1. Introduction
Solidago, often known as golden rod, is a member of the
Asteraceae family. There are roughly 130 species in this genus, most of which are found in North America [
1].
Solidago virgaurea,
S. canadensis, and
S. memoralis are species grown in beds, borders, and rock gardens. They are widely utilized as cut flowers for bouquets and indoor decorating [
2,
3]. One of the most important species in the Solidago genus is the European goldenrod (
Solidago virgaurea L.) plant. It is grown as a perennial flowering plant that is prized for its tall, straight flower stalks [
3,
4]. The aerial parts of Solidago have been used in traditional medicine for millennia as anti-inflammatory, spasmolytic, and diuretic remedies for a variety of ailments, particularly as a urological agent in kidney and bladder inflammation, urolithiasis, and cystitis, and a yellow dye was produced from the flowers [
4,
5,
6]. Furthermore, it is a source of several significant secondary metabolites, all of which have great therapeutic potential. Moreover, Solidago has been shown to have anticancer activity on human prostate (PC-3), breast (MDA435), melanoma (C8161), and small cell lung carcinomas (H520) [
6]. According to the European Medicines Agency,
S. virgaurea is one of the most investigated and used species of the Solidago genus in Europe [
3]. Flavonoids (mainly quercetin glycosides); salicylic acid derivatives (leiocarposide, virgaureoside); caffeoylquinic acid derivatives (chlorogenic acid, caffeic acid); triterpene saponins (oleanane type); tannins; and essential oils are all found in
S. virgaurea extracts [
7]. Several bioactive chemicals found in
S. virgaurea extracts work together: flavonoids, saponins, caffeic acid derivatives, and leiocarposide have anti-inflammatory properties, polyphenolic compounds have antioxidant properties, and flavonoids have spasmolytic properties [
8,
9,
10]. Soil plays a crucial part in defining an agro-long-term ecosystem’s productivity. The ability of soil to provide critical nutrients to growing plants determines its sustainable productivity. Micronutrient insufficiency has become a key barrier to soil productivity, stability, and sustainability [
9]. Soil alkalinity is one of the most common concerns in arid and semi-arid areas, with pH levels ranging from 7.5 to 8.7 [
11,
12,
13]. Arid land makes up 97% of Egypt’s total land area, and it is characterized by high temperatures, low relative humidity, rapid evaporation, and minimal rainfall, resulting in degraded soils [
12]. Soil alkalinization induced by NaHCO
3 and Na
2CO
3 may be more serious than soil salinization caused by neutral salts such as NaCl and Na
2SO
4 [
14,
15,
16]. Egypt’s soils are characterized by pH values that range from slightly alkaline to alkaline, owing to the country’s arid climate. As soil salinization and alkalinization frequently co-occur and are quite complicated, total salt concentration, composition, and the proportion of neutral to alkaline salts can all differ significantly between soils [
15]. Furthermore, alkali stress is characterized by a mixture of stresses, osmotic, ion-induced damage, and a high pH level [
12]. Alkalinity interferes with the uptake of nutrients that contribute to growth and the accumulation of bioactive components. Foliar nutrition is one of the most effective agricultural practices for health growth. Foliar application can also provide 85% of the plant nutritional requirements [
16]. Otherwise, there has recently been a worldwide movement toward using natural compounds that are both safe and non-polluting to the environment.
In particular, the plant’s capacity to produce primary and secondary metabolites is significantly influenced by soil conditions [
17]. Prior research showed that some chemical features of soil are very unique to certain plant species [
18]. Klimien and his co-worker [
19] found that oregano grown in acidic soil reaction led to a lower content of total phenols and extractives in the raw material compared to an alkaline soil.
Active dry yeast (YE) is a natural and safe biofertilizer that plays an important function in plant growth. It is a rich source of essential nutrients, particularly cytokinins, which operate as a rapidly available growth supplement for plants, resulting in increased yield [
20]. As a result, it aids in cell division and expansion, protein and nucleic acid synthesis, and chlorophyll creation, all of which contribute to improved plant growth [
21]. It is also high in amino acids, peptides, and B-complex vitamins, including B1, B2, B6, and B12, as well as carbohydrates, sugars, and minerals. It contains several amino acids, vitamins, and essential elements such as Na, Ca, Fe, K, P, S, Mg, Zn, and Si [
22]. It also emits CO
2, resulting in an improvement in net photosynthesis [
23]. In marigold plants, using yeast as a foliar fertilizer improved growth and plant nutrition [
24]. In the white lupine, when compared to untreated plants, the varied active yeast extract treatments significantly improved the growth and physiological properties [
25]. Internal variables such as hormonal and nutritional balance regulate plant growth and development. Growth regulators, which are increasingly being utilized to influence the growth and flowering of ornamental and medicinal plants, are responsible for the balanced development of plants [
26]. At optimal concentrations, plant growth regulators are known to coordinate and control several phases of growth and development, including flowering. The exogenous application of plant growth regulators alters the concentrations of naturally existing hormones, which then affects the plant’s growth and development [
27]. Gibberellins (GAs) are found in both flowering and non-flowering plants, and they are widely distributed. GAs, especially gibberellic acid (GA
3), belong to the diterpenoid class of bioactive growth regulators [
28]. GA
3 has the ability to alter the growth pattern of plants by influencing DNA and RNA levels, cell division and expansion, enzyme production, protein, carbohydrate biosynthesis, and photosynthetic pigment biosynthesis [
1]. Different GA
3 concentrations substantially affected all golden rod growth, flower quality, and yield characteristics [
29]. The application of GA
3 at various doses to marigold plants positively influenced many growth parameters, flower quality, and yield attributes [
30].
