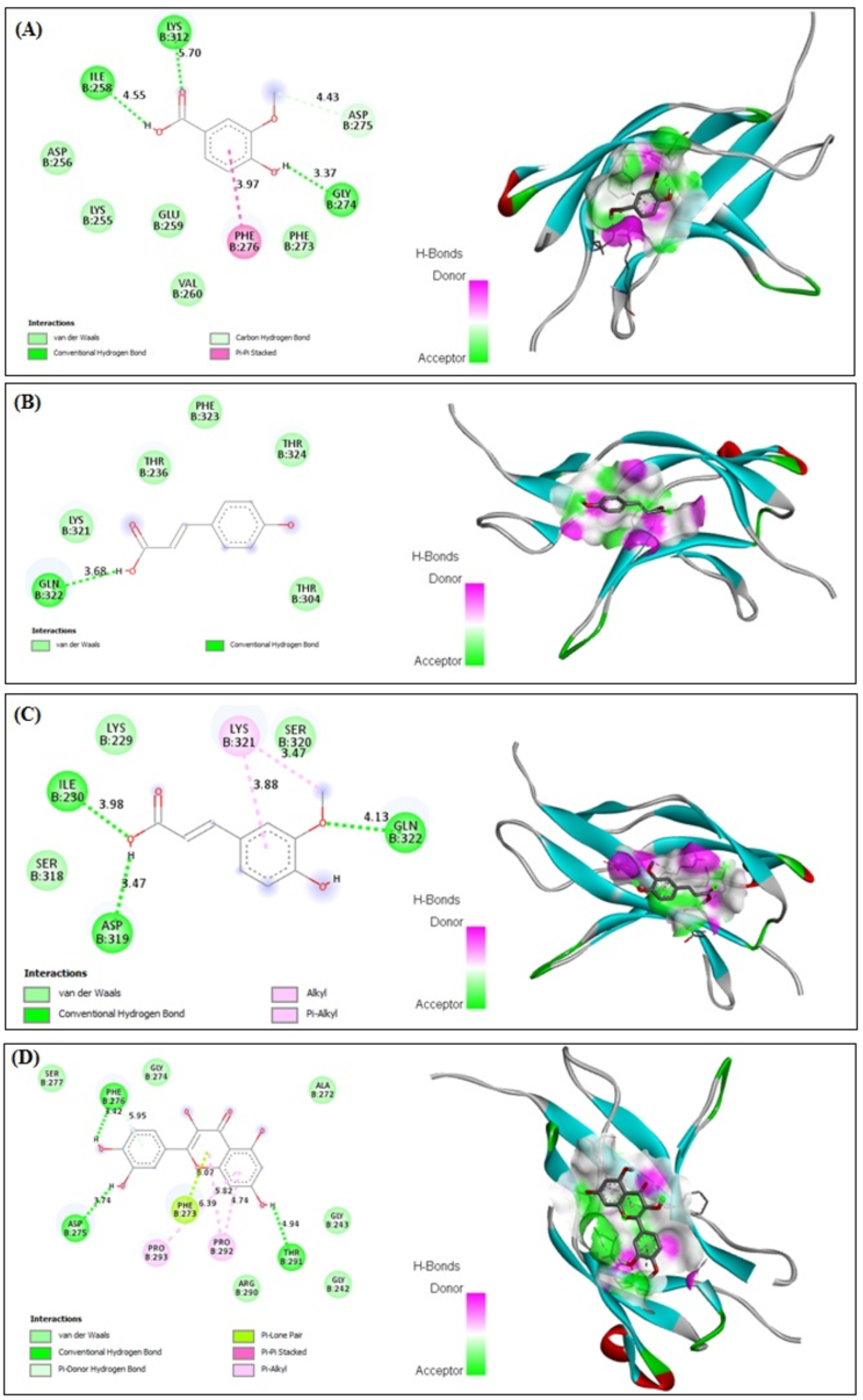
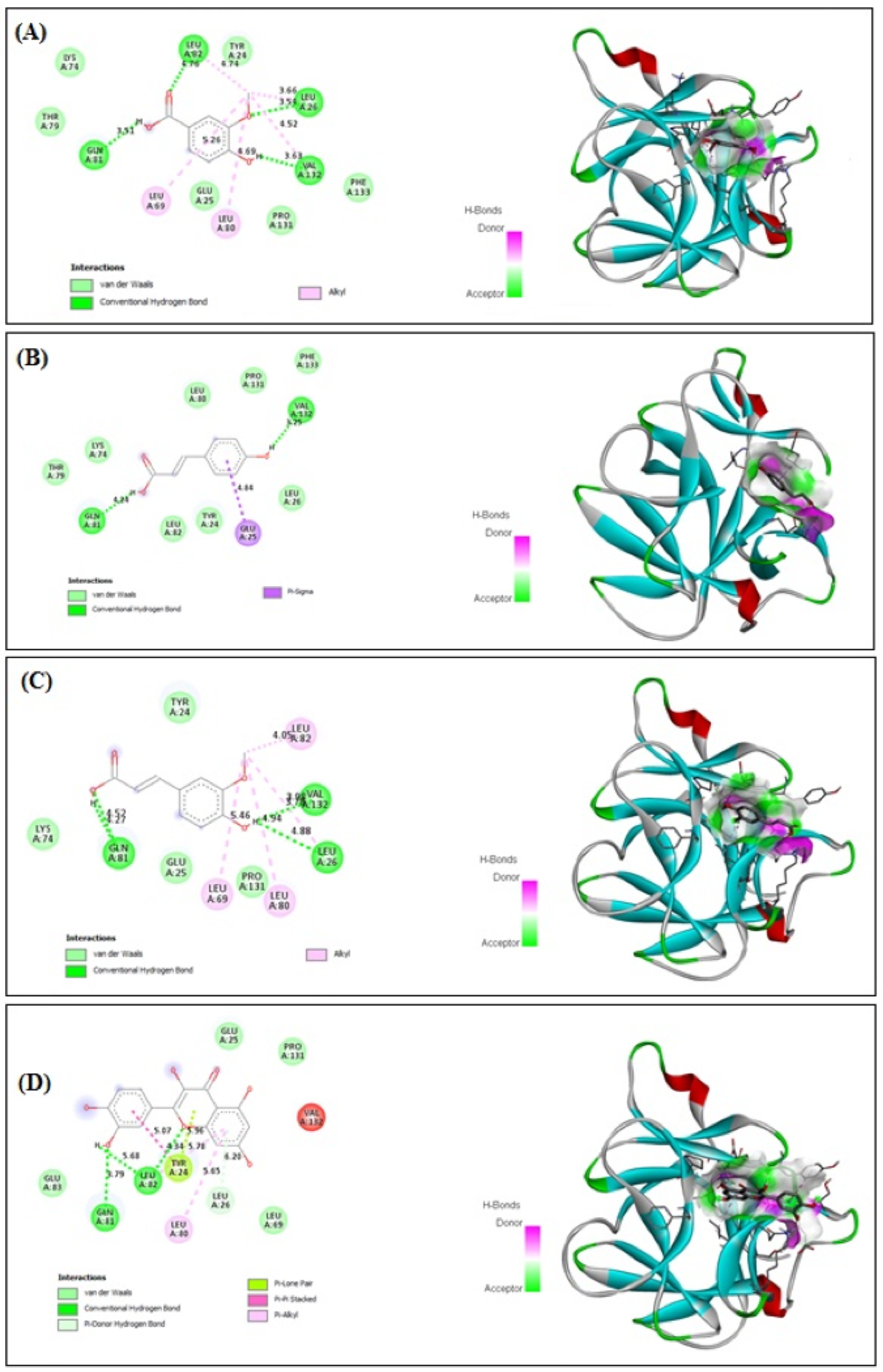
3.10. In Silico Binding of VA, PCA, FA and QRN with NF-κB and IL-1β Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, P.D.; Barri, Y.; Shah, S.V. Oxidant mechanisms in gentamicin nephrotoxicity. Ren. Fail. 1999, 21, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Vysakh, A.; Abhilash, S.; Kuriakose, J.; Midhun, S.J.; Jyothis, M.; Latha, M. Protective effect of Rotula aquatica Lour against gentamicin induced oxidative stress and nephrotoxicity in Wistar rats. Biomed. Pharmacother. 2018, 106, 1188–1194. [Google Scholar]
- Ali, F.E.; Hassanein, E.H.; Bakr, A.G.; El-Shoura, E.A.; El-Gamal, D.A.; Mahmoud, A.R.; Abd-Elhamid, T.H. Ursodeoxycholic acid abrogates gentamicin-induced hepatotoxicity in rats: Role of NF-κB-p65/TNF-α, Bax/Bcl-xl/Caspase-3, and eNOS/iNOS pathways. Life Sci. 2020, 254, 117760. [Google Scholar] [CrossRef]
- Khalil, H.E.; Abdelwahab, M.F.; Emeka, P.M.; Badger-Emeka, L.I.; Abdel Hafez, S.M.N.; AlYahya, K.A.; Ahmed, A.-S.F.; Anter, A.F.; Abdel-Wahab, N.M.; Matsunami, K.; et al. Chemical Composition and Valorization of Broccoli Leaf By-Products (Brassica oleracea L. Variety: Italica) to Ameliorate Reno-Hepatic Toxicity Induced by Gentamicin in Rats. Appl. Sci. 2022, 12, 6903. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Abd El-Ghafar, O.A.; Alzoghaibi, M.A.; Hassanein, E.H. Agomelatine prevents gentamicin nephrotoxicity by attenuating oxidative stress and TLR-4 signaling, and upregulating PPARγ and SIRT1. Life Sci. 2021, 278, 119600. [Google Scholar] [CrossRef]
- Liebman, S.E.; Le, T.H. Eat your broccoli: Oxidative stress, nrf2, and sulforaphane in chronic kidney disease. Nutrients 2021, 13, 266. [Google Scholar] [CrossRef] [PubMed]
- Stickel, F.; Schuppan, D. Herbal medicine in the treatment of liver diseases. Dig. Liver Dis. 2007, 39, 293–304. [Google Scholar] [CrossRef]
- Tlili, N.; Feriani, A.; Saadoui, E.; Nasri, N.; Khaldi, A. Capparis spinosa leaves extract: Source of bioantioxidants with nephroprotective and hepatoprotective effects. Biomed. Pharmacother. 2017, 87, 171–179. [Google Scholar] [CrossRef]
- Kiliś-Pstrusińska, K.; Wiela-Hojeńska, A. Nephrotoxicity of Herbal Products in Europe—A Review of an Underestimated Problem. Int. J. Mol. Sci. 2021, 22, 4132. [Google Scholar] [CrossRef]
