

Curcumin Modulates Oxidative Stress, Fibrosis, and Apoptosis in Drug-Resistant Cancer Cell Lines

 , ,
, ,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
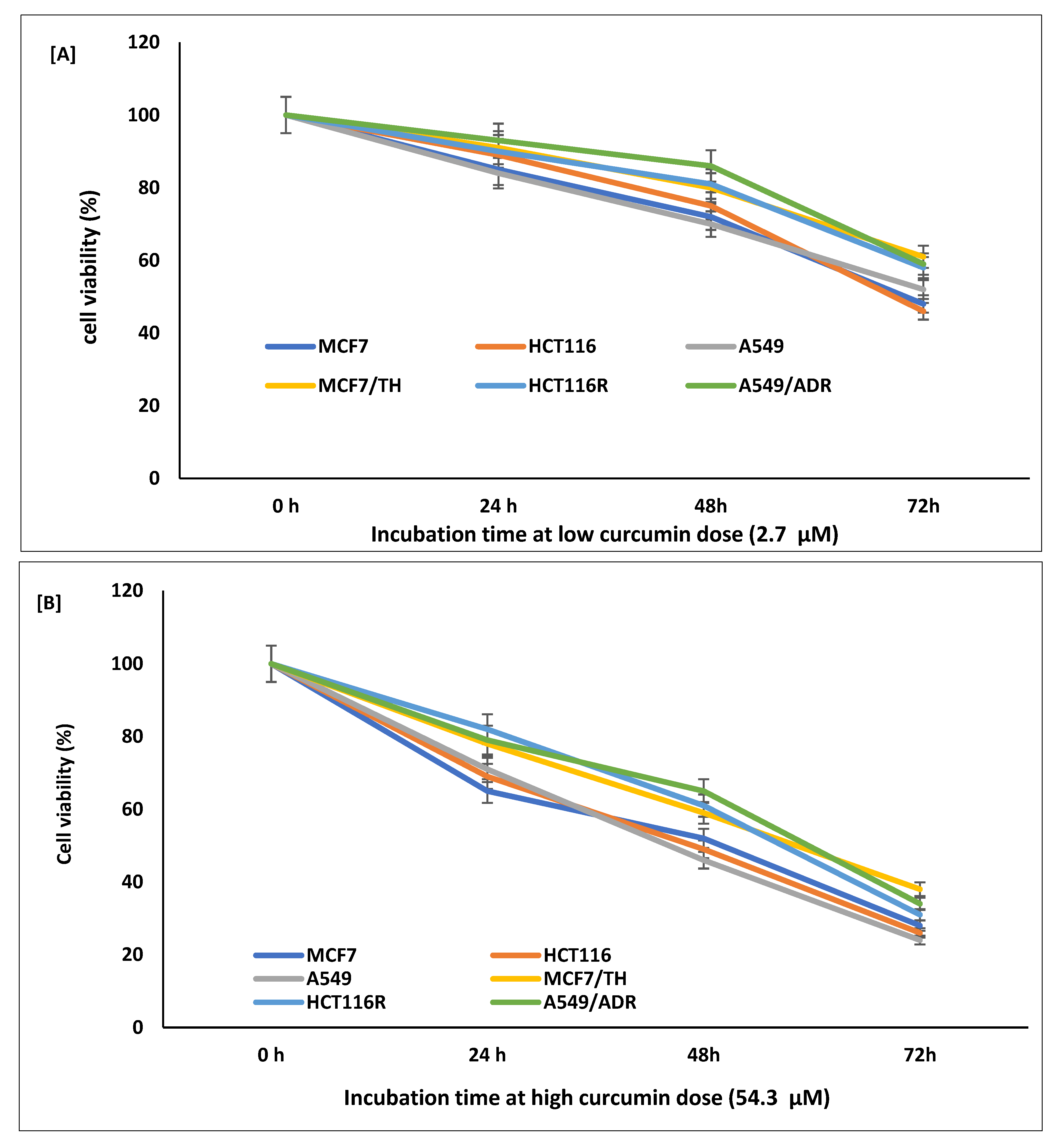
2.1. Cytotoxicity and Inhibitory Effect of Curcumin
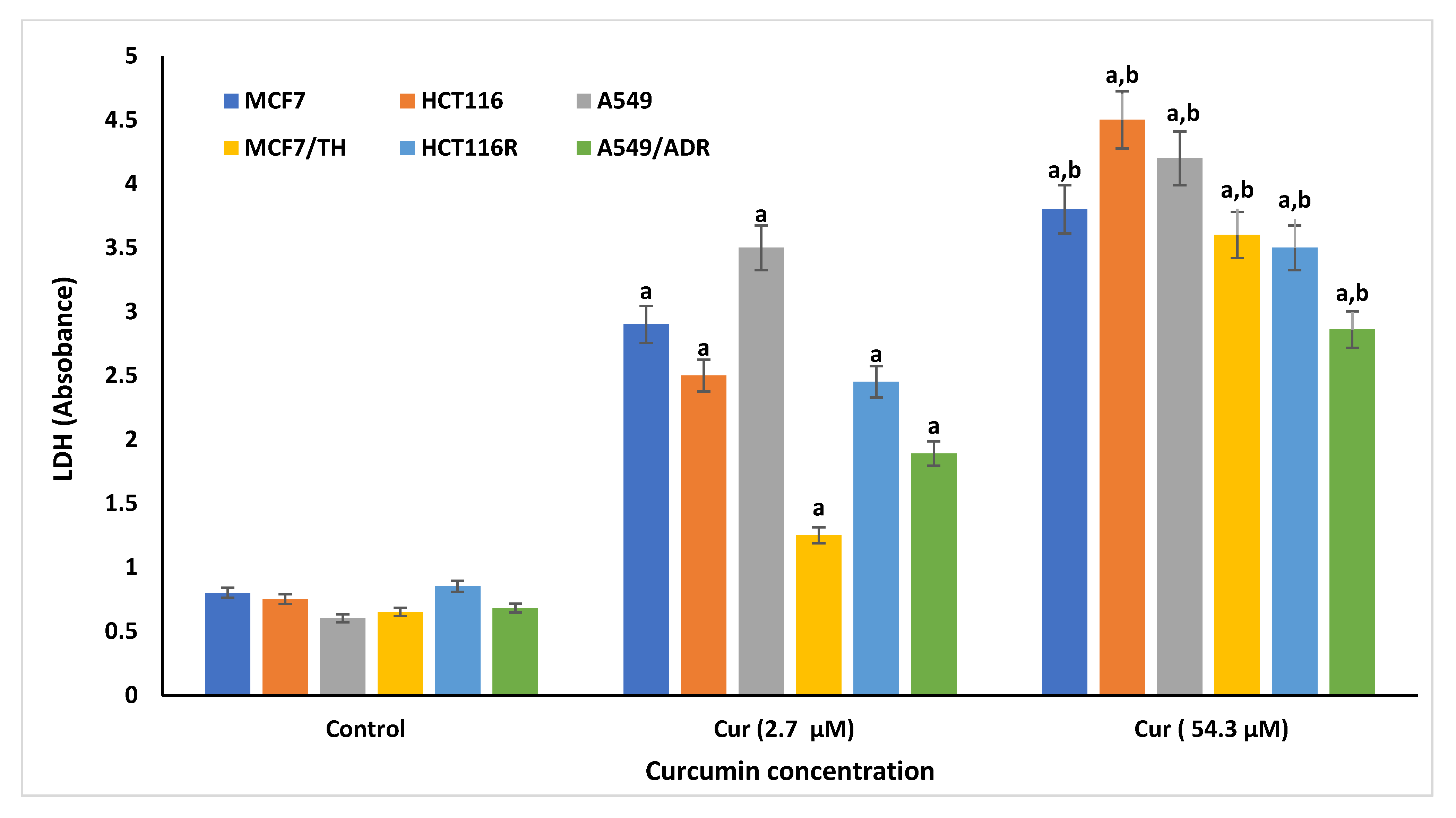
2.2. LDH Activity
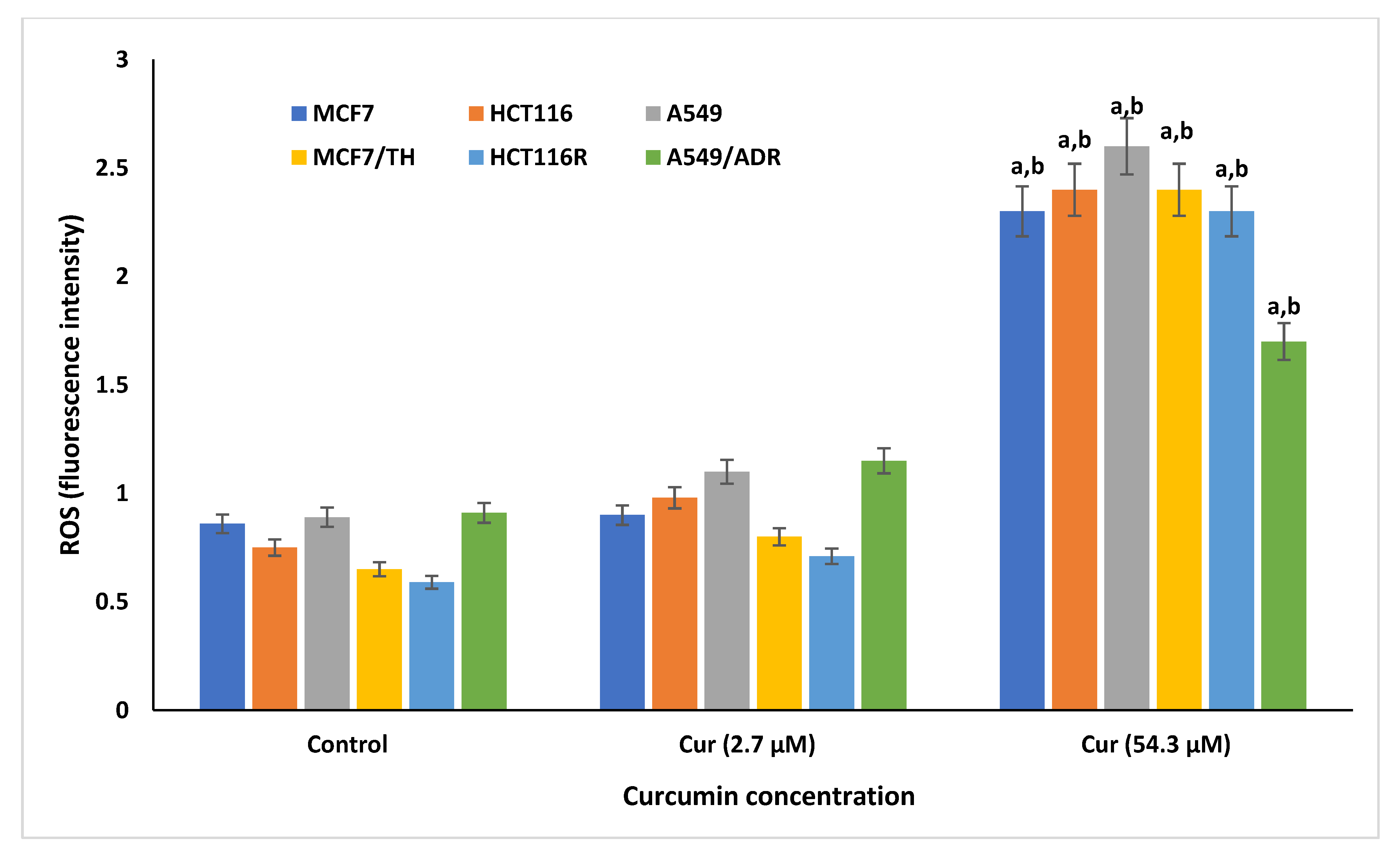
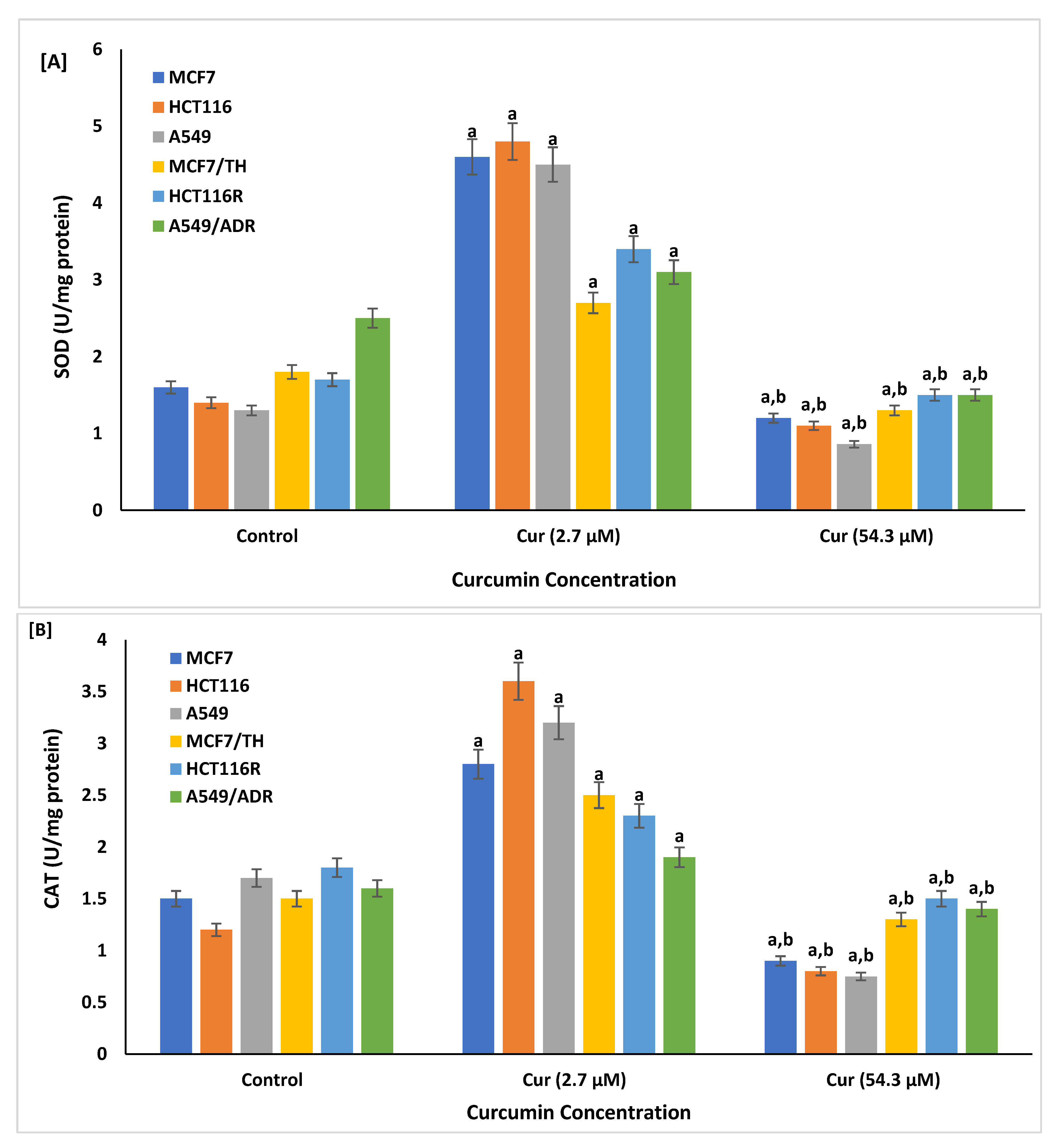
2.3. Effect of Curcumin on the Cellular Reactive Oxygen Species and Antioxdant Enzymes
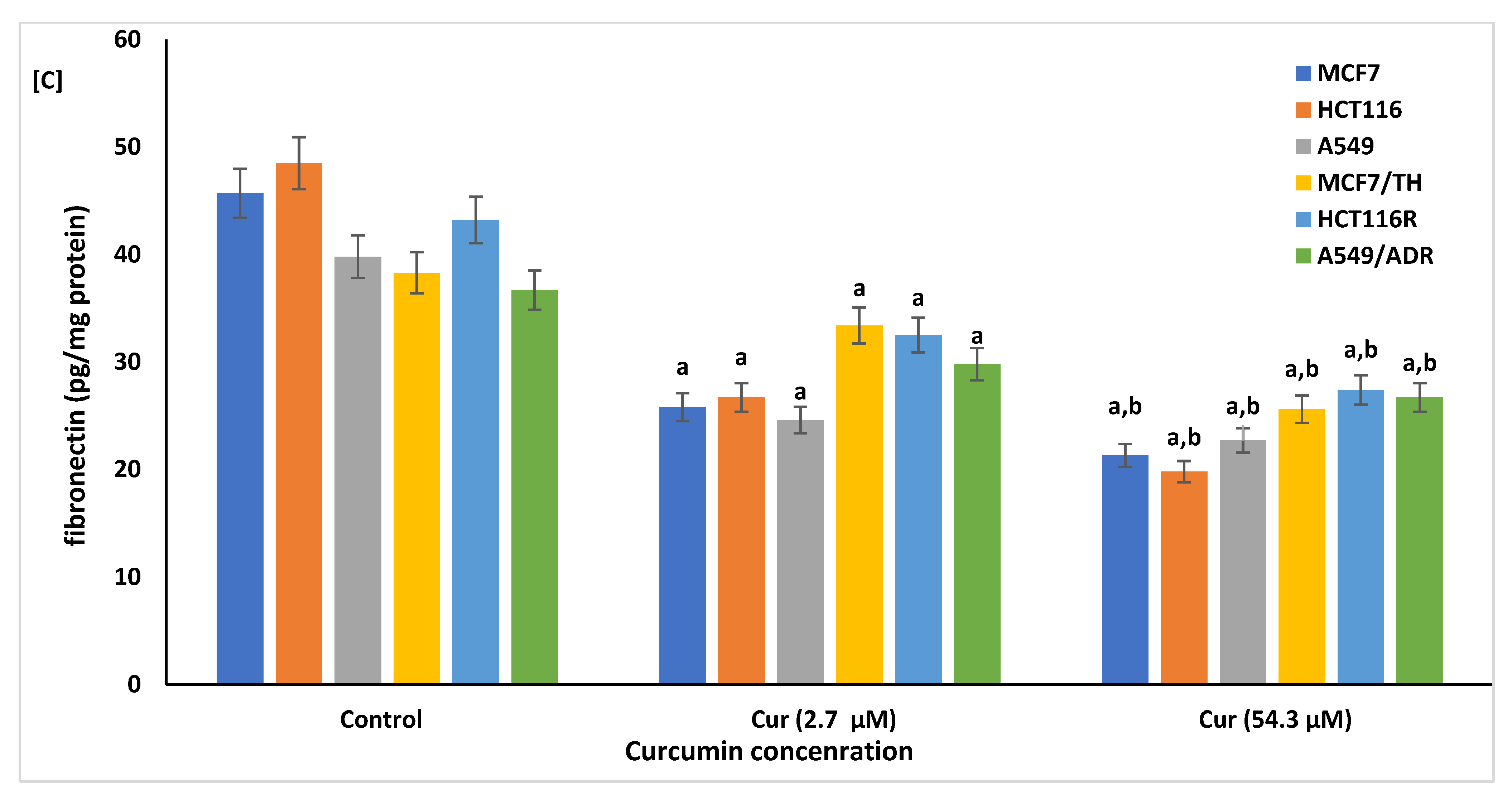
2.4. Effect of Curcumin on the Cellular Fibrosis
2.5. Effect of Curcumin on Expression of Sirtuin1 (SIRT1)
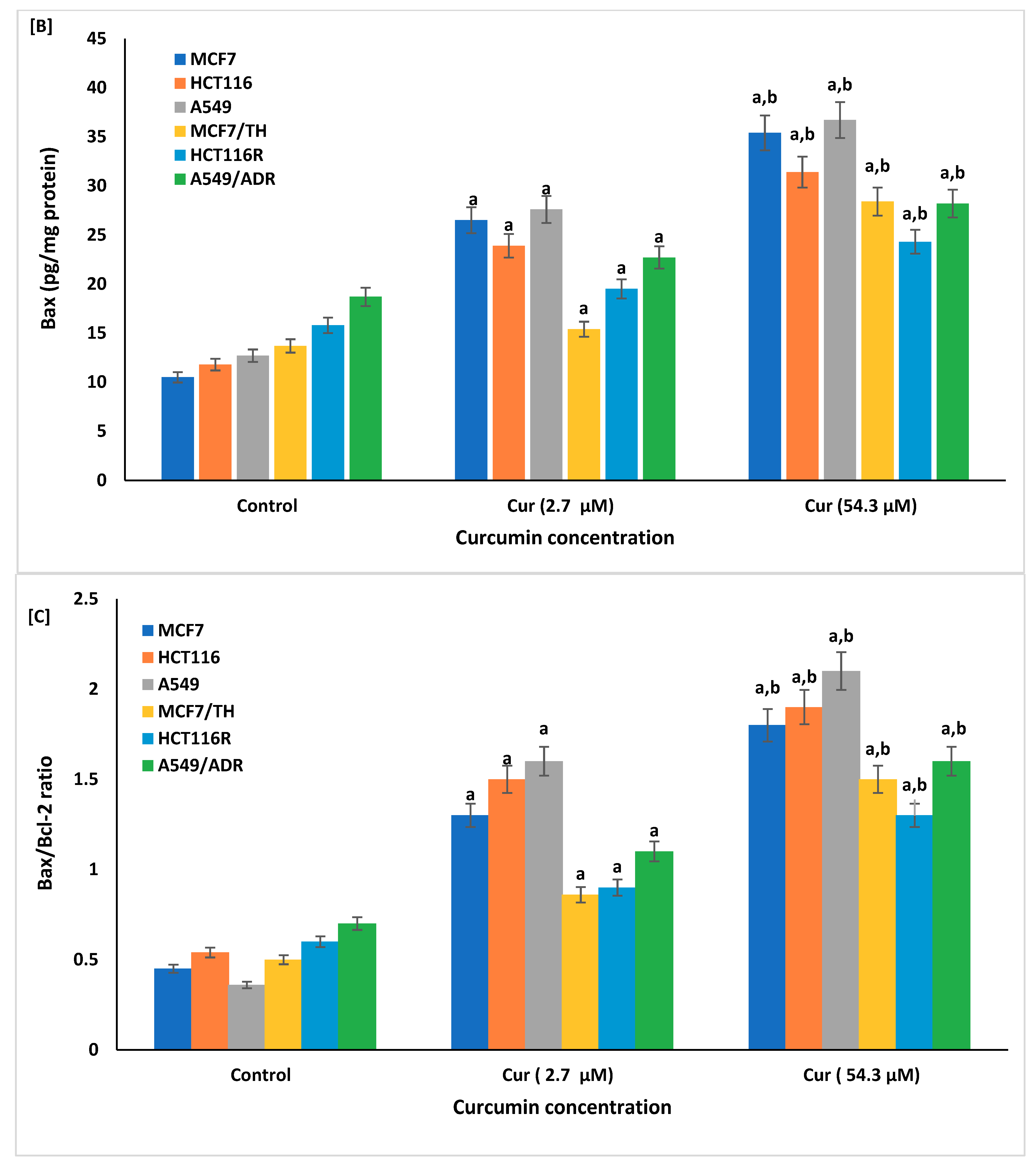
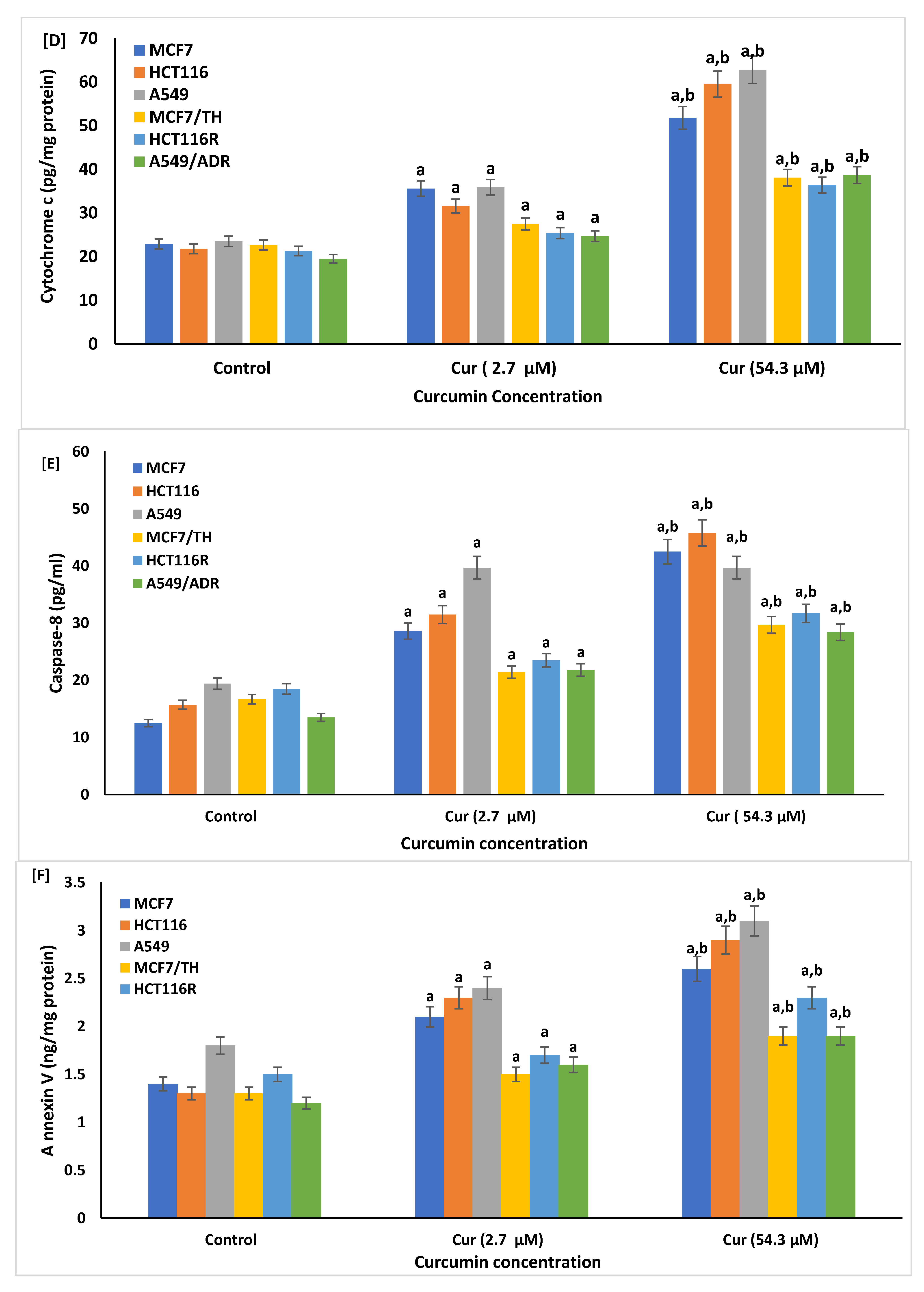
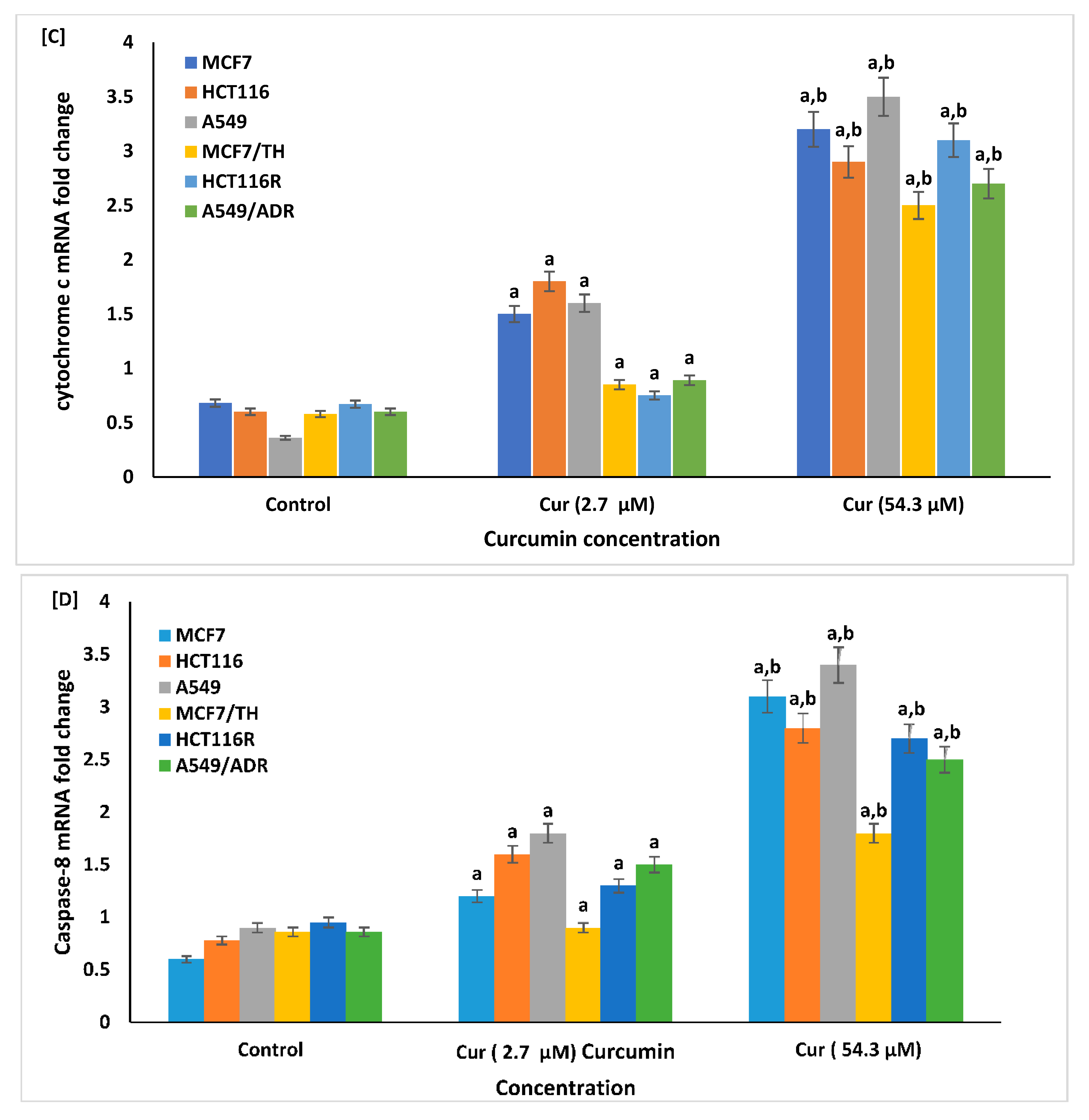
2.6. Effect of Curcumin on the Cellular Apoptosis
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of Curcumin
4.3. Cell Lines
4.4. Cell Culture
4.5. Cell Viability Assay
4.6. Biochemical and Cellular Investigations
4.6.1. Lactate Dehydrogenase (LDH) Assay
4.6.2. Assessment of Cellular Oxidative Stress
4.6.3. Assessment of Antioxidant Enzymes
4.6.4. Assessment of Cellular Fibrosis
4.7. Apoptotic Markers
4.7.1. ELISA Immunoassay Analysis of Bcl-2, Cytochrome c, and Caspase-8
4.7.2. Assay of Sirtuin1 (SIRT1), Bax, and Anti-Annexin V
4.8. Real-Time Quantitative (RT-PCR) Analysis of the mRNA Expression of Apoptotic Markers and SIRT1
4.8.1. RNA Extraction and cDNA Preparation
4.8.2. Real-Time RT-PCR Analysis of Apoptotic Markers and SIRT1
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.P.; Pandotra, P.; Sharma, R.; Kushwaha, M.; Gupta, S. Marine resource: A promising future for anticancer drugs. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2013; Volume 40, pp. 229–325. [Google Scholar]
- Keyvani-Ghamsari, S.; Khorsandi, K.; Gul, A. Curcumin effect on cancer cells’ multidrug resistance: An update. Phytother. Res. 2020, 34, 2534–2556. [Google Scholar] [CrossRef] [PubMed]
- Kumari, N.; Dwarakanath, B.; Das, A.; Bhatt, A.N. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumor Biol. 2016, 37, 11553–11572. [Google Scholar] [CrossRef] [PubMed]
- Kozakai, N.; Kikuchi, E.; Hasegawa, M.; Suzuki, E.; Ide, H.; Miyajima, A.; Horiguchi, Y.; Nakashima, J.; Umezawa, K.; Shigematsu, N. Enhancement of radiosensitivity by a unique novel NF-κB inhibitor, DHMEQ, in prostate cancer. Br. J. Cancer 2012, 107, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Naugler, W.E.; Karin, M. NF-κB and cancer—identifying targets and mechanisms. Curr. Opin. Genet. Dev. 2008, 18, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.; Rose-John, S.; Garbers, C. Interleukin-6 and its receptors: A highly regulated and dynamic system. Cytokine 2014, 70, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Woo, S.; Kang, J.; Kim, K.; Shin, H.; Gwak, H.; Park, S.; Chwae, Y. NF-κB and STAT3 cooperatively induce IL6 in starved cancer cells. Oncogene 2012, 31, 3467–3481. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Wang, J.-Q.; Assaraf, Y.G.; Ren, L.; Gupta, P.; Wei, L.; Ashby, C.R., Jr.; Yang, D.-H.; Chen, Z.-S. Modulating ROS to overcome multidrug resistance in cancer. Drug Resist. Updates 2018, 41, 1–25. [Google Scholar] [CrossRef]
- Gatenby, R.; Brown, J. The evolution and ecology of resistance in cancer therapy. Cold Spring Harb. Perspect. Med. 2018, 8, a033415. [Google Scholar] [CrossRef]
- Wang, X.; Fang, Y.; Sun, W.; Xu, Z.; Zhang, Y.; Wei, X.; Ding, X.; Xu, Y. Endocrinotherapy resistance of prostate and breast cancer: Importance of the NF-κB pathway. Int. J. Oncol. 2020, 56, 1064–1074. [Google Scholar] [CrossRef]
- Yang, C.; Ma, X.; Wang, Z.; Zeng, X.; Hu, Z.; Ye, Z.; Shen, G. Curcumin induces apoptosis and protective autophagy in castration-resistant prostate cancer cells through iron chelation. Drug Des. Dev. Ther. 2017, 11, 431. [Google Scholar] [CrossRef]
- Hamed, A.; Abdel-Razek, A.S.; Frese, M.; Stammler, H.G.; El-Haddad, A.F.; Ibrahim, T.; Sewald, N.; Shaaban, M. Terretonin n: A new meroterpenoid from Nocardiopsis sp. Molecules 2018, 23, 299. [Google Scholar] [CrossRef]