To date, no research has been conducted on the effects of yeast extract alone or in combination with gibberellic acid on European goldenrod. Therefore, in light of the previous facts, the current investigation was implemented to assess the advantages of YE and/or GA3 amendments and their reflections on morpho-physiological characteristics, nutritional accumulation, and biochemical determinations of Solidago.
3. Results
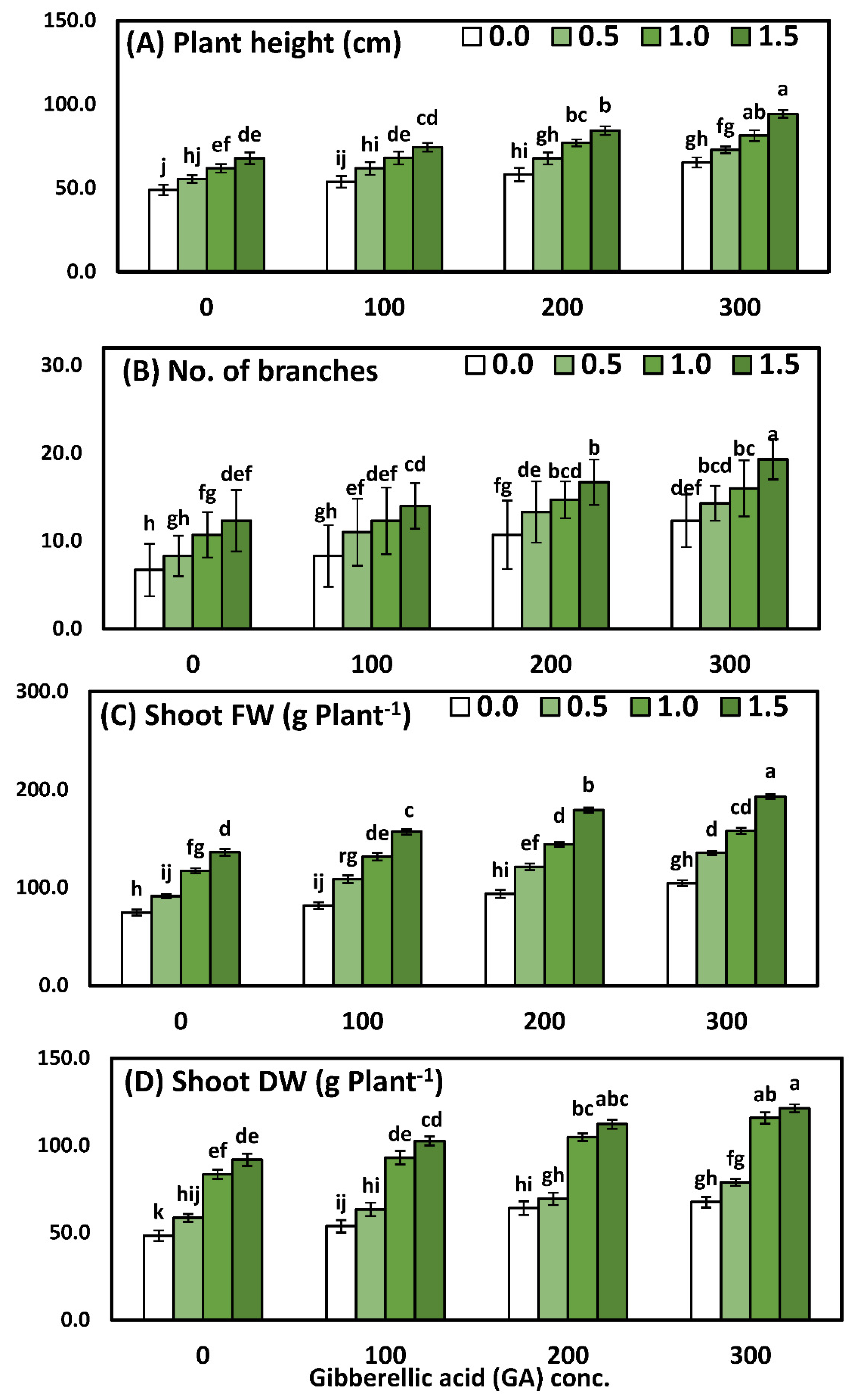
3.1. Effects of YE and GA3 Spraying on the Morphological Measurements of the Solidago Virgaurea Plant, as Well as Their Interactions
Solidago morphological measurements, namely plant height, the number of branches, the fresh and dry weight of shoots responses YE, and the GA
3 concentrations and interaction of foliar application YE with GA3 concentrations, are illustrated in
Table 3 and
Figure 1. Using a two-way analysis of variance, a highly significant difference was seen in the morphological parameters in response to different yeast concentrations (
p < 0.001), GA concentrations (
p < 0.001), and the interaction between YE and GA treatments (
p < 0.001).
Comparisons among the four concentrations of YE indicated that all the growth parameters increased significantly and progressively with every increment of the YE concentrations. Likewise, the application of the highest concentration of YE (1.5 g/L) was pioneering and recorded significantly higher mean values of plant height 80.2 cm, the no. of branches 15.6/plant, shoot fresh weight 166.6 g, and shoot dry weight 107.0 g, compared to the plants that were sprayed with distilled water. With regard to the effect of GA3, the results show that all GA3 concentrations caused significant enhancement for all growth parameters. Furthermore, the highest values of plant height 78.4 cm, the no. of branches 15.5/plant, shoot fresh weight 148.0 g, and shoot dry weight 95.9 g were detected by using GA3 at 300 ppm, in comparison to the control group, which achieved the lowest results. Comparisons among the sixteen mean values of the interaction between the two studied factors had a significant effect on all the growth parameters, as compared to the control or each one alone. Furthermore, the application of YE at 1.5 g/L together with 300 ppm GA3 significantly achieved the highest values, which were plant height 94.3 cm, the no. of branches 19.3/plant, shoot fresh weight 193.3 g, and shoot dry weight 121.3 g.
