


Stress Induced Activation of LTR Retrotransposons in the Drosophila melanogaster Genome


Abstract
:1. Introduction
2. Materials and Methods
2.1. Drosophila melanogaster Strains and Cultivation Conditions
2.2. Induction of Oxidative and Chronic Heat Stress
2.3. RNA Isolation, Reverse Transcription, and Real-Time PCR
2.4. DNA Isolation and Nanopore Sequencing
2.5. Evaluation of the Number of Copies of Transposable Elements by PCR
3. Results
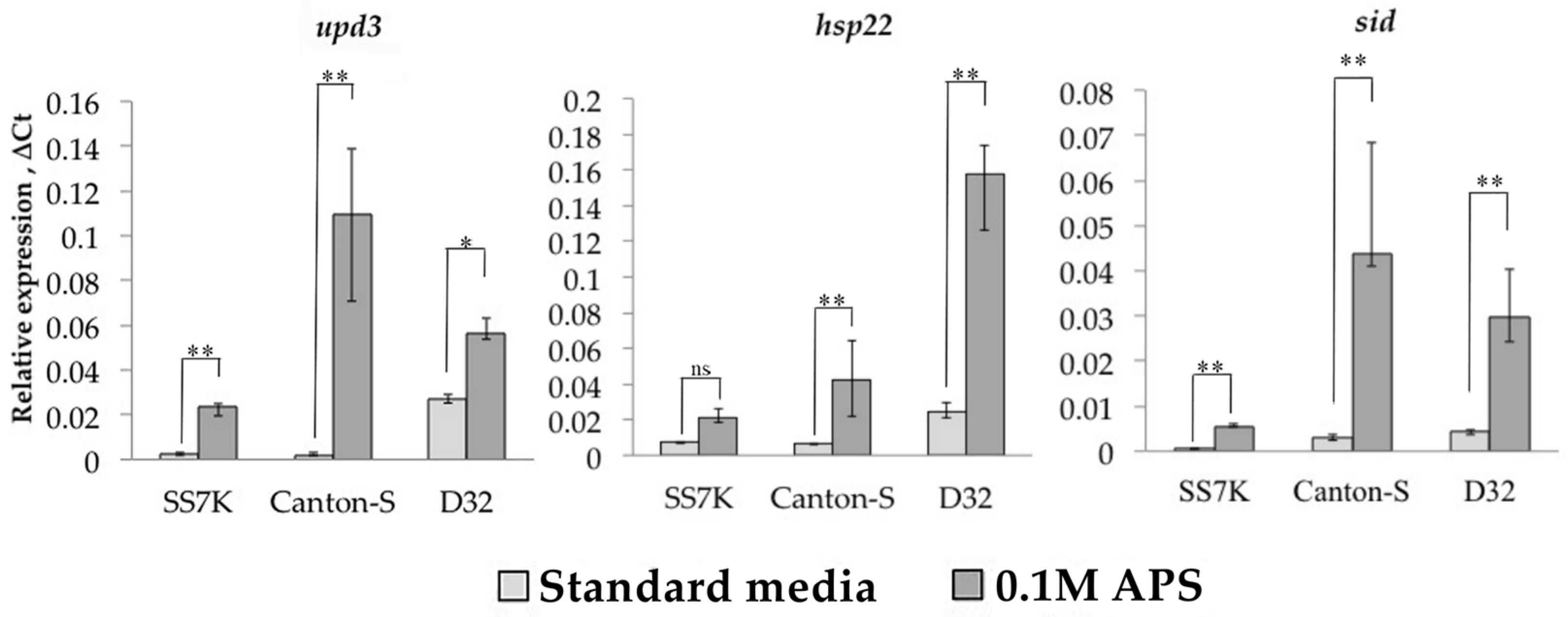
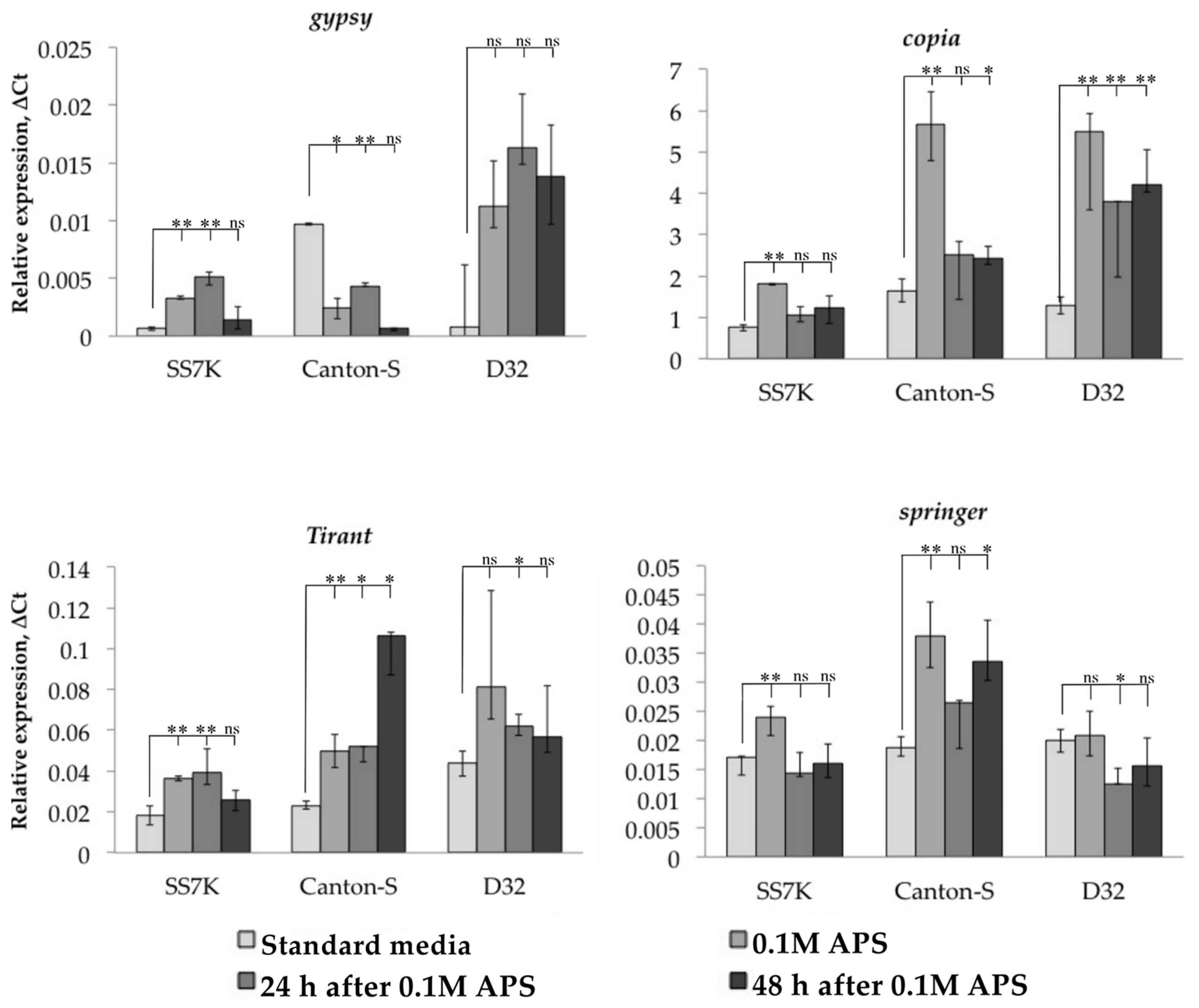
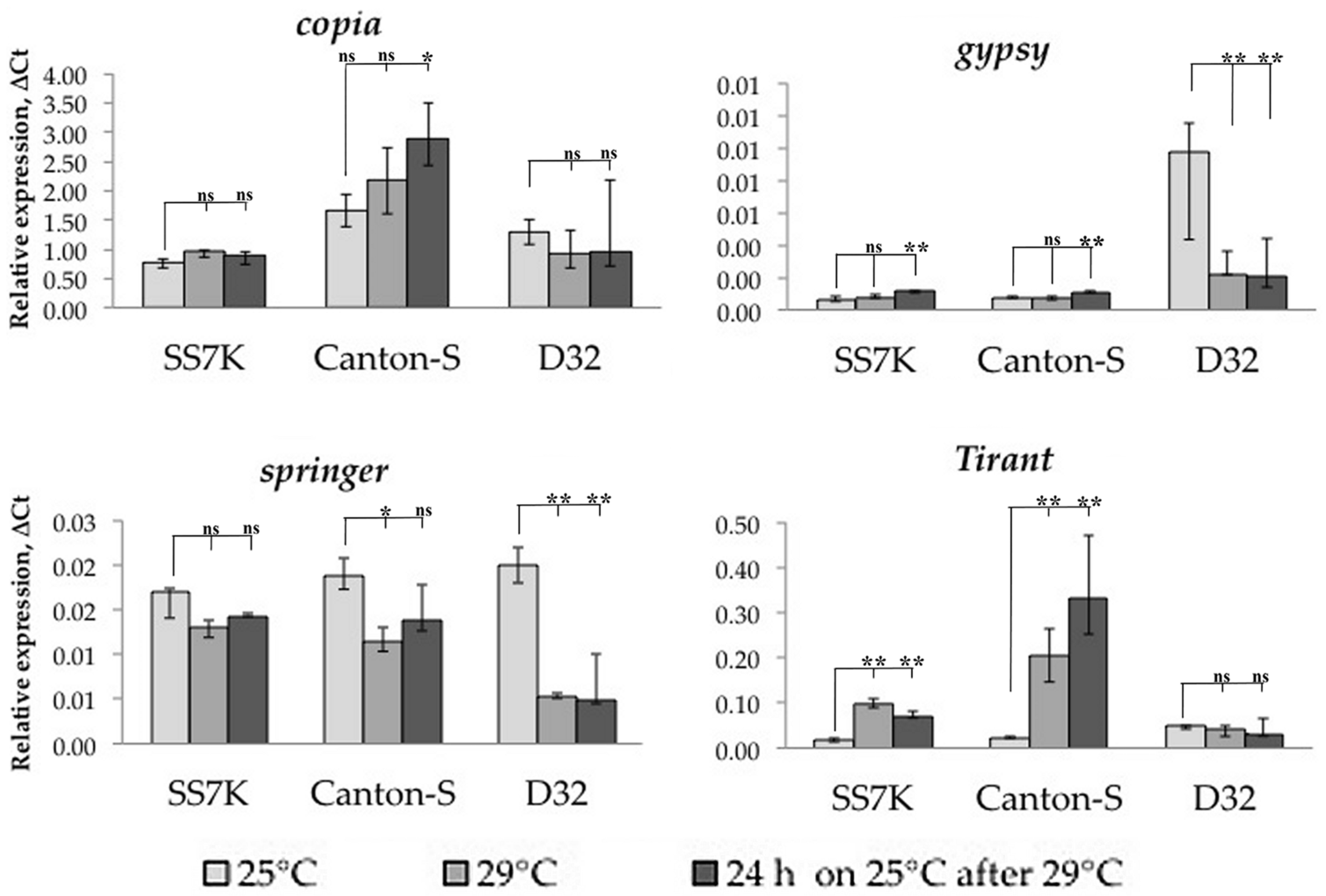
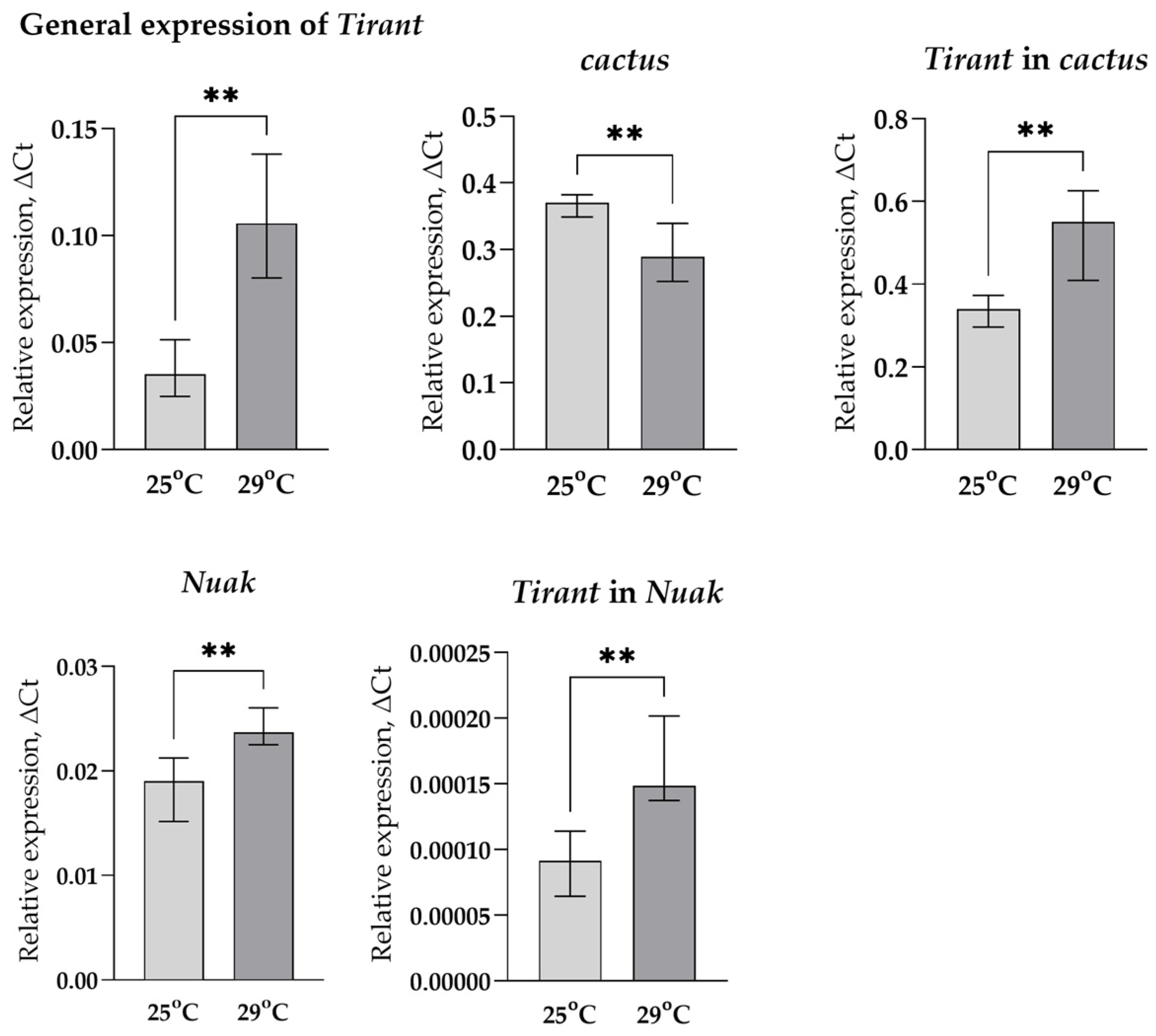