- Alibeiki, F.; Jafari, N.; Karimi, M.; Peeri Dogaheh, H. Potent anti-cancer effects of less polar Curcumin analogues on gastric adenocarcinoma and esophageal squamous cell carcinoma cells. Sci. Rep. 2017, 7, 2559. [Google Scholar] [CrossRef]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef]
- Perrone, D.; Ardito, F.; Giannatempo, G.; Dioguardi, M.; Troiano, G.; Lo Russo, L.; De Lillo, A.; Laino, L.; Lo Muzio, L. Biological and therapeutic activities, and anticancer properties of curcumin. Exp. Ther. Med. 2015, 10, 1615–1623. [Google Scholar] [CrossRef]
- Ammon, H.P.; Wahl, M.A. Pharmacology of Curcuma longa. Planta Med. 1991, 57, 1–7. [Google Scholar] [CrossRef]
- Smith, M.R.; Gangireddy, S.R.; Narala, V.R.; Hogaboam, C.M.; Standiford, T.J.; Christensen, P.J.; Kondapi, A.K.; Reddy, R.C. Curcumin inhibits fibrosis-related effects in IPF fibroblasts and in mice following bleomycin-induced lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, L616–L625. [Google Scholar] [CrossRef]
- Nagahama, K.; Utsumi, T.; Kumano, T.; Maekawa, S.; Oyama, N.; Kawakami, J. Discovery of a new function of curcumin which enhances its anticancer therapeutic potency. Sci. Rep. 2016, 6, 30962. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Deb, L.; Prasad, S. Curcumin differs from tetrahydrocurcumin for molecular targets, signaling pathways and cellular responses. Molecules 2015, 20, 185–205. [Google Scholar] [CrossRef]
- Shehzad, A.; Wahid, F.; Lee, Y.S. Curcumin in cancer chemoprevention: Molecular targets, pharmacokinetics, bioavailability, and clinical trials. Arch. Der Pharm. 2010, 343, 489–499. [Google Scholar] [CrossRef]
- Chattopadhyay, I.; Biswas, K.; Bandyopadhyay, U.; Banerjee, R.K. Turmeric and curcumin: Biological actions and medicinal applications. Curr. Sci. 2004, 87, 44–53. [Google Scholar]
- Jaruga, E.; Sokal, A.; Chrul, S.; Bartosz, G. Apoptosis-independent alterations in membrane dynamics induced by curcumin. Exp. Cell Res. 1998, 245, 303–312. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sung, B. Pharmacological basis for the role of curcumin in chronic diseases: An age-old spice with modern targets. Trends Pharmacol. Sci. 2009, 30, 85–94. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: Multitargeting for multiple chronic diseases. Br. J. Pharmacol. 2017, 174, 1325–1348. [Google Scholar] [CrossRef]
- Gerber, A.; Wille, A.; Welte, T.; Ansorge, S.; Bühling, F. Interleukin-6 and transforming growth factor-β1 control expression of cathepsins B and L in human lung epithelial cells. J. Interferon Cytokine Res. 2001, 21, 11–19. [Google Scholar] [CrossRef]
- Jiang, H.-D.; Guan, H.-S. MS80, a novel sulfated oligosaccharide, inhibits pulmonary fibrosis by targeting TGF-β1 both in vitro and in vivo. Acta Pharmacol. Sin. 2009, 30, 973–979. [Google Scholar] [CrossRef]
- Sharma, A.; Thakur, R.; Lingaraju, M.C.; Kumar, D.; Mathesh, K.; Telang, A.G.; Singh, T.U.; Kumar, D. Betulinic acid attenuates renal fibrosis in rat chronic kidney disease model. Biomed. Pharmacother. 2017, 89, 796–804. [Google Scholar] [CrossRef]
- Gabr, S.A.; Alghadir, A.H. Handgrip Strength and Vitamin D as Predictors of Liver Fibrosis and Malnutrition in Chronic Hepatitis C Patients. Dis. Markers 2021, 2021, 6665893. [Google Scholar] [CrossRef]
- Fu, S.; Kurzrock, R. Development of curcumin as an epigenetic agent. Cancer 2010, 116, 4670–4676. [Google Scholar] [CrossRef]
- Rice, J.C.; Ozcelik, H.; Maxeiner, P.; Andrulis, I.; Futscher, B.W. Methylation of the BRCA1 promoter is associated with decreased BRCA1 mRNA levels in clinical breast can-cer specimens. Carcinogenesis 2000, 21, 1761–1765. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.F.; Howell, A.; Sartini, M.; Sotgia, F.; Lisanti, M.P. Downregulation of stromal BRCA1 drives breast cancer tumor growth via upregulation of HIF-1α, autophagy and ketone body production. Cell Cycle 2012, 11, 4167–4173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetoni, A.R.; Paciello, F.; Mezzogori, D.; Rolesi, R.; Eramo, S.L.M.; Paludetti, G.; Troiani, D. Molecular targets for anticancer redox chemotherapy and cisplatin-induced ototoxicity: The role of curcumin on pSTAT3 and Nrf-2 signalling. Br. J. Cancer 2015, 113, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Sivanantham, B.; Sethuraman, S.; Krishnan, U.M. Combinatorial effects of curcumin with an anti-neoplastic agent on head and neck squamous cell carcinoma through the regulation of EGFR-ERK1/2 and apoptotic signaling pathways. ACS Comb. Sci. 2016, 18, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, I.F.; Borné, Y.; Melander, O.; Orho-Melander, M.; Nilsson, J.; Söderholm, M.; Engström, G. FADD (Fas-associated protein with death domain), caspase-3, and caspase-8 and incidence of ischemic stroke. Stroke 2018, 49, 2224–2226. [Google Scholar] [CrossRef]