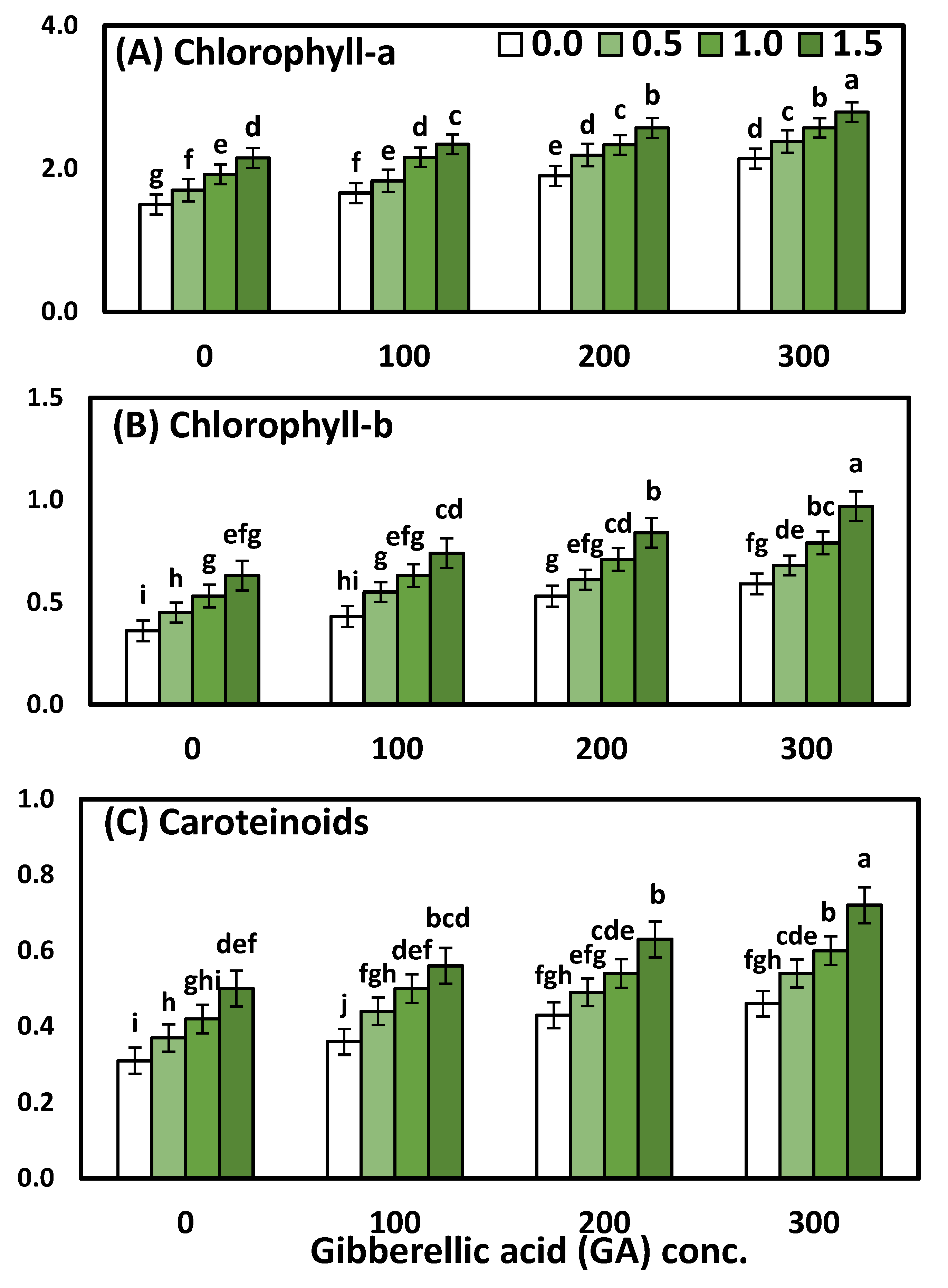
3.2. Effects of YE and GA3 Spraying on the Physiological Measurements of the Solidago Virgaurea Plant, as Well as Their Interactions
Chlorophyll (a), chlorophyll (b), and total carotenoids were affected significantly (
p < 0.001 ***) by YE, the GA
3 concentrations, as well as their interactions, which are presented in
Table 4 and
Figure 2. The two-way analysis of variance revealed a highly significant difference in the physiological parameters (chl-a, chl-b, carotenoids) in response to different YE concentrations (
p < 0.001), GA concentrations (
p < 0.001), and the interaction between the YE and GA treatments (
p < 0.001).
Regarding the effect of YE, the data showed that the general influence of YE spray at different concentrations on all chl-a, chl-ba, and carotenoids was significant. Likewise, the mean values of chlorophyll (a), chlorophyll (b), and total carotenoids were in ascending order, as the YE concentration increased up to the highest concentration. Concerning the GA3 concentrations, it is clear that the highest one (300 ppm) of GA3 excelled the rest of the concentrations, which were markedly increased in all physiological parameters compared to the plants that were sprayed with distilled water and other concentrations of GA3. Furthermore, the increment reached 35.71, 55.10 and 45%, chlorophyll (a) chlorophyll (b), and total carotenoids, respectively. A significant interaction effect between the two studied factors on all the physiological parameters was evident in both seasons. Otherwise, the treatment combination of YE at 1.5 g/L with GA3 at 300 ppm gave the highest significant chlorophyll (a) 2.79 mg mm−2, chlorophyll (b) 0.97 mg mm−2, and total carotenoids 0.72 mg mm−2, as compared to the control and each one alone.