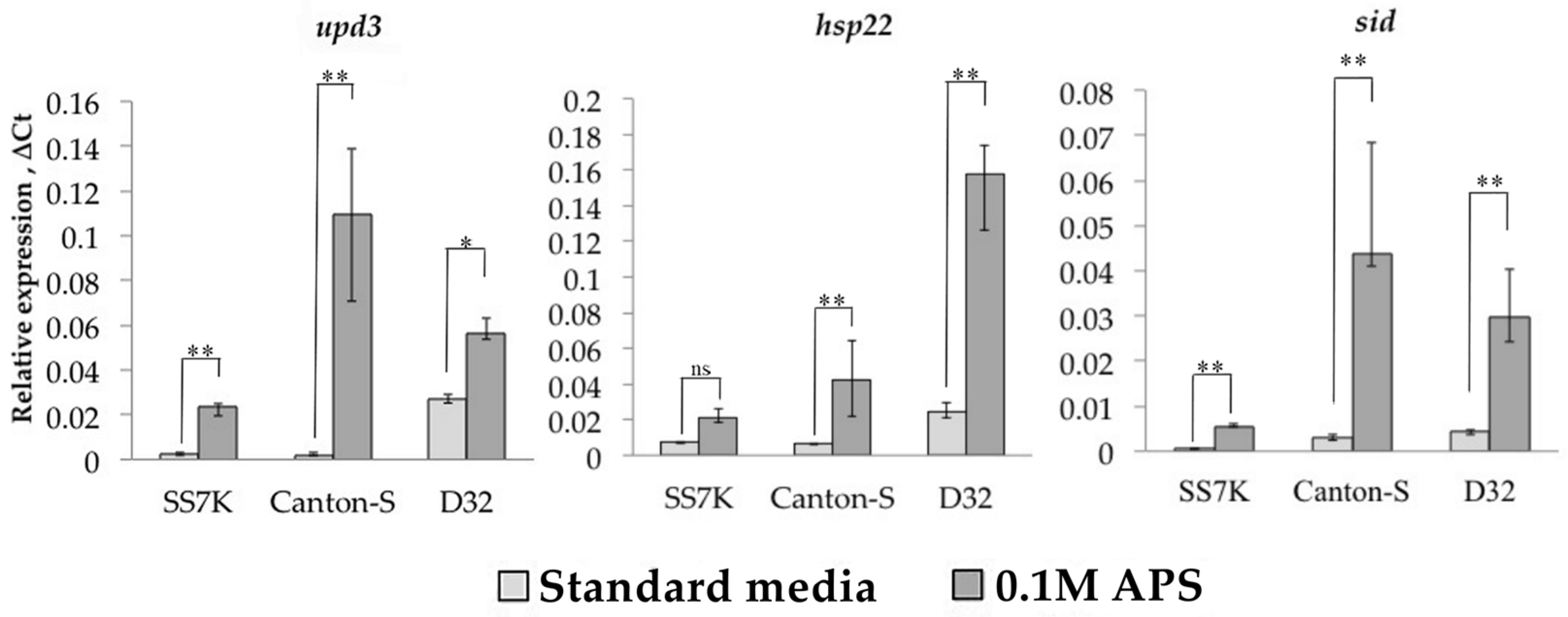
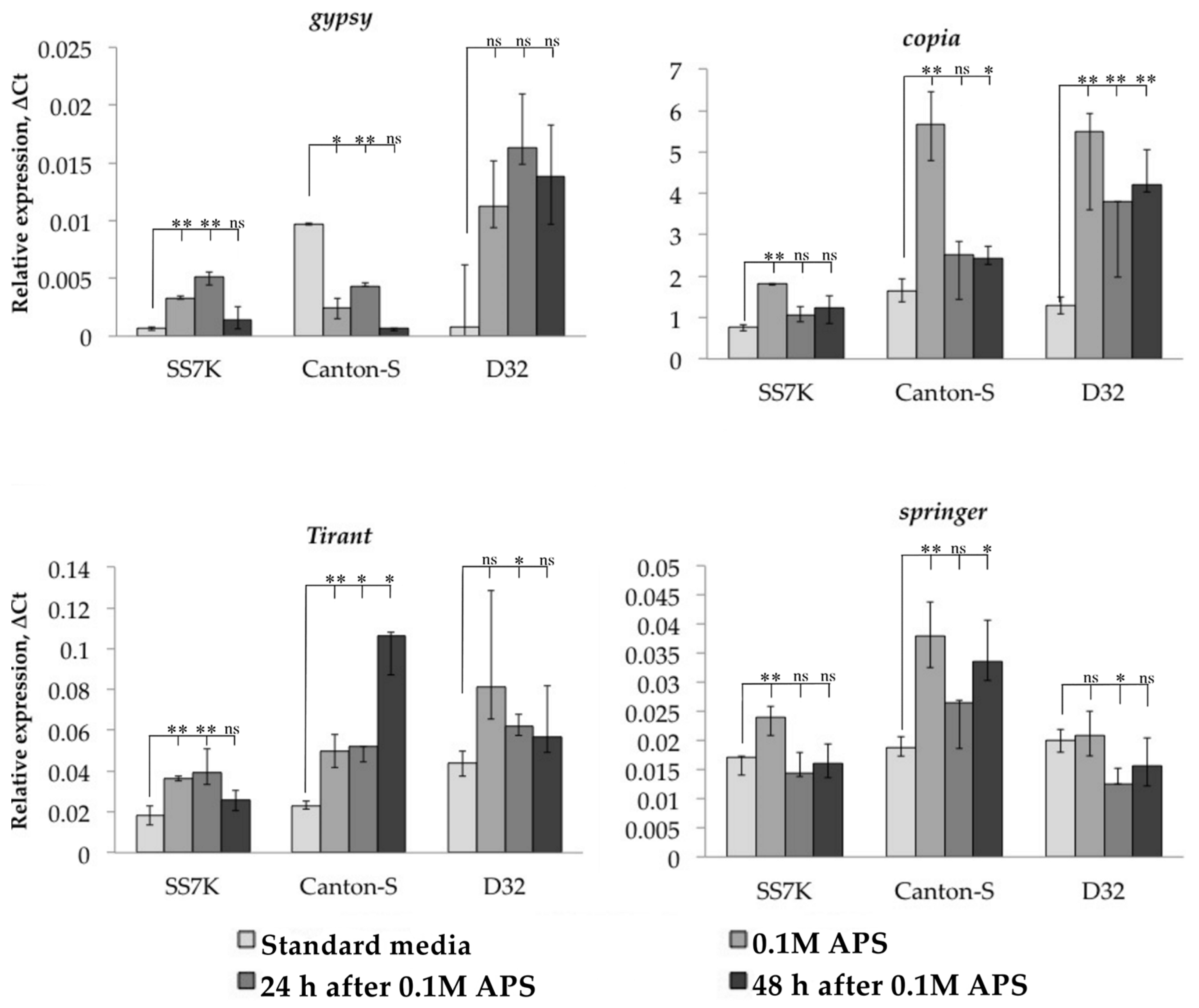
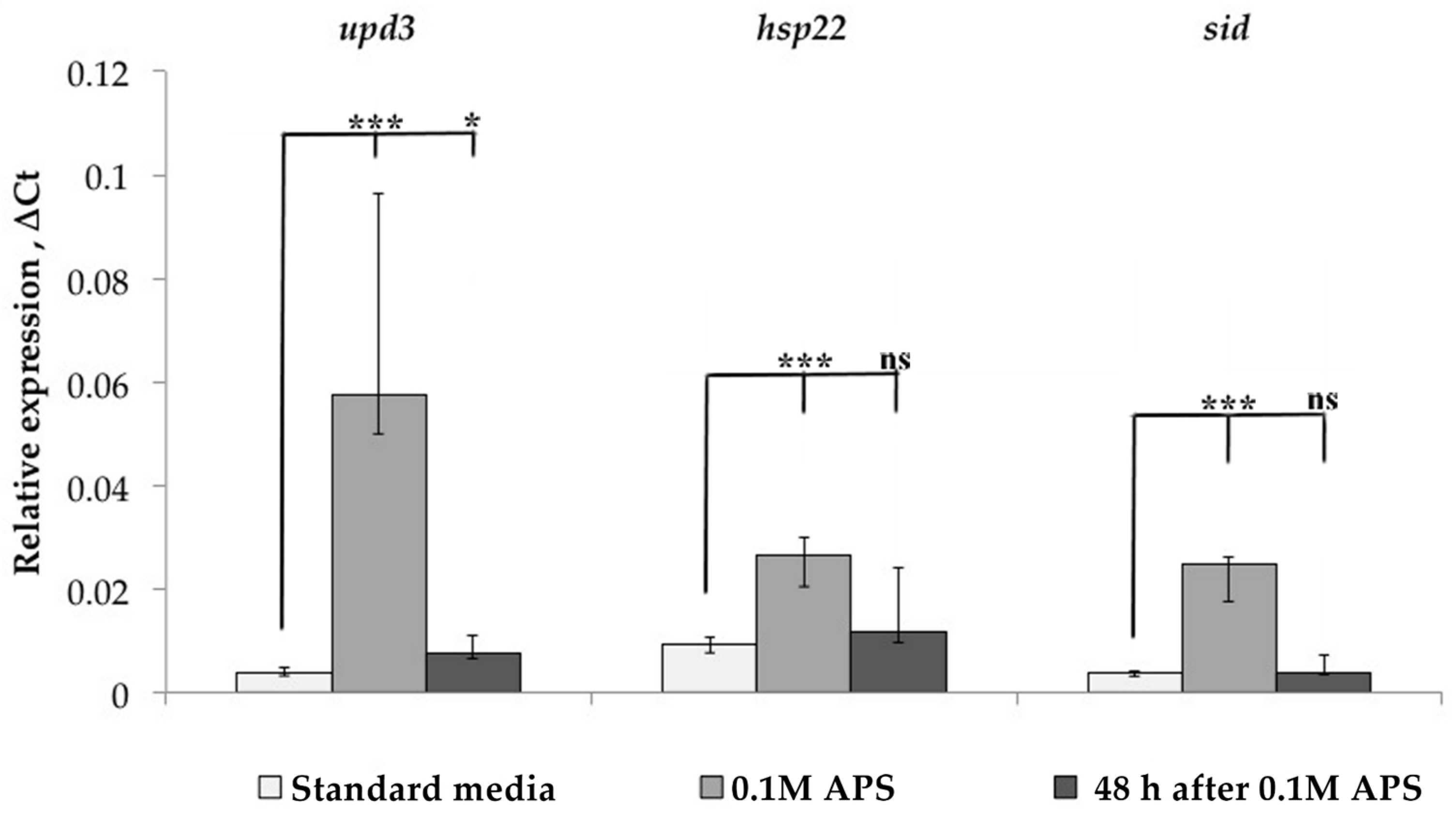
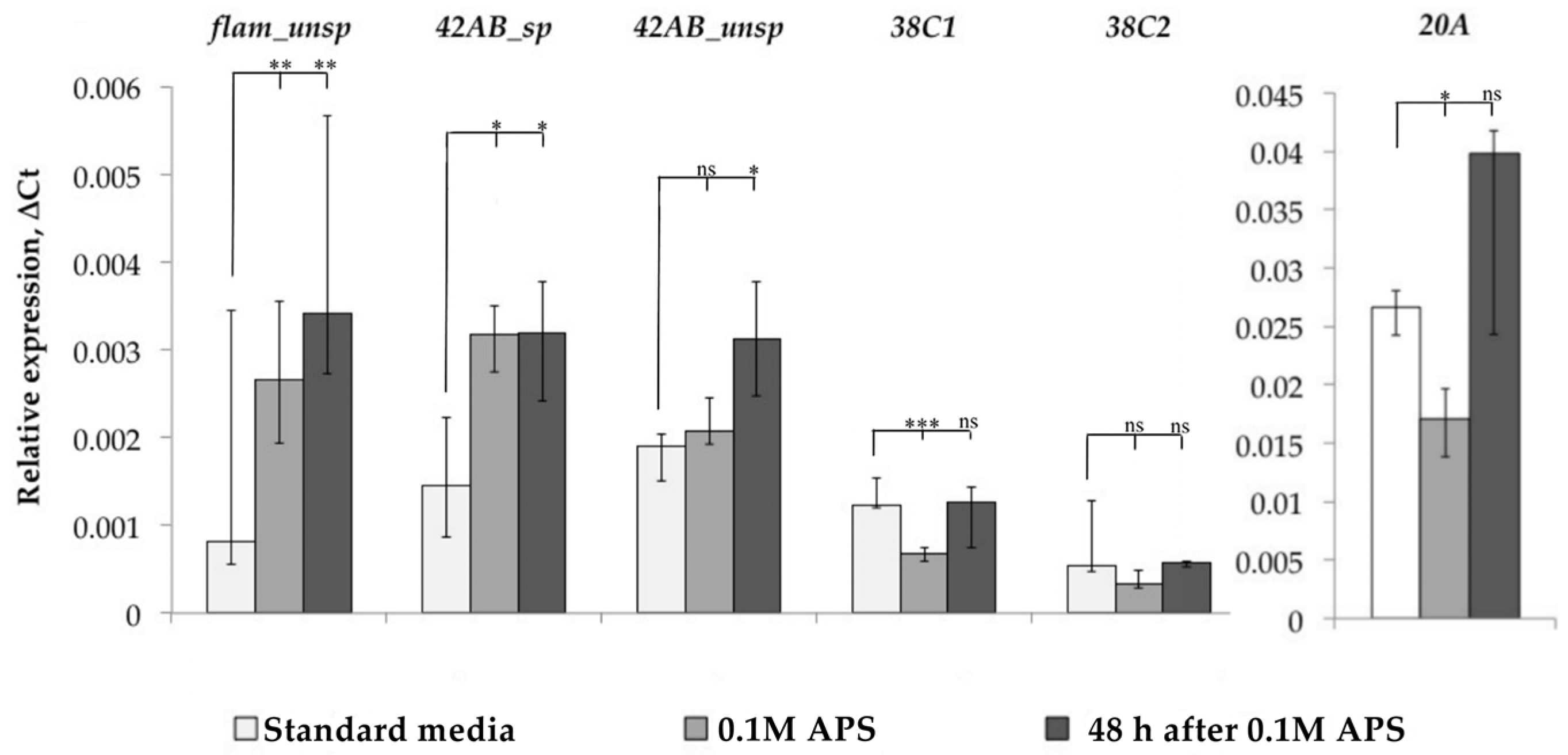
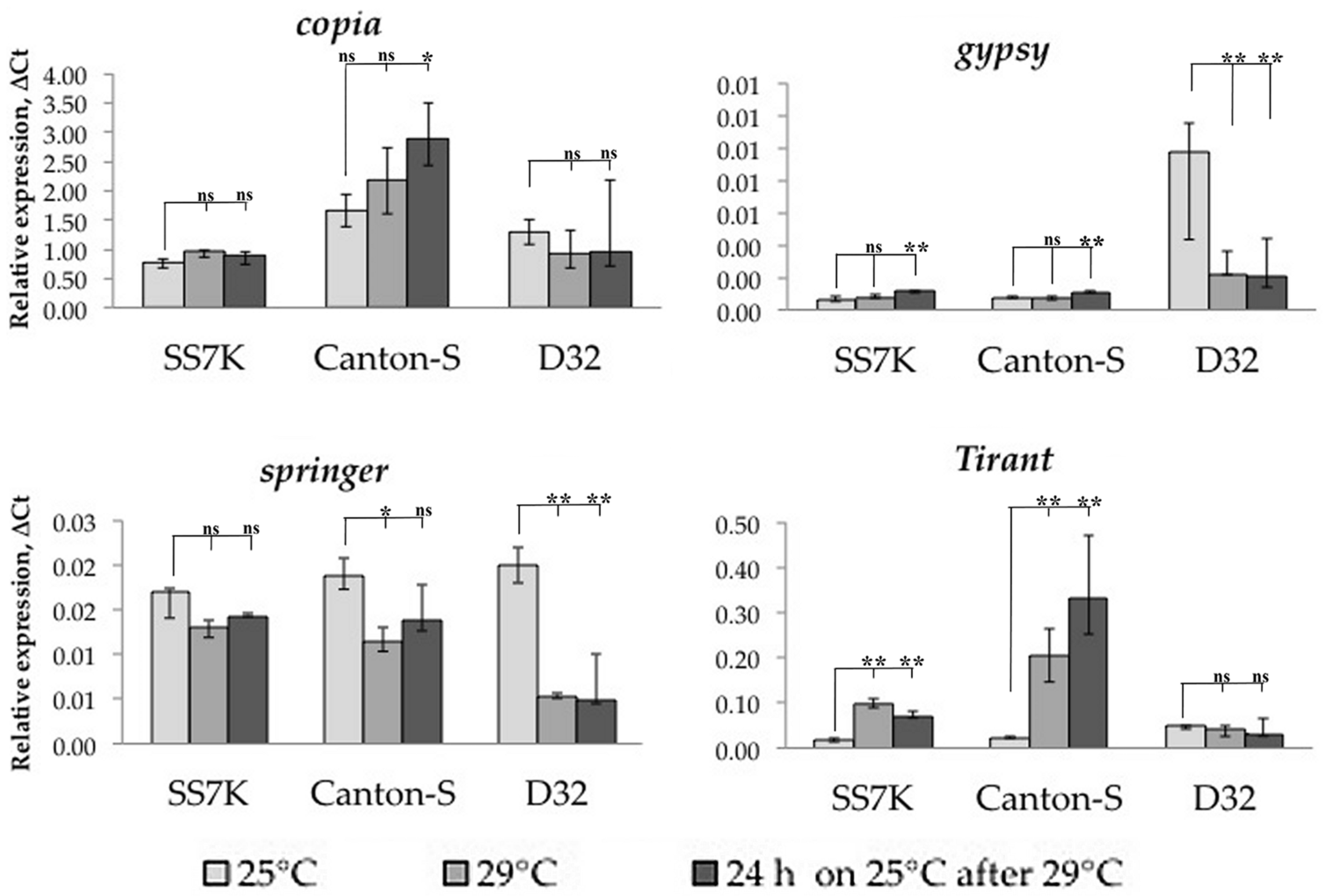
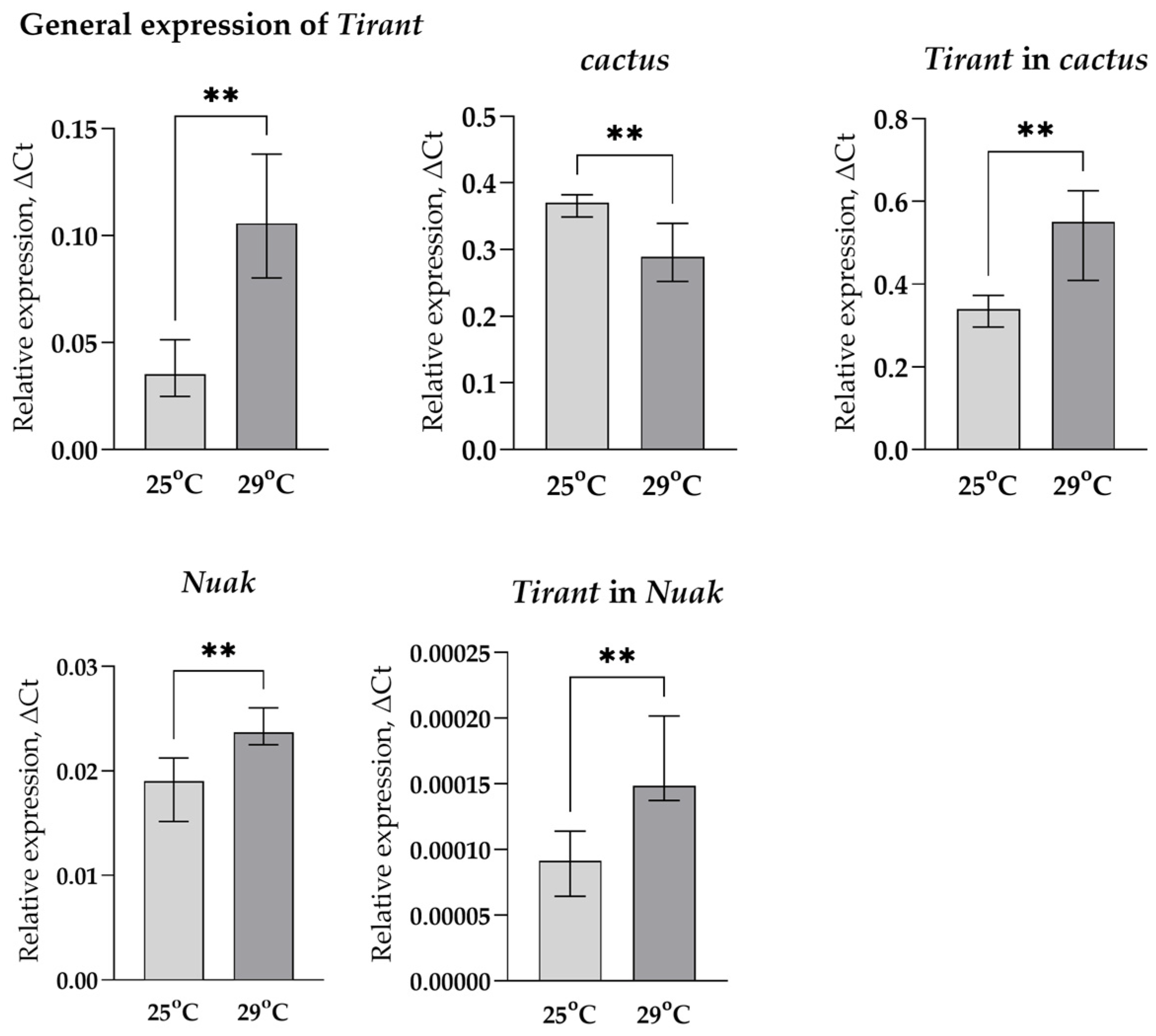
3.1. Transcription Analysis of LTR Retrotransposons under Oxidative and Chronic Heat Stress Conditions