- Narayan, S. Curcumin, a multi-functional chemopreventive agent, blocks growth of colon cancer cells by targeting β-catenin-mediated transactivation and cell–cell adhesion pathways. J. Mol. Histol. 2004, 35, 301–307. [Google Scholar] [CrossRef]
- Li, S.; Fu, H.; Wang, Y.; Wang, L.; Jia, B.; Bian, Y. Curcumin inhibits CT26 cells metastasis by decreasing heparanase expression. J. Leukoc. Biol. 2020, 108, 1727–1733. [Google Scholar] [CrossRef]
- Deng, X.; Chen, C.; Wu, F.; Qiu, L.; Ke, Q.; Sun, R.; Duan, Q.; Luo, M.; Luo, Z. Curcumin Inhibits the Migration and Invasion of Non-Small-Cell Lung Cancer Cells Through Radiation-Induced Suppression of Epithelial-Mesenchymal Transition and Soluble E-Cadherin Expression. Technol. Cancer Res. Treat. 2020, 19, 1533033820947485. [Google Scholar] [CrossRef]
- Chen, P.; Huang, H.-P.; Wang, Y.; Jin, J.; Long, W.-G.; Chen, K.; Zhao, X.-H.; Chen, C.-G.; Li, J. Curcumin overcome primary gefitinib resistance in non-small-cell lung cancer cells through inducing autophagy-related cell death. J. Exp. Clin. Cancer Res. 2019, 38, 254. [Google Scholar] [CrossRef]
- Goel, A.; Aggarwal, B.B. Curcumin, the golden spice from Indian saffron, is a chemosensitizer and radiosensitizer for tumors and chemoprotector and radioprotector for normal organs. Nutr. Cancer 2010, 62, 919–930. [Google Scholar] [CrossRef]
- Yoshida, K.; Toden, S.; Ravindranathan, P.; Han, H.; Goel, A. Curcumin sensitizes pancreatic cancer cells to gemcitabine by attenuating PRC2 subunit EZH2, and the lncRNA PVT1 expression. Carcinogenesis 2017, 38, 1036–1046. [Google Scholar] [CrossRef]
- Shaikh, S.; Shaikh, J.; Naba, Y.S.; Doke, K.; Ahmed, K.; Yusufi, M. Curcumin: Reclaiming the lost ground against cancer resistance. Cancer Drug Resist. 2021, 4, 298–320. [Google Scholar] [CrossRef]
- Xiao, J.; Sheng, X.; Zhang, X.; Guo, M.; Ji, X. Curcumin protects against myocardial infarction-induced cardiac fibrosis via SIRT1 activation in vivo and in vitro. Drug Des. Devel Ther. 2016, 10, 1267–1277. [Google Scholar] [CrossRef]
- Kars, M.D.; Iseri, O.D.; Gündüz, U.; Ural, A.U.; Arpaci, F.; Molnár, J. Development of rational in vitro models for drug resistance in breast cancer and modulation of MDR by selected compounds. Anticancer Res. 2006, 26, 4559–4568. [Google Scholar]
- Al-Oqail, M.M.; Farshori, N.N.; Al-Sheddi, E.S.; Musarrat, J.; Al-Khedhairy, A.A.; Siddiqui, M.A. In vitro cytotoxic activity of seed oil of fenugreek against various cancer cell lines. Asian Pac. J. Cancer Prev. 2013, 14, 1829–1832. [Google Scholar] [CrossRef]
- Al-Oqail, M.M.; Al-Sheddi, E.S.; Siddiqui, M.A.; Musarrat, J.; Al-Khedhairy, A.A.; Farshori, N.N. Anticancer activity of chloroform extract and sub-fractions of nepeta deflersiana on human breast and lung cancer cells: An in vitro cytotoxicity assessment. Pharmacogn. Mag. 2015, 11 (Suppl. 4), S598–S605. [Google Scholar] [CrossRef]
- Al-Sheddi, E.S.; Farshori, N.N.; Al-Oqail, M.M.; Musarrat, J.; Al-Khedhairy, A.A.; Siddiqui, M.A. Cytotoxicity of Nigella sativa seed oil and extract against human lung cancer cell line. Asian Pac. J. Cancer Prev. 2014, 15, 983–987. [Google Scholar] [CrossRef]
- Farshori, N.N.; Al-Sheddi, E.S.; Al-Oqail, M.M.; Musarrat, J.; Al-Khedhairy, A.A.; Siddiqui, M.A. Anticancer activity of Petroselinum sativum seed extracts on MCF-7 human breast cancer cells. Asian Pac. J. Cancer Prev. 2013, 14, 5719–5723. [Google Scholar] [CrossRef]
- Farshori, N.N.; Al-Sheddi, E.S.; Al-Oqail, M.M.; Musarrat, J.; Al-Khedhairy, A.A.; Siddiqui, M.A. Cytotoxicity assessments of Portulaca oleracea and Petroselinum sativum seed extracts on human hepatocellular carcinoma cells (HepG2). Asian Pac. J. Cancer Prev. 2014, 15, 6633–6638. [Google Scholar] [CrossRef]