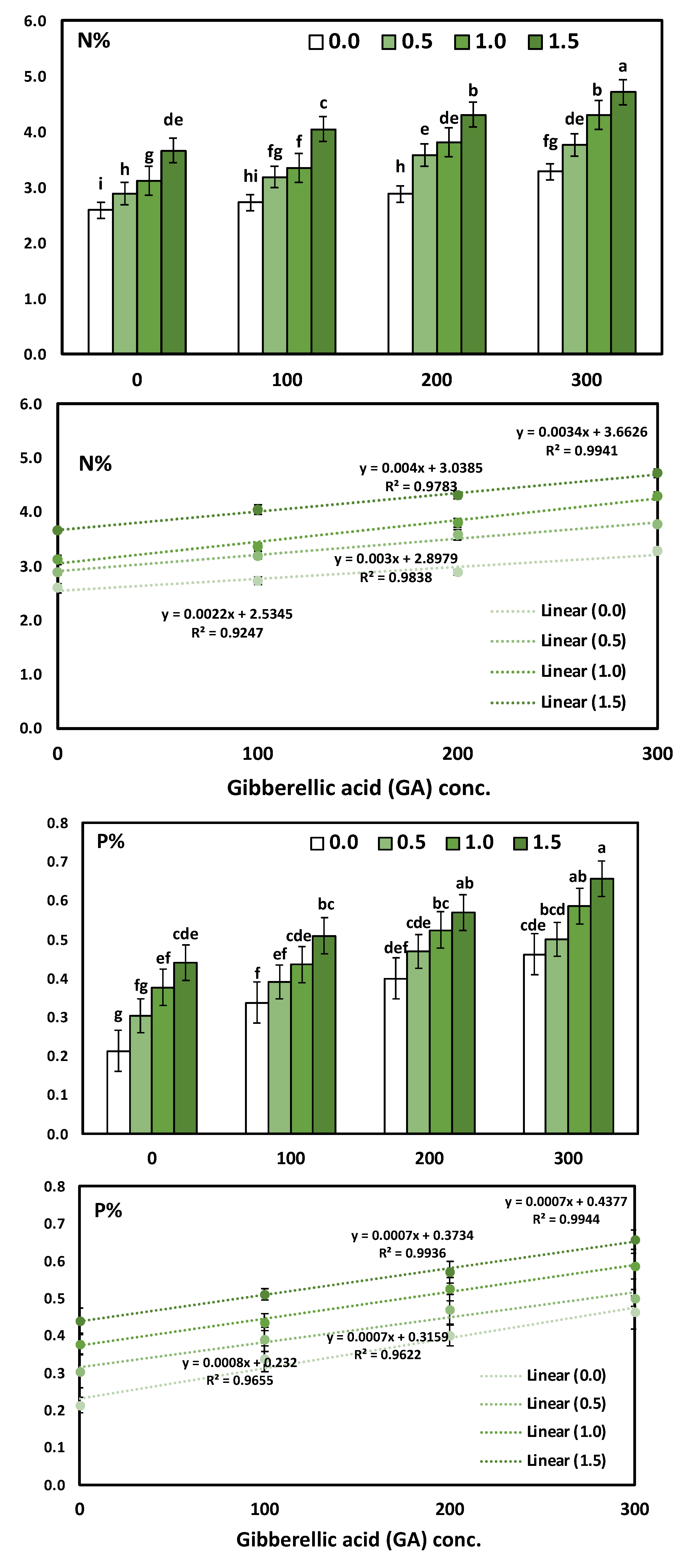
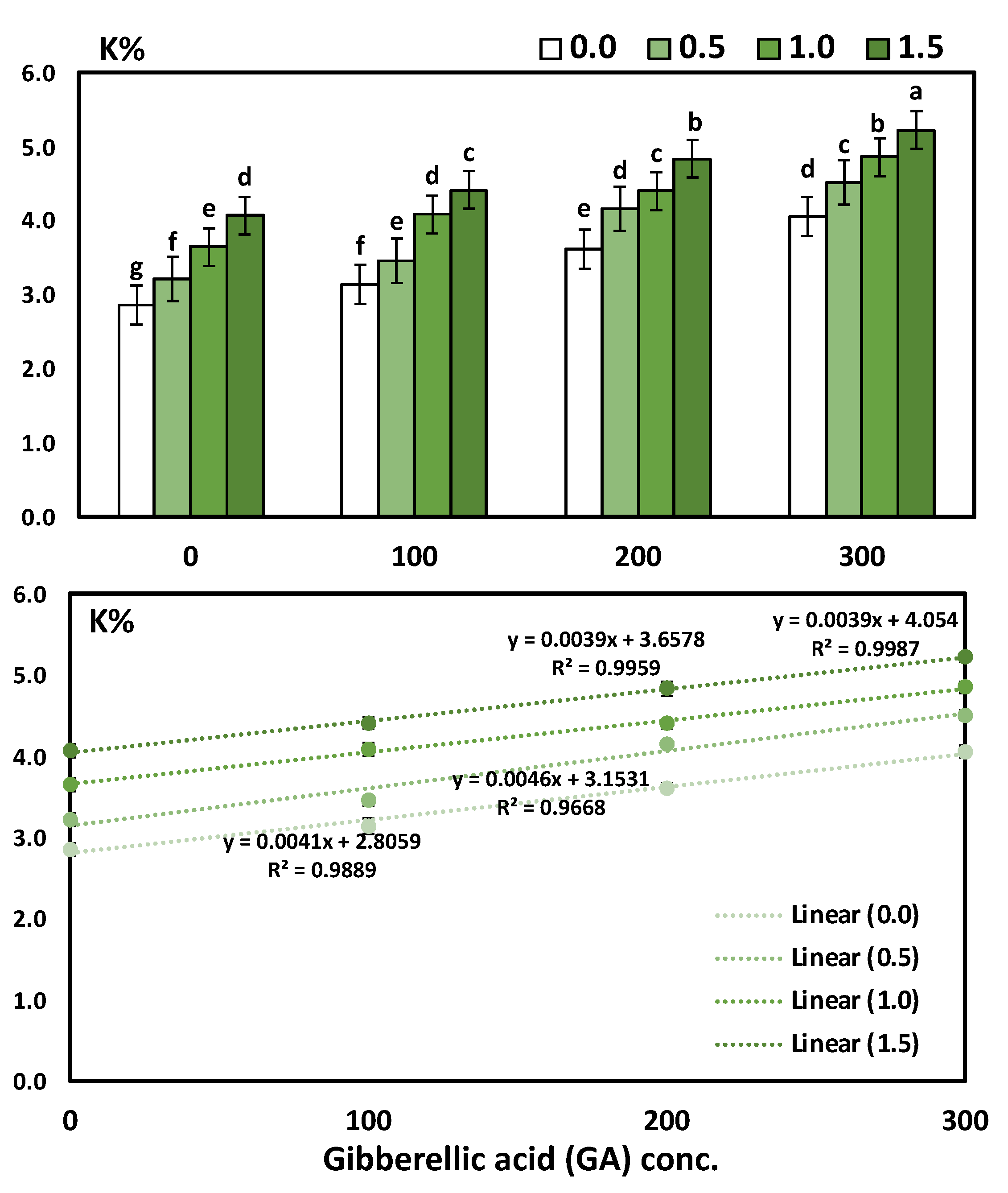
3.3. Effects of YE and GA3 Spraying on the Macronutrient’s Measurements of the Solidago Virgaurea Plant, as Well as Their Interactions
Table 5 and
Figure 3 shows N, P, and K in response to YE, the GA
3 concentrations, and their interactions. The two-way analysis of variance revealed a highly significant difference in macronutrients (N, P, and K) in response to different yeast concentrations (
p < 0.001), GA concentrations (
p < 0.001), and the interaction between the YE and GA treatments (
p < 0.001).
The main effect of YE applied at different concentrations on the accumulation of the content N, P, and K % in the leaves showed a positive correlation. Likewise, the application of YE irrespective to the concentrations used significantly increased the leaves’ N, P and K %, as compared to the control treatment. Moreover, the application of YE at a high concentration (1.5 g/L) showed a highly significant increase in the leaves’ N (4.18%), P (0.54%), and K (4.64%). As for the effect of GA3, the results indicate that all the GA3 concentrations significantly augmented in the N, P, and K % content in the leaves compared to the control treatment. Furthermore, the increases in the mean values of N, P, and K % were linearly correlated with the increase in GA3. The treatment combinations of YE and GA3 concentrations seemed to have a more significant effect on the N, P, and K % in the leaves. Otherwise, the