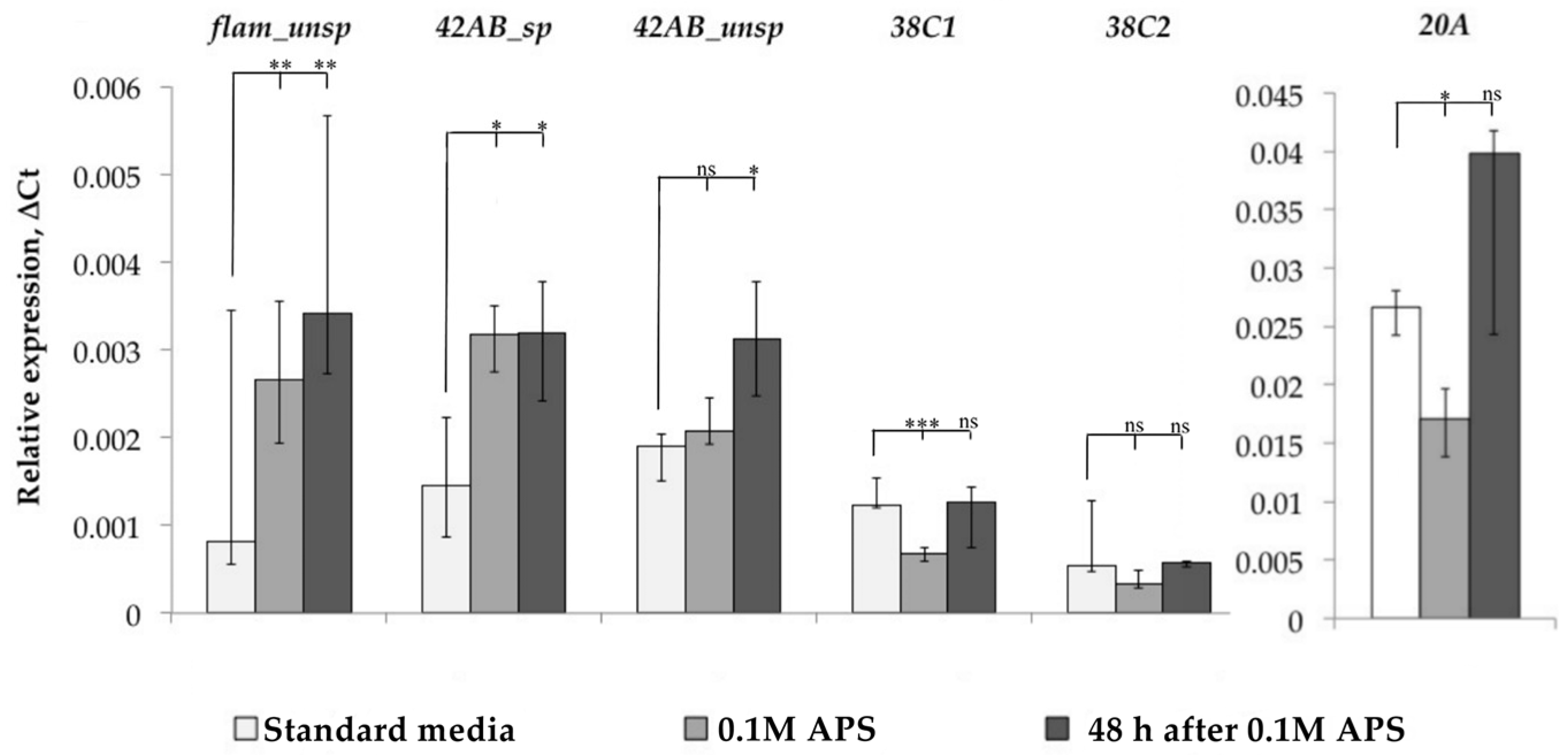
3.2. Analysis of the Relationship between Transposable Element Copy Number and Their Position in the Genome with Their Transcription Activation
3.3. Search for Transcription Factor Binding Sites in the Regulatory Regions of LTR Retrotransposons