- Zhou, Q.; Ye, M.; Lu, Y.; Zhang, H.; Chen, Q.; Huang, S.; Su, S. Curcumin improves the tumoricidal effect of mitomycin C by suppressing ABCG2 expression in stem cell-like breast cancer cells. PLoS ONE 2015, 10, e0136694. [Google Scholar]
- Quispe-Soto, E.T.; Calaf, G.M. Effect of curcumin and paclitaxel on breast carcinogenesis. Int. J. Oncol. 2016, 49, 2569–2577. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, J.; Zhou, J.; Huang, Z.; Hu, H.; Qiao, M.; Zhao, X.; Chen, D. Synergistic effect of cucurbitacin B in combination with curcumin via enhancing apoptosis induction and reversing multidrug resistance in human hepatoma cells. Eur. J. Pharmacol. 2015, 768, 28–40. [Google Scholar] [CrossRef]
- Khan, M.A.; Zafaryab, M.; Mehdi, S.H.; Ahmad, I.; Rizvi, M.M. Characterization and anti-proliferative activity of curcumin loaded chitosan nanoparticles in cervical cancer. Int. J. Biol. Macromol. 2016, 93, 242–253. [Google Scholar] [CrossRef]
- Lee, H.M.; Patel, V.; Shyur, L.F.; Lee, W.L. Copper supplementation amplifi es the anti-tumor effect of curcumin in oral cancer cells. Phytomedicine 2016, 23, 1535–1544. [Google Scholar] [CrossRef]
- He, G.; Feng, C.; Vinothkumar, R.; Chen, W.; Dai, X.; Chen, X.; Ye, Q.; Qiu, C.; Zhou, H.; Wang, Y.; et al. Curcumin analog EF24 induces apoptosis via ROS-dependent mitochondrial dysfunction in human colorectal cancer cells. Cancer Chemother Pharm. 2016, 78, 1151–1161. [Google Scholar] [CrossRef]
- Montgomery, A.; Adeyeni, T.; San, K.; Heuertz, R.M.; Ezekiel, U.R. Curcumin sensitizes silymarin to exert synergistic anticancer activity in colon cancer cells. J. Cancer 2016, 7, 1250–1257. [Google Scholar] [CrossRef]
- Shakibaei, M.; Buhrmann, C.; Kraehe, P.; Shayan, P.; Lueders, C.; Goel, A. Curcumin chemosensitizes 5-fl uorouracil resistant MMR-deficient human colon cancer cells in high density cultures. PLoS ONE 2014, 9, e85397. [Google Scholar] [CrossRef]
- George, P.A.B.; Tynga, I.M.; Abrahamse, H. In vitro antiproliferative effect of the acetone extract of Rubus fairholmianus gard. BioMed Res. Int. 2015, 2015, 165037. [Google Scholar] [CrossRef]
- Saad, B.; Dakwar, S.; Said, O.; Abu-Hijleh, G.; Al Battah, F.; Kmeel, A.; Aziazeh, H. Evaluation of medicinal plant hepatotoxicity in co-cultures of hepatocytes and monocytes. Evid. Based Complement. Alternat. Med. 2006, 3, 93–98. [Google Scholar]
- Anto, R.J.; Mukhopadhyay, A.; Denning, K.; Aggarwal, B.B. Curcumin (diferuloylmethane) induces apoptosis through activation of caspase-8, BID cleavage and cytochrome C release: Its suppression by ectopic expression of Bcl-2 and Bcl-XL. Carcinogenesis 2002, 23, 143–150. [Google Scholar] [CrossRef]
- Yallapu, M.M.; Jaggi, M.; Chauhan, S.C. Curcumin nanomedicine: A road to cancer therapeutics. Curr. Pharm. Des. 2013, 19, 1994–2010. [Google Scholar]
- Simon, A.; Allais, D.; Duroux, J.; Basly, J.; Durand-Fontanier, S.; Delage, C. Inhibitory effect of curcuminoids on MCF-7 cell proliferation and structure–activity relationships. Cancer Lett. 1998, 129, 111–116. [Google Scholar] [CrossRef]
- Hashemi, M. The study of pentoxifylline drug effects on renal apoptosis and BCL-2 gene expression changes following ischemic reperfusion injury in rat. Iran. J. Pharm. Res. 2014, 13, 181. [Google Scholar] [PubMed]
- Kizhakkayil, J.; Thayyullathil, F.; Chathoth, S.; Hago, A.; Patel, M.; Galadari, S. Modulation of curcumin-induced Akt phosphorylation and apoptosis by PI3K inhibitor in MCF-7 cells. Biochem. Biophys. Res. Commun. 2010, 394, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Sureshbabul, M.; Chen, H.-W.; Lin, Y.-S.; Lee, J.-Y.; Hong, Q.-S.; Yang, Y.-C.; Yu, S.-L. Curcumin Suppresses Metastasis via Sp-1, FAK Inhibition, and E-Cadherin Upregulation in Colorectal Cancer. Evid. Based Complement. Altern. Med. 2013, 2013, 541695. [Google Scholar] [CrossRef]
- Notarbartolo, M.; Poma, P.; Perri, D.; Dusonchet, L.; Cervello, M.; D’Alessandro, N. Antitumor effects of curcumin, alone or in combination with cisplatin or doxorubicin, on human hepatic cancer cells. Analysis of their possible relationship to changes in NF-κB activation levels and in IAP gene expression. Cancer Lett. 2005, 224, 53–65. [Google Scholar] [CrossRef]