highest results were gained in the content of N, P, and K % in the leaves due to using YE at 1.5 g/L together with GA3 at 300 ppm in the two growing seasons, as compared to the control treatment. The above-mentioned N, P, and K % enhanced by 81.85%, 214.29%, and 82.52%, respectively.
The GA
3 concentration significantly induced a significant positive increase in N, P, and K, as revealed by Pearson’s correlation and simple linear regression. The determination coefficient R
2 ranged from 0.924 to 0.9941, which indicates a strong positive increase in the GA concentrations. The regression trendline was presented in
Figure 3 for N, P, and K with the simple linear regression equation and determination coefficient (R
2).
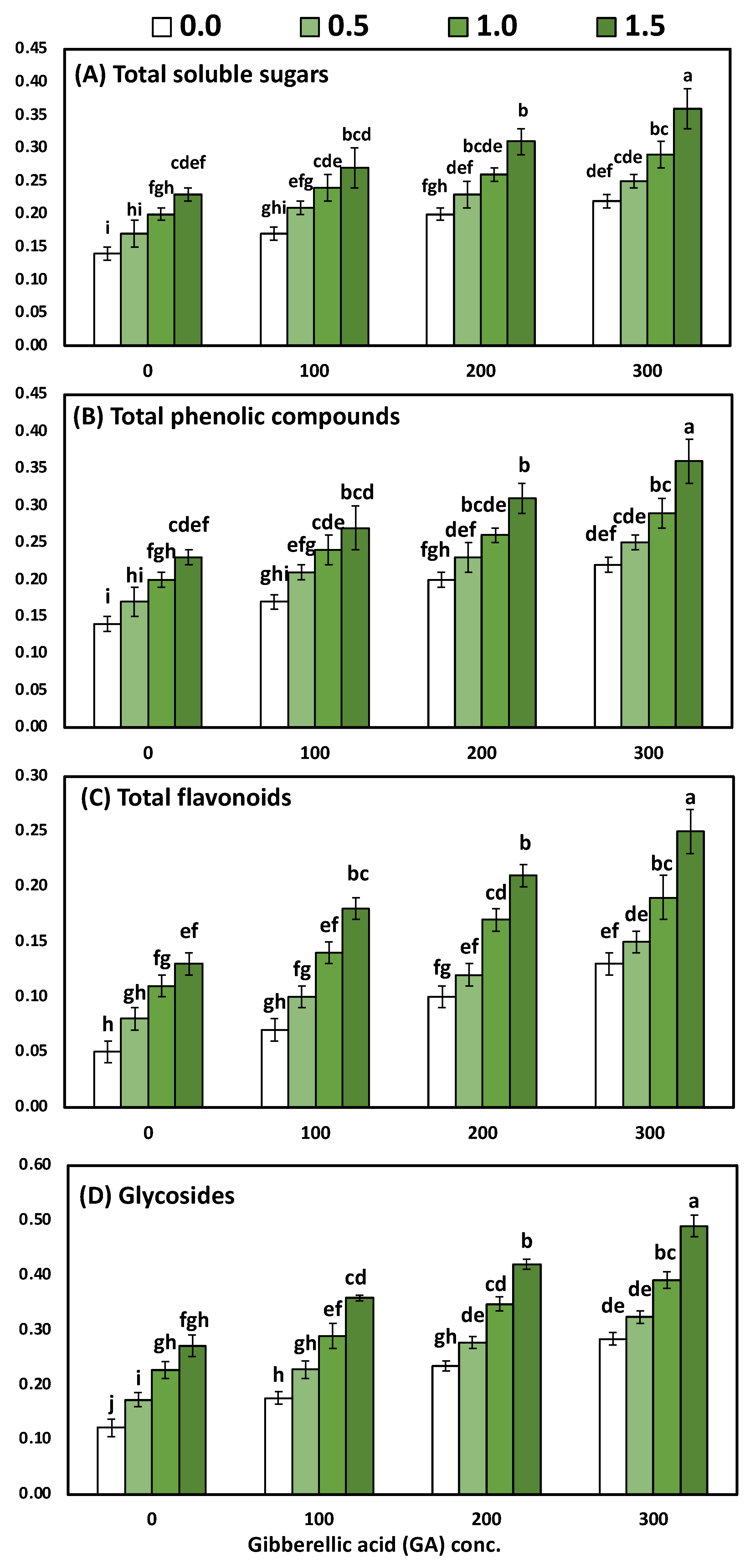
3.4. Effects of YE and GA3 Spraying on the Total Soluble Sugars, Total Phenolic, Total Flavonoids and Total Glycosides Measurements of the Solidago Virgaurea Plant, as Well as Their Interactions
Table 6 and
Figure 4 display the main effects of the two studied factors (YE and GA
3 concentrations) and their interactions on total soluble sugars, total phenolic, total flavonoids and total glycosides in leaves. The two-way analysis of variance revealed a highly significant difference in the biochemical parameters (total soluble sugars, total flavonoids, total phenolic compounds) in response to different yeast concentrations (
p < 0.001), GA concentrations (
p < 0.001), and the interaction between YE and GA treatments (