- Vanholder, R.; Massy, Z.; Argiles, A.; Spasovski, G.; Verbeke, F.; Lameire, N. Chronic kidney disease as cause of cardiovascular morbidity and mortality. Nephrol. Dial. Transplant. 2005, 20, 1048–1056. [Google Scholar] [CrossRef]
- Aranaz, P.; Navarro-Herrera, D.; Romo-Hualde, A.; Zabala, M.; López-Yoldi, M.; González-Ferrero, C.; Gil, A.G.; Martinez, J.A.; Vizmanos, J.L.; Milagro, F.I. Broccoli extract improves high fat diet-induced obesity, hepatic steatosis and glucose intolerance in Wistar rats. J. Funct. Foods 2019, 59, 319–328. [Google Scholar] [CrossRef]
- Vasanthi, H.R.; Mukherjee, S.; Das, D.K. Potential health benefits of broccoli-a chemico-biological overview. Mini-Rev. Med. Chem. 2009, 9, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Utoiu, E.; Oancea, A.; Gaspar, A.; Seciu, A.-M.; Ștefan, L.M.; Coroiu, V.; Craciunescu, O.; Badiu, C.D.; Oancea, F. Selenium biofortification treatment of cauliflower enhances their content in chemopreventive compounds and in vitro antitumoral activity. Sci. Bulletin. Ser. F. Biotechnol. 2017, 21, 33–40. [Google Scholar]
- Schmidt, R.; Acarkan, A.; Boivin, K. Comparative structural genomics in the Brassicaceae family. Plant Physiol. Biochem. 2001, 39, 253–262. [Google Scholar] [CrossRef]
- Wassermann, B.; Rybakova, D.; Müller, C.; Berg, G. Harnessing the microbiomes of Brassica vegetables for health issues. Sci. Rep. 2017, 7, 17649. [Google Scholar] [CrossRef]
- Ahmed, F.A.; Ali, R.F. Bioactive compounds and antioxidant activity of fresh and processed white cauliflower. Biomed. Res. Int. 2013, 2013, 367819. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, Y.; Li, X.; Kang, H. Antioxidant extract from cauliflower leaves effectively improve the stability of pork patties during refrigerated storage. J. Food Process. Preserv. 2020, 44, e14510. [Google Scholar] [CrossRef]
- Drabińska, N.; Jeż, M.; Nogueira, M. Variation in the Accumulation of Phytochemicals and Their Bioactive Properties among the Aerial Parts of Cauliflower. Antioxidants 2021, 10, 1597. [Google Scholar] [CrossRef]
- Li, Z.; Lee, H.W.; Liang, X.; Liang, D.; Wang, Q.; Huang, D.; Ong, C.N. Profiling of phenolic compounds and antioxidant activity of 12 cruciferous vegetables. Molecules 2018, 23, 1139. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef]