- Kuttan, R.; Bhanumathy, P.; Nirmala, K.; George, M.C. Potential anticancer activity of turmeric (Curcuma longa). Cancer Lett. 1985, 29, 197–202. [Google Scholar] [CrossRef]
- Anuchapreeda, S.; Limtrakul, P.; Thanarattanakorn, P.; Sittipreechacharn, S.; Chanarat, P. Inhibitory effect of curcumin on WT1 gene expression in patient leukemic cells. Arch. Pharm. Res. 2006, 29, 80–87. [Google Scholar] [CrossRef]
- Gu, Y.; Zhang, Z.; Yin, J.; Ye, J.; Song, Y.; Liu, H.; Xiong, Y.; Lu, M.; Zheng, G.; He, Z. Epigenetic silencing of miR-493 increases the resistance to cisplatin in lung cancer by targeting tongue cancer resistance-related protein 1 (TCRP1). J. Exp. Clin. Cancer Res. 2017, 36, 1–13. [Google Scholar] [CrossRef]
- Koch, J.; Hau, J.; Jensen, H.E.; Rieneck, K. Cancer resistance as an acquired and inheritable trait. Anticancer Res. 2014, 34, 6315–6325. [Google Scholar]
- Marin, J.J.; Briz, O.; Rodríguez-Macias, G.; Díez-Martín, J.L.; Macias, R.I. Role of drug transport and metabolism in the chemoresistance of acute myeloid leukemia. Blood Rev. 2016, 30, 55–64. [Google Scholar] [CrossRef]
- Kartal-Yandim, M.; Adan-Gokbulut, A.; Baran, Y. Molecular mechanisms of drug resistance and its reversal in cancer. Crit. Rev. Biotechnol. 2016, 36, 716–726. [Google Scholar] [CrossRef]
- Wieringa, H.W.; van der Zee, A.G.; de Vries, E.G.; van Vugt, M.A. Breaking the DNA damage response to improve cervical cancer treatment. Cancer Treat. Rev. 2016, 42, 30–40. [Google Scholar] [CrossRef]
- Mohammad, R.M.; Muqbil, I.; Lowe, L.; Yedjou, C.; Hsu, H.-Y.; Lin, L.-T.; Siegelin, M.D.; Fimognari, C.; Kumar, N.B.; Dou, Q.P. Broad Targeting of Resistance to Apoptosis in Cancer, Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2015; pp. S78–S103. [Google Scholar]
- Nantajit, D.; Lin, D.; Li, J.J. The network of epithelial–mesenchymal transition: Potential new targets for tumor resistance. J. Cancer Res. Clin. Oncol. 2015, 141, 1697–1713. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Chen, X.; Song, M.; Zhang, B.; Zhang, Y. Reactive Oxygen Species Regulate T Cell Immune Response in the Tumor Microenvironment. Oxid. Med. Cell Longev. 2016, 2016, 1580967. [Google Scholar] [CrossRef]
- Yokoyama, C.; Sueyoshi, Y.; Ema, M.; Mori, Y.; Takaishi, K.; Hisatomi, H. Induction of oxidative stress by anticancer drugs in the presence and absence of cells. Oncol. Lett. 2017, 14, 6066–6070. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, Z. Increased oxidative stress as a selective anticancer therapy. Oxidative Med. Cell. Longev. 2015, 2015, 294303. [Google Scholar] [CrossRef]
- Ahsan, H.; Parveen, N.; Khan, N.U.; Hadi, S. Pro-oxidant, anti-oxidant and cleavage activities on DNA of curcumin and its derivatives demethoxycurcumin and bisdemethoxycurcumin. Chem. Biol. Interact. 1999, 121, 161–175. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A review of its effects on human health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Khan, M.A.; Gahlot, S.; Majumdar, S. Oxidative stress induced by curcumin promotes the death of cutaneous T-cell lymphoma (HuT-78) by disrupting the function of several molecular targets. Mol. Cancer Ther. 2012, 11, 1873–1883. [Google Scholar] [CrossRef]
- Swatson, W.S.; Katoh-Kurasawa, M.; Shaulsky, G.; Alexander, S. Curcumin affects gene expression and reactive oxygen species via a PKA dependent mechanism in Dictyostelium discoideum. PLoS ONE 2017, 12, e0187562. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Duan, X.; Cai, H.; Wang, L.; Li, M.; Qu, J.; Li, W.; Wang, Y.; Wang, J. Curcumin inhibits the invasion of lung cancer cells by modulating the PKCα/Nox-2/ROS/ATF-2/MMP-9 signaling pathway. Oncol. Rep. 2015, 34, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Pongrakhananon, V.; Nimmannit, U.; Luanpitpong, S.; Rojanasakul, Y.; Chanvorachote, P. Curcumin sensitizes non-small cell lung cancer cell anoikis through reactive oxygen species-mediated Bcl-2 downregulation. Apoptosis 2010, 15, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Palanikumar, L.; Ragunathan, I.; Panneerselvam, N. Chromosome aberrations induced by curcumin and aloin in Allium cepa L. root meristem cells. Turk. J. Biol. 2011, 35, 145–152. [Google Scholar] [CrossRef]