p < 0.001).
The differences between the four concentrations of YE on the leaves’ total soluble sugars, total phenolic, total flavonoids, and total glycosides were significant. Likewise, progressive increases in all the biochemical constituents occurred due to the foliar application of YE up to the highest concentration, whereas the highest values (0.29, 0.31, 0.20 and 0.38 mg/g DW, total soluble sugars, total phenolic, total flavonoids, and total glycosides, respectively) were detected by 1.5 g/L with YE in both seasons. The detected differences among the mean values of total soluble sugars, total phenolic, total flavonoids, and total glycosides in the leaves within the four utilized concentrations of GA3 were enough to be significant. Likewise, all the biochemical constituents in the leaves were positively correlated with increasing GA3 concentrations, whereas the 300-ppm treatment occupied the first rank in all cases for raising the quantity of all the biochemical constituents in the leaves. The combined influence of the YE and GA3 concentrations had a significant effect on the total soluble sugars, total phenolic, total flavonoids, and total glycosides in the leaves. Furthermore, the highest values of all the biochemical constituents in the leaves were detected by utilizing YE at 1.5 g/L with GA3 at 300 ppm, as compared to the control, which produced the lowest values. The above-mentioned biochemical constituents were increased by 157.14%, 111.76%, 400% and 308.33%, respectively.