- Hashish, E.A.; Elgaml, S.A. Hepatoprotective and nephroprotective effect of curcumin against copper toxicity in rats. Indian J. Clin. Biochem. 2016, 31, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Süzek, H.; Celik, I.; Doğan, A. Nephroprotective hepatoprotective potential and antioxidant role of carob pods (Cerotonia siliqua L) against carbon tetrachloride-induced toxicity in rats. Indian J. Pharm. Educ. Res. 2017, 51, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Arjinajarn, P.; Chueakula, N.; Pongchaidecha, A.; Jaikumkao, K.; Chatsudthipong, V.; Mahatheeranont, S.; Norkaew, O.; Chattipakorn, N.; Lungkaphin, A. Anthocyanin-rich Riceberry bran extract attenuates gentamicin-induced hepatotoxicity by reducing oxidative stress, inflammation and apoptosis in rats. Biomed. Pharmacother. 2017, 92, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Llorach, R.; Espín, J.C.; Tomás-Barberán, F.A.; Ferreres, F. Valorization of cauliflower (Brassica oleracea L. var. botrytis) by-products as a source of antioxidant phenolics. J. Agric. Food Chem. 2003, 51, 2181–2187. [Google Scholar]
- Khalil, H.E.; Mohamed, M.E.; Morsy, M.A.; Kandeel, M. Flavonoid and phenolic compounds from Carissa macrocarpa: Molecular docking and cytotoxicity studies. Pharmacogn. Mag. 2018, 14, 304–310. [Google Scholar] [CrossRef]
- Khalil, H.E.; Al Ahmed, A. Phytochemical Analysis and Free Radical Scavenging Activity of Carthamus oxyacantha growing in Saudi Arabia: A Comparative Study. Int. J. Pharm. Sci. Rev. Res. 2017, 45, 51–55. [Google Scholar]
- Emeka, P.M.; Rasool, S.T.; Morsy, M.A.; Islam, M.I.H.; Chohan, M.S. Protective effects of lutein against vancomycin-induced acute renal injury in mice via upregulation of peroxisome proliferator-activated receptor gamma/nuclear factor erythroid 2-related factor 2 and inhibition nuclear factor-kappaB/caspase 3. Korean J. Physiol. Pharmacol. 2021, 25, 321–331. [Google Scholar] [CrossRef]
- Chatterjee, A.; Khanra, R.; Chattopadhyay, M.; Ghosh, S.; Sahu, R.; Nandi, G.; Maji, H.S.; Chakraborty, P. Pharmacological studies of rhizomes of extract of Cyperus tegetum, emphasized on anticancer, anti-inflammatory and analgesic activity. J. Ethnopharmacol. 2022, 289, 115035. [Google Scholar] [CrossRef]