- Bhaumik, S.; Anjum, R.; Rangaraj, N.; Pardhasaradhi, B.; Khar, A. Curcumin mediated apoptosis in AK-5 tumor cells involves the production of reactive oxygen intermediates. FEBS Lett. 1999, 456, 311–314. [Google Scholar] [CrossRef]
- Wilken, R.; Veena, M.S.; Wang, M.B.; Srivatsan, E.S. Curcumin: A review of anti-cancer properties and therapeutic activity in head and neck squamous cell carcinoma. Mol. Cancer 2011, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.L.; Lin, J.K.; Hsu, M.M.; Ho, Y.F.; Shen, T.S.; Ko, Y.K.; Lin, J.T.; Lin, B.R.; Ming-Shiang, W.; Yu, H.S.; et al. Phase I clinical trial of curcumin, a chemo preventive agent, in patients with high-risk or pre-malignant le-sions. Anticancer Res. 2001, 21, 2895–2900. [Google Scholar]
- Wu, W.-S.; Wu, J.-R.; Hu, C.-T. Signal cross talks for sustained MAPK activation and cell migration: The potential role of reactive oxygen species. Cancer Metastasis Rev. 2008, 27, 303–314. [Google Scholar] [CrossRef]
- Das, L.; Vinayak, M. Long term effect of curcumin in regulation of glycolytic pathway and angiogenesis via modulation of stress activated genes in prevention of cancer. PLoS ONE 2014, 9, e99583. [Google Scholar] [CrossRef]
- Lee, J.C.; Kinniry, P.A.; Arguiri, E.; Serota, M.; Kanterakis, S.; Chatterjee, S.; Solomides, C.C.; Javvadi, P.; Koumenis, C.; Cengel, K.A. Dietary curcumin increases antioxidant defenses in lung, ameliorates radiation-induced pulmonary fibrosis, and improves survival in mice. Radiat. Res. 2010, 173, 590–601. [Google Scholar] [CrossRef]
- Chen, Q.-Y.; Zheng, Y.; Jiao, D.-M.; Chen, F.-Y.; Hu, H.-Z.; Wu, Y.-Q.; Song, J.; Yan, J.; Wu, L.-J.; Lv, G.-Y. Curcumin inhibits lung cancer cell migration and invasion through Rac1-dependent signaling pathway. J. Nutr. Biochem. 2014, 25, 177–185. [Google Scholar] [CrossRef]
- Chen, W.-C.; Lai, Y.-A.; Lin, Y.-C.; Ma, J.-W.; Huang, L.-F.; Yang, N.-S.; Ho, C.-T.; Kuo, S.-C.; Way, T.-D. Curcumin suppresses doxorubicin-induced epithelial–mesenchymal transition via the inhibition of TGF-β and PI3K/AKT signaling pathways in triple-negative breast Cancer cells. J. Agric. Food Chem. 2013, 61, 11817–11824. [Google Scholar] [CrossRef]
- Wardhani, B.W.; Puteri, M.U.; Watanabe, Y.; Louisa, M.; Setiabudy, R.; Kato, M. TGF-β-induced TMEPAI attenuates the response of triple-negative breast cancer cells to doxorubicin and paclitaxel. J. Exp. Pharmacol. 2020, 12, 17. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhou, L.; Zhang, T.; Qin, C.; Wei, P.; Luo, L.; Luo, L.; Huang, G.; Chen, A.; Liu, G. Anti-fibrosis activity of quercetin attenuates rabbit tracheal stenosis via the TGF-β/AKT/mTOR signaling pathway. Life Sci. 2020, 250, 117552. [Google Scholar] [CrossRef]
- Liu, R.; Wang, Q.; Ding, Z.; Zhang, X.; Li, Y.; Zang, Y.; Zhang, G. Silibinin augments the antifibrotic effect of valsartan through inactivation of TGF-β1 signaling in kidney. Drug Des. Dev. Ther. 2020, 14, 603. [Google Scholar] [CrossRef]
- Fang, L.; Li, Y.; Wang, S.; Li, Y.; Chang, H.-M.; Yi, Y.; Yan, Y.; Thakur, A.; Leung, P.C.; Cheng, J.-C. TGF-β1 induces VEGF expression in human granulosa-lutein cells: A potential mechanism for the pathogenesis of ovarian hyperstimulation syndrome. Exp. Mol. Med. 2020, 52, 450–460. [Google Scholar] [CrossRef]
- Qin, F.; Liu, X.; Chen, J.; Huang, S.; Wei, W.; Zou, Y.; Liu, X.; Deng, K.; Mo, S.; Chen, J. Anti-TGF-β attenuates tumor growth via polarization of tumor associated neutrophils towards an anti-tumor phenotype in colorectal cancer. J. Cancer 2020, 11, 2580. [Google Scholar] [CrossRef]
- Celik, H.; Aydin, T.; Solak, K.; Khalid, S.; Farooqi, A.A. Curcumin on the “flying carpets” to modulate different signal transduction cascades in cancers: Next-generation approach to bridge translational gaps. J. Cell. Biochem. 2018, 119, 4293–4303. [Google Scholar] [CrossRef]
- Mo, N.; Li, Z.-Q.; Li, J.; Cao, Y.-D. Curcumin inhibits TGF-β1-induced MMP-9 and invasion through ERK and Smad signaling in breast cancer MDA-MB-231 cells. Asian Pac. J. Cancer Prev. 2012, 13, 5709–5714. [Google Scholar] [CrossRef]