A correlation matrix plot showed the interrelationships between the studied variables. The blue colour indicates a positive correlation and red a negative correlation. Boxes indicate a significant correlation (
Figure 5). It shows a positive strong significant correlation between treatments, morphological, physiological, and biochemical parameters.
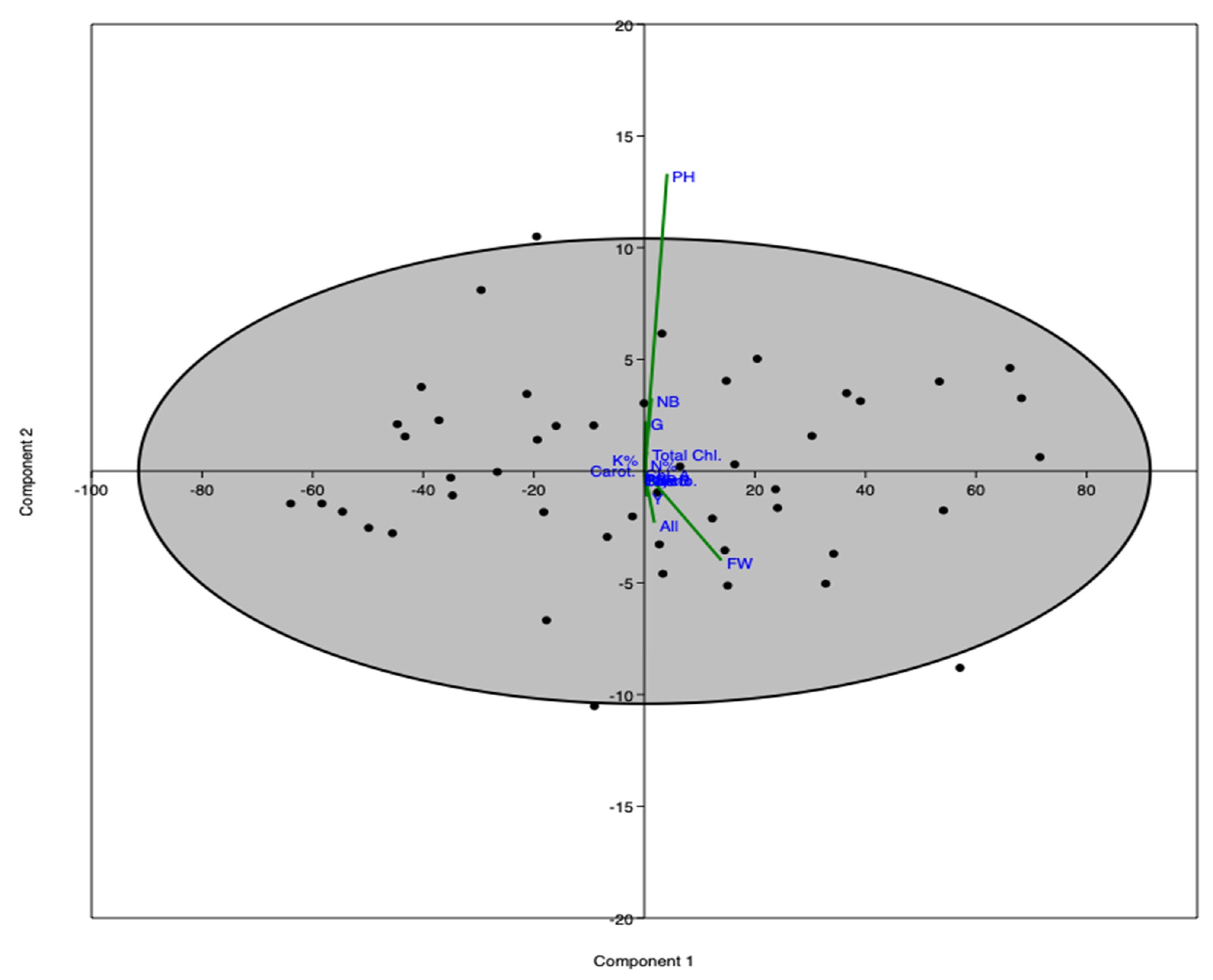
PCA-ordination shows the interrelationship between the variables of the study. PCA-1 and PCA-2 represent more than 99% of the total variance of the study (
Figure 6). The PCA indicates that various morphological and physiological parameters were most affected by the treatments (YE, GA).
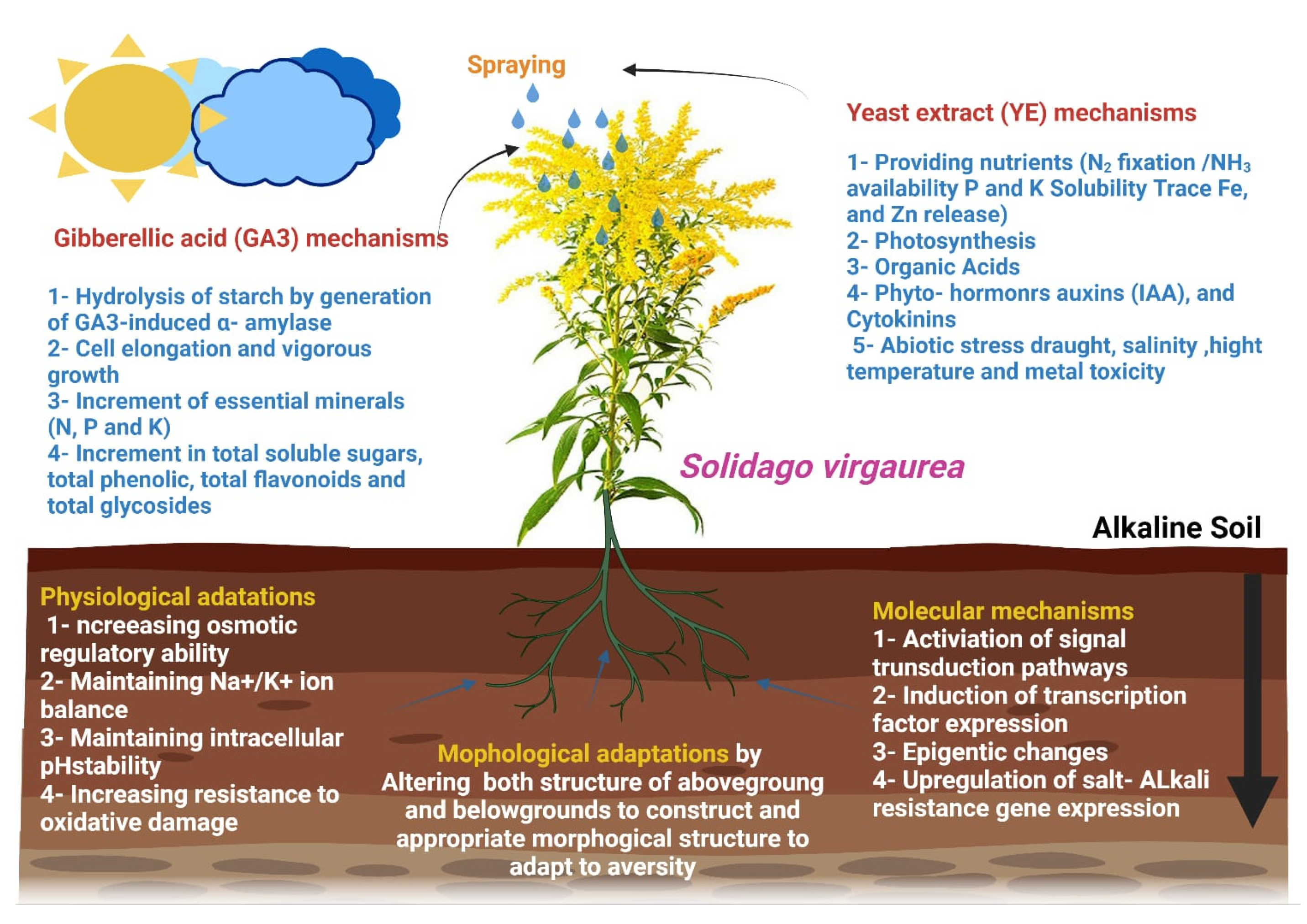
4. Discussion
The alkalization of soil has become a global environmental problem and is an important factor limiting agricultural productivity [
44]. Using biological safety compounds to improve plant productivity and quality has recently received much attention. Bio-stimulants improve plant growth and development by enhancing photosynthesis, endogenous hormones, ion uptake, nucleic acid, and protein synthesis, among other metabolic activities [
45]. In the current study, spraying Solidago plants with yeast extract (YE) and gibberellic acid (GA
3) individually or in combination markedly enhanced the morphological parameters, physiological responses, macronutrients and biochemical determinations compared to the non-treated plants.