- Khalil, H.E.; Abdelwahab, M.F.; Emeka, P.M.; Badger-Emeka, L.I.; Thirugnanasambantham, K.; Ibrahim, H.-I.M.; Naguib, S.M.; Matsunami, K.; Abdel-Wahab, N.M. Ameliorative Effect of Ocimum forskolei Benth on Diabetic, Apoptotic, and Adipogenic Biomarkers of Diabetic Rats and 3T3-L1 Fibroblasts Assisted by In Silico Approach. Molecules 2022, 27, 2800. [Google Scholar] [CrossRef]
- Beshay, O.N.; Ewees, M.G.; Abdel-Bakky, M.S.; Hafez, S.M.N.A.; Abdelrehim, A.B.; Bayoumi, A.M. Resveratrol reduces gentamicin-induced EMT in the kidney via inhibition of reactive oxygen species and involving TGF-β/Smad pathway. Life Sci. 2020, 258, 118178. [Google Scholar] [CrossRef]
- Ibrahim, H.-I.M.; Darrag, H.M.; Alhajhoj, M.R.; Khalil, H.E. Biomolecule from Trigonella stellata from Saudi Flora to Suppress Osteoporosis via Osteostromal Regulations. Plants 2020, 9, 1610. [Google Scholar] [CrossRef] [PubMed]
- Khalil, H.E.; Ibrahim, H.-I.M.; Ahmed, E.A.; Emeka, P.M.; Alhaider, I.A. Orientin, a Bio-Flavonoid from Trigonella hamosa L., Regulates COX-2/PGE-2 in A549 Cell Lines via miR-26b and miR-146a. Pharmaceuticals 2022, 15, 154. [Google Scholar] [CrossRef] [PubMed]
- Khalil, H.E.; Ibrahim, H.-I.M.; Darrag, H.M.; Matsunami, K. Insight into Analysis of Essential Oil from Anisosciadium lanatum Boiss.—Chemical Composition, Molecular Docking, and Mitigation of Hepg2 Cancer Cells through Apoptotic Markers. Plants 2021, 11, 66. [Google Scholar] [CrossRef]
- Chang, S.-W.; Kim, K.-H.; Lee, I.-K.; Choi, S.-U.; Ryu, S.-Y.; Lee, K.-R. Phytochemical constituents of Bistorta manshuriensis. Nat. Prod. Sci. 2009, 15, 234–240. [Google Scholar]
- Rho, T.; Yoon, K.D. Chemical constituents of Nelumbo nucifera seeds. Nat. Prod. Sci. 2017, 23, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Elgazzar, D.; Aboubakr, M.; Bayoumi, H.; Ibrahim, A.N.; Sorour, S.M.; El-Hewaity, M.; Elsayed, A.M.; Shehata, S.A.; Bayoumi, K.A.; Alsieni, M. Tigecycline and Gentamicin-Combined Treatment Enhances Renal Damage: Oxidative Stress, Inflammatory Reaction, and Apoptosis Interplay. Pharmaceuticals 2022, 15, 736. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.; Elkomy, A.; Alkafafy, M.; Elkammar, R.; El-Shafey, A.; Soliman, A.; Aboubakr, M. Testicular toxicity of cisplatin in rats: Ameliorative effect of lycopene and N-acetylcysteine. Environ. Sci. Pollut. Res. 2022, 29, 24077–24084. [Google Scholar] [CrossRef]