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casacuberta, E.; González, J. The Impact of Transposable Elements in Environmental Adaptation. Mol. Ecol. 2013, 22, 1503–1517. [Google Scholar] [CrossRef] [PubMed]
- Kidwell, M.G. Transposable Elements and the Evolution of Genome Size in Eukaryotes. Genetica 2002, 115, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Park, E.G.; Ha, H.; Lee, D.H.; Kim, W.R.; Lee, Y.J.; Bae, W.H.; Kim, H.-S. Genomic Analyses of Non-Coding RNAs Overlapping Transposable Elements and Its Implication to Human Diseases. Int. J. Mol. Sci. 2022, 23, 8950. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.S.; Hutvágner, G.; Haley, B.; Zamore, P.D. Evidence That SiRNAs Function as Guides, Not Primers, in the Drosophila and Human RNAi Pathways. Mol. Cell 2002, 10, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, M.P.G. What Makes Transposable Elements Move in the Drosophila Genome. Heredity 2012, 108, 461–468. [Google Scholar] [CrossRef]
- Théron, E.; Dennis, C.; Brasset, E.; Vaury, C. Distinct Features of the PiRNA Pathway in Somatic and Germ Cells: From PiRNA Cluster Transcription to PiRNA. Process. Amplif. 2014, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Yashiro, R