The Increment in growth traits as a result of YE might be due to the presence of different macro and micronutrients, growth regulators, proteins, and vitamins (especially vitamin B) that encourage the plant to produce dry matter [
46]. It is also a natural source of cytokinins, which promote cell proliferation and differentiation while also governing shoot and root morphogenesis, chloroplast maturation, protein and nucleic acid synthesis [
47], or may be due to YE being high in tryptophan, which is a precursor to indole acetic acid (IAA). This substance promotes cell division and elongation [
48]. In addition, the increment might be due to the various roles of amino acids in the protein structure of several plant enzymes that are required for vegetative development [
49]. The improvement of physiological properties in response to the foliar application of YE may be attributed to its bio-regulator role in plants, affecting the balance of photosynthesis and photorespiration [
50] and delaying the leaf senescence by reducing the degradation of chlorophyll, improving protein and RNA synthesis [
51]. The importance of YE at different concentrations on the accumulation of the N, P and K % in leaves may be due to its diverse range of amino acids and vitamins. In addition, YE is a natural source of many growth components as a protective agent, as well as the majority of nutritional elements (Na, Ca, Fe, K, P, S, Mg, Zn, and Si), cytokinins, and several organic compounds [
23]. Furthermore, the positive effect of YE on promoting vegetative growth could explain why the concentration of nutritional elements in leaves is increasing [
52]. YE stimulated the production of endogenous hormones, which led to the accumulation of secondary metabolites such as total soluble sugars, phenolic, flavonoids, and glycosides [
53]. YE plays a major role in increasing carbon dioxide release through the fermentation process, which results in an increase in photosynthetic pigments and successfully activates the photosynthesis process. Alternatively, this might have been due to it promoting cell division and cell elongation, resulting in an increased leaf area [
54] as a result of the biosynthesis of carbohydrates being accelerated [
55]. Similar reports were earlier published by [
56] on
flax plants [
52], wheat plants [
22], Chinese carnation, and [
24] white lupine plants.
On the other hand, plants sprayed with GA
3 also caused an increase in plant growth traits. This impact could be explained by gibberellic acid’s ability to boost auxin levels, resulting in increased cell division and elongation [
57]. Furthermore, the mechanism involves the hydrolysis of starch as a result of the generation of GA
3-induced α- amylase, which could raise the concentration of sugars in the cell sap, hence, elevating the osmotic pressure; water enters the cell, causing the cell wall to stretch [
58]. Moreover, it is quoted that the meristematic region’s increasing size and the fraction of cells undergoing division [
59] contribute to cell elongation and vigorous growth. Foliar application with GA
3 also caused an increase in pigments, i.e., chlorophyll a, b, and total carotenoids. This effect may be due to GA
3’s effective role in preventing chloroplast and chlorophyll degradation, resulting in a reduction in leaf senescence and yellowing due to increased chlorophyll synthesis and chloroplast development [
57]. Furthermore, under the impact of GA
3, GA
3 in the chloroplast membrane enables photosynthesis to be regulated, more light to be trapped, a larger leaf surface, and increased leaf longevity [
60]. It is worth mentioning that the increment of essential minerals (N, P, and K) by increasing the concentration of GA
3 may be a result of augmenting the leaves’ dry weights to a greater extent than its effect on reducing N, P, and K percentages in the leaves [
61]. Therefore, the increment in total soluble sugars, total phenolic, total flavonoids and total glycosides as a result of GA
3 application might be due to the role in increasing the amount of chlorophyll in the leaves, which was reflected in raising the photosynthetic rate and, consequently, the accumulation of secondary metabolites increased [
62]. Otherwise, it is likely that increased photosynthetic CO
2 fixation may increase the amount of carbohydrates available for metabolism and export them to the sink [
63]. In harmony with these results were those emphasized by [
64] on
chrysanthemums [
65], tuberose plants, and [
1] golden rod plants.
Soil osmotic pressure is increased during saline-alkali stress due to sodium ion build-up. To sustain intracellular water potential, plant cells generate and store proline, soluble proteins, betaine, sugar, polyols, and polyamines [
66]. These compounds modify water’s solvent characteristics, stabilize the internal osmotic potential, increase protein folding stability, and safeguard the macromolecular structure [
37]. Sorghum seedlings respond to salt-alkali conditions by synthesizing proline and soluble proteins. Wheat responded to salt and alkali stress by increasing its proline, soluble sugar, and polyol (sorbitol) content. Furthermore, under saline-alkali stress, many plants release considerable quantities of organic acids, which may buffer intracellular pH and ion balance [
66]. Proton pump H+-ATPase may have a role in organic acid release from roots under NaHCO
3 stress, according to related research [
38]. Finally, saline-alkali stress causes osmotic and ionic stress, which generates reactive oxygen species (ROS) such as H
2O
2 and hydroxyl radicals [
39]. Alkali stress damaged rice cell membranes, which boosted the plant’s antioxidant defence mechanism [
40]. Taken together, our findings demonstrate that plants of various species and cultivars within the same species may alter their osmotic adjustment components in response to salt-alkali stress, as shown in
Figure 7.



 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}