- Laaroussi, H.; Bakour, M.; Ousaaid, D.; Ferreira-Santos, P.; Genisheva, Z.; El Ghouizi, A.; Aboulghazi, A.; Teixeira, J.A.; Lyoussi, B. Protective effect of honey and propolis against gentamicin-induced oxidative stress and hepatorenal damages. Oxid. Med. Cell. Longev. 2021, 2021, 9719906. [Google Scholar] [CrossRef]
- Banday, A.A.; Farooq, N.; Priyamvada, S.; Yusufi, A.N.; Khan, F. Time dependent effects of gentamicin on the enzymes of carbohydrate metabolism, brush border membrane and oxidative stress in rat kidney tissues. Life Sci. 2008, 82, 450–459. [Google Scholar] [CrossRef]
- Kalayarasan, S.; Prabhu, P.N.; Sriram, N.; Manikandan, R.; Arumugam, M.; Sudhandiran, G. Diallyl sulfide enhances antioxidants and inhibits inflammation through the activation of Nrf2 against gentamicin-induced nephrotoxicity in Wistar rats. Eur. J. Pharmacol. 2009, 606, 162–171. [Google Scholar] [CrossRef]
- Köksal, E.; Gülçin, İ. Antioxidant activity of cauliflower (Brassica oleracea L.). Turk. J. Agric. For. 2008, 32, 65–78. [Google Scholar]
- Hasanvand, A.; Kharazmkia, A.; Mir, S.; Khorramabadi, R.M.; Darabi, S. Ameliorative effect of ferulic acid on gentamicininduced nephrotoxicity in a rat model; role of antioxidant effects. J. Ren. Inj. Prev. 2018, 7, 73–77. [Google Scholar] [CrossRef]
- Hakyemez, I.N.; Cevizci, M.N.; Aksoz, E.; Yilmaz, K.; Uysal, S.; Altun, E. Protective effects of p-coumaric acid against gentamicin-induced nephrotoxicity in rats. Drug Chem. Toxicol. 2021, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Raheem, I.T.; Abdel-Ghany, A.A.; Mohamed, G.A. Protective effect of quercetin against gentamicin-induced nephrotoxicity in rats. Biol. Pharm. Bull. 2009, 32, 61–67. [Google Scholar] [CrossRef]
- Andreev, E.; Koopman, M.; Arisz, L. A rise in plasma creatinine that is not a sign of renal failure: Which drugs can be responsible? J. Intern. Med. 1999, 246, 247–252. [Google Scholar] [CrossRef]
- Tanaka, S.; Ninomiya, T.; Taniguchi, M.; Tokumoto, M.; Masutani, K.; Ooboshi, H.; Kitazono, T.; Tsuruya, K. Impact of blood urea nitrogen to creatinine ratio on mortality and morbidity in hemodialysis patients: The Q-Cohort Study. Sci. Rep. 2017, 7, 14901. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Jin, W.W.; Huang, M.; Ji, H.; Capen, D.E.; Xia, Y.; Yuan, J.; Păunescu, T.G.; Lu, H.A.J. Gentamicin-induced acute kidney injury in an animal model involves programmed necrosis of the collecting duct. J. Am. Soc. Nephrol. 2020, 31, 2097–2115. [Google Scholar] [CrossRef]