- Malik, M.; Mendoza, M.; Payson, M.; Catherino, W.H. Curcumin, a nutritional supplement with antineoplastic activity, enhances leiomyoma cell apoptosis and decreases fibronectin expression. Fertil. Steril. 2009, 91, 2177–2184. [Google Scholar] [CrossRef]
- Hu, C.; Li, M.; Guo, T.; Wang, S.; Huang, W.; Yang, K.; Liao, Z.; Wang, J.; Zhang, F.; Wang, H. Anti-metastasis activity of curcumin against breast cancer via the inhibition of stem cell-like properties and EMT. Phytomedicine 2019, 58, 152740. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-P.; Zhai, P.; Yamamoto, T.; Maejima, Y.; Matsushima, S.; Hariharan, N.; Shao, D.; Takagi, H.; Oka, S.; Sadoshima, J. Silent information regulator 1 protects the heart from ischemia/reperfusion. Circulation 2010, 122, 2170–2182. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, X.; Zhou, Q.; Huang, C.; Meng, X.; Xu, F.; Li, J. Silent information regulator 1 (SIRT1) ameliorates liver fibrosis via promoting activated stellate cell apoptosis and reversion. Toxicol. Appl. Pharmacol. 2015, 289, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Rizk, S.M.; El-Maraghy, S.A.; Nassar, N.N. A novel role for SIRT-1 in L-arginine protection against STZ induced myocardial fibrosis in rats. PLoS ONE 2014, 9, e114560. [Google Scholar]
- Yang, Y.; Duan, W.; Lin, Y.; Yi, W.; Liang, Z.; Yan, J.; Wang, N.; Deng, C.; Zhang, S.; Li, Y. SIRT1 activation by curcumin pretreatment attenuates mitochondrial oxidative damage induced by myocardial ischemia reperfusion injury. Free Radic. Biol. Med. 2013, 65, 667–679. [Google Scholar] [CrossRef]
- Sun, Q.; Jia, N.; Wang, W.; Jin, H.; Xu, J.; Hu, H. Activation of SIRT1 by curcumin blocks the neurotoxicity of amyloid-β25–35 in rat cortical neurons. Biochem. Biophys. Res. Commun. 2014, 448, 89–94. [Google Scholar] [CrossRef]
- Hou, M.; Zuo, X.; Li, C.; Zhang, Y.; Teng, Y. Mir-29b regulates oxidative stress by targeting SIRT1 in ovarian cancer cells. Cell. Physiol. Biochem. 2017, 43, 1767–1776. [Google Scholar] [CrossRef]
- Hacioglu, C. Capsaicin inhibits cell proliferation by enhancing oxidative stress and apoptosis through SIRT1/NOX4 signaling pathways in HepG2 and HL-7702 cells. J. Biochem. Mol. Toxicol. 2022, 36, e22974. [Google Scholar] [CrossRef]
- Hung, L.V.M.; Song, Y.W.; Cho, S.K. Effects of the Combination of Gliotoxin and Adriamycin on the Adriamycin-Resistant Non-Small-Cell Lung Cancer A549 Cell Line. Mar. Drugs 2018, 16, 105. [Google Scholar] [CrossRef]
- Lv, Z.D.; Liu, X.P.; Zhao, W.J.; Dong, Q.; Li, F.N.; Wang, H.B.; Kong, B. Curcumin induces apoptosis in breast cancer cells and inhibits tumor growth in vitro and in vivo. Int. J. Clin. Exp. Pathol. 2014, 7, 2818–2824. [Google Scholar]
- Paramita, P.; Wardhani, B.W.K.; Wanandi, S.I.; Louisa, M. Curcumin for the prevention of epithelial-mesenchymal transition in endoxifen-treated MCF-7 breast cancer cells. Asian Pac. J. Cancer Prev. 2018, 19, 1243. [Google Scholar]
- Freshney, R.I. Basic Principles of Cell Culture. Culture Cells for Tissue Enginnerging; Freshney, R.I., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006; pp. 1–22. [Google Scholar]
- Shamim, S.; Rehman, A. Antioxidative enzyme profiling and biosorption ability of Cupriavidus metallidurans CH34 and Pseudomonas putida mt2 under cadmium stress. J. Basic Microbiol. 2015, 55, 374–381. [Google Scholar] [CrossRef]
- Shaikh, S.B.; Najar, M.A.; Prabhu, A.; Rex, D.A.B.; Chanderasekaran, J.; Behera, S.K.; Modi, P.K.; Prasad, T.S.K.; Bhandary, Y.P. The unique molecular targets associated antioxidant and antifibrotic activity of curcumin in vitro model of acute lung injury: A proteomic approach. Biofactors 2021, 47, 627–644. [Google Scholar] [CrossRef]
- Bergman, I.; Loxley, R. Two improved and simplified methods for the spectrophotometric determination of hydroxyproline. Anal. Chem. 1963, 35, 1961–1965. [Google Scholar] [CrossRef]
- Attallah, A.M.; Abdallah, S.O.; Attallah, A.A.; Omran, M.M.; Farid, K.; Nasif, W.A.; Shiha, G.E.; Abdel-Aziz, A.-A.F.; Rasafy, N.; Shaker, Y.M. Diagnostic value of fibronectin discriminant score for predicting liver fibrosis stages in chronic hepatitis C virus patients. Ann. Hepatol. 2013, 12, 44–53. [Google Scholar] [CrossRef]