- Clemente Plaza, N.; Reig García-Galbis, M.; Martínez-Espinosa, R.M. Effects of the Usage of l-Cysteine (l-Cys) on Human Health. Molecules 2018, 23, 575. [Google Scholar] [CrossRef]
- Al-Kenanny, E.; Al-Hayaly, L.; Al-Badrany, A. Protective effect of arabic gum on liver injury experimentally induced by gentamycin in mice. Kufa J. Vet. Med. Sci. 2012, 3, 174–189. [Google Scholar]
- Galaly, S.; Ahmed, O.; Mahmoud, A. Thymoquinone and curcumin prevent gentamicin-induced liver injury by attenuating oxidative stress, inflammation and apoptosis. J. Physiol. Pharmacol. 2014, 65, 823–832. [Google Scholar]
- Jaikumkao, K.; Pongchaidecha, A.; Thongnak, L.-o.; Wanchai, K.; Arjinajarn, P.; Chatsudthipong, V.; Chattipakorn, N.; Lungkaphin, A. Amelioration of renal inflammation, endoplasmic reticulum stress and apoptosis underlies the protective effect of low dosage of atorvastatin in gentamicin-induced nephrotoxicity. PLoS ONE 2016, 11, e0164528. [Google Scholar]
- Zheng, R.; Tan, Y.; Gu, M.; Kang, T.; Zhang, H.; Guo, L. N-acetyl cysteine inhibits lipopolysaccharide-mediated synthesis of interleukin-1β and tumor necrosis factor-α in human periodontal ligament fibroblast cells through nuclear factor-kappa B signaling. Medicine 2019, 98, e17126. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Hershko, D.D.; Robb, B.W.; Wray, C.J.; Hasselgren, P.-O. IL-1β stimulates IL-6 production in cultured skeletal muscle cells through activation of MAP kinase signaling pathway and NF-κB. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, 1249–1254. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Larocca, M.; Perna, A.M.; Simonetti, A.; Gambacorta, E.; Iannuzzi, A.; Perucatti, A.; Rossano, R. Antioxidant and anti-inflammatory effects of cauliflower leaf powder-enriched diet against LPS induced toxicity in rabbits. Food Funct. 2017, 8, 3288–3296. [Google Scholar] [CrossRef]
- Kalkan, Y.; Kapakin, K.A.T.; Kara, A.; Atabay, T.; Karadeniz, A.; Simsek, N.; Karakus, E.; Can, I.; Yildirim, S.; Ozkanlar, S. Protective effect of Panax ginseng against serum biochemical changes and apoptosis in kidney of rats treated with gentamicin sulphate. J. Mol. Histol. 2012, 43, 603–613. [Google Scholar] [CrossRef]
- Ezema, A.S.; Ihedioha, T.E.; Num-Adom, S.M.; Ihedioha, J.I. Alterations in serum activity of hepatocellular enzymes, levels of liver function markers, and liver histology of dogs given high (nephrotoxic) doses of gentamicin. Comp. Clin. Path. 2022, 31, 115–122. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, H.E.; Abdelwahab, M.F.; Emeka, P.M.; Badger-Emeka, L.I.; Ahmed, A.-S.F.; Anter, A.F.; Abdel Hafez, S.M.N.; AlYahya, K.A.; Ibrahim, H.-I.M.; Thirugnanasambantham, K.; et al. Brassica oleracea L. var. botrytis Leaf Extract Alleviates Gentamicin-Induced Hepatorenal Injury in Rats—Possible Modulation of IL-1β and NF-κB Activity Assisted with Computational Approach. Life 2022, 12, 1370. https://doi.org/10.3390/life12091370
Khalil HE, Abdelwahab MF, Emeka PM, Badger-Emeka LI, Ahmed A-SF, Anter AF, Abdel Hafez SMN, AlYahya KA, Ibrahim H-IM, Thirugnanasambantham K, et al. Brassica oleracea L. var. botrytis Leaf Extract Alleviates Gentamicin-Induced Hepatorenal Injury in Rats—Possible Modulation of IL-1β and NF-κB Activity Assisted with Computational Approach. Life. 2022; 12(9):1370. https://doi.org/10.3390/life12091370
Chicago/Turabian StyleKhalil, Hany Ezzat, Miada F. Abdelwahab, Promise Madu Emeka, Lorina I. Badger-Emeka, Al-Shaimaa F. Ahmed, Aliaa F. Anter, Sara Mohamed Naguib Abdel Hafez, Khalid A. AlYahya, Hairul-Islam Mohamed Ibrahim, Krishnaraj Thirugnanasambantham, and et al. 2022. "Brassica oleracea L. var. botrytis Leaf Extract Alleviates Gentamicin-Induced Hepatorenal Injury in Rats—Possible Modulation of IL-1β and NF-κB Activity Assisted with Computational Approach" Life 12, no. 9: 1370. https://doi.org/10.3390/life12091370
APA StyleKhalil, H. E., Abdelwahab, M. F., Emeka, P. M., Badger-Emeka, L. I., Ahmed, A.-S. F., Anter, A. F., Abdel Hafez, S. M. N., AlYahya, K. A., Ibrahim, H.-I. M., Thirugnanasambantham, K., Matsunami, K., & Ibrahim Selim, A. H. (2022). Brassica oleracea L. var. botrytis Leaf Extract Alleviates Gentamicin-Induced Hepatorenal Injury in Rats—Possible Modulation of IL-1β and NF-κB Activity Assisted with Computational Approach. Life, 12(9), 1370. https://doi.org/10.3390/life12091370