- Kojima, K.; Ohhashi, R.; Fujita, Y.; Hamada, N.; Akao, Y.; Nozawa, Y.; Deguchi, T.; Ito, M. A role for SIRT1 in cell growth and chemoresistance in prostate cancer PC3 and DU145 cells. Biochem. Biophys. Res. Commun. 2008, 373, 423–428. [Google Scholar] [CrossRef]
- Borhani, N.; Manoochehri, M.; Gargari, S.S.; Novin, M.G.; Mansouri, A.; Omrani, M.D. Decreased expression of proapoptotic genes caspase-8-and BCL2-associated agonist of cell death (BAD) in ovarian cancer. Clin. Ovarian Other Gynecol. Cancer 2014, 7, 18–23. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Curcumin Concentrations (µM) | Cell Inhibition/Cytotoxicity (%) on Cell Lines | |||||
|---|---|---|---|---|---|---|
| Sensitive Cells | Resistant Cells | |||||
| MCF7 | HCT116 | A549 | MCF7/TH | HCT116R | A549/ADR | |
| 2.7 µM | 45.6 a | 44.3 a | 41.5 a | 35.8 a,b | 37.3 a,b | 32.7 a,b |
| 6.8 µM | 55.3 a | 51.9 a | 59.2 a | 46.7 a | 45.8 a | 39.8 a |
| 13.6 µM | 62.5 a | 68.4 a | 67.5 a | 56.3 a | 59.5 a | 53.6 a |
| 27.0 µM | 78.6 a | 82.5 a | 79.4 a | 72.8 a | 76.4 a | 73.3 a |
| 54.3 µM | 92.7 a | 91.5 a | 90.6 a | 85.1 a,b | 87.4 a,b | 86.2 a,b |
| DMSO (0.1% w/v) control | N.I. | N.I. | N.I. | N.I. | N.I. | N.I. |
| Parameters | Control | Treated Cells | ||||
|---|---|---|---|---|---|---|
| SIRT1 Activation | SIRT1 Activation | |||||
| Curcumin (2.7 µM) | Curcumin (54.3 µM) | |||||
| R | p-Value | R | p-Value | R | p-Value | |
| Oxidative stress | ||||||
| Cellular ROS | 0.67 | 0.002 | 0.69 | 0.001 | 0.75 | 0.001 |
| SOD activity | 0.48 | 0.001 | 0.34 | 0.001 | −0.82 | 0.001 |
| CAT activity | 0.28 | 0.001 | 0.72 | 0.001 | −0.69 | 0.001 |
| Fibrosis markers | ||||||
| TGF-β1 | −0.52 | 0.001 | −0.89 | 0.004 | −0.96 | 0.006 |
| Hydroxyproline | −0.84 | 0.002 | −0.72 | 0.001 | −0.85 | 0.001 |
| Fibronectin | −0.67 | 0.01 | −0.54 | 0.01 | −0.63 | 0.001 |
| Apoptotic markers | ||||||
| anti-Annexin V | 0.58 | 0.01 | 0.38 | 0.01 | 0.29 | 0.01 |
| Bcl-2 | −0.35 | 0.002 | −0.41 | 0.001 | −0.56 | 0.001 |
| BAX | 0.65 | 0.001 | 0.49 | 0.001 | 0.51 | 0.001 |
| BAX/Bcl-2 ratio | 0.85 | 0.001 | 0.78 | 0.001 | 0.54 | 0.001 |
| Cytochrome c | 0.39 | 0.001 | 0.42 | 0.001 | 0.61 | 0.001 |
| Caspase-3 | 0.58 | 0.002 | 0.38 | 0.002 | 0.45 | 0.003 |
| Primer ID | Primer Sequences |
|---|---|
| SIRT1 gene | |
| Forward sequence | 5′-TGGCAAAGGAGCAGATTAGTAGG-3′ |
| Reverse sequence | 5′-CTGCCACAAGAACTAGAGGATAAGA |
| GAPDH gene | |
| Forward sequence | 5′-AAGCTCATTTCCTGGTATG-3′ |
| Reverse sequence | 5′-CTTCCTCTTGTGCTCTTG-3′ |
| Bcl-2 gene | |
| Forward sequence | 5′-ATCGCCCTGTGGATGACTGAGT-3′ |
| Reverse sequence | 5′-GCCAGGAGAAATCAAACAGAGGC-3′ |
| Bax gene | |
| Forward sequence | 5′-ATG GAC GGG TCC GGG GAG CA-3′ |
| Reverse sequence | 5′-CCC AGT TGA AGT TGC CGT CA-3′ |
| Caspase-8 gene | |
| Forward sequence | 5′-AGAGTCTGTGCCCAAATCAAC-3′ |
| Reverse sequence | 5′-GCTGCTTCTCTCTTTGCTGAA-3′ |
| Cytochrome c gene | |
| Forward sequence: | 5′-AAGGGAGGCAAGCACAAGACTG-3′ |
| Reverse sequence | 5′-CTCCATCAGTGTATCCTCTCCC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabr, S.A.; Elsaed, W.M.; Eladl, M.A.; El-Sherbiny, M.; Ebrahim, H.A.; Asseri, S.M.; Eltahir, Y.A.M.; Elsherbiny, N.; Eldesoqui, M. Curcumin Modulates Oxidative Stress, Fibrosis, and Apoptosis in Drug-Resistant Cancer Cell Lines. Life 2022, 12, 1427. https://doi.org/10.3390/life12091427
Gabr SA, Elsaed WM, Eladl MA, El-Sherbiny M, Ebrahim HA, Asseri SM, Eltahir YAM, Elsherbiny N, Eldesoqui M. Curcumin Modulates Oxidative Stress, Fibrosis, and Apoptosis in Drug-Resistant Cancer Cell Lines. Life. 2022; 12(9):1427. https://doi.org/10.3390/life12091427
Chicago/Turabian StyleGabr, Sami A., Wael M. Elsaed, Mohamed Ahmed Eladl, Mohamed El-Sherbiny, Hasnaa Ali Ebrahim, Saad Mohamed Asseri, Yasir A. M. Eltahir, Nehal Elsherbiny, and Mamdouh Eldesoqui. 2022. "Curcumin Modulates Oxidative Stress, Fibrosis, and Apoptosis in Drug-Resistant Cancer Cell Lines" Life 12, no. 9: 1427. https://doi.org/10.3390/life12091427
APA StyleGabr, S. A., Elsaed, W. M., Eladl, M. A., El-Sherbiny, M., Ebrahim, H. A., Asseri, S. M., Eltahir, Y. A. M., Elsherbiny, N., & Eldesoqui, M. (2022). Curcumin Modulates Oxidative Stress, Fibrosis, and Apoptosis in Drug-Resistant Cancer Cell Lines. Life, 12(9), 1427. https://doi.org/10.3390/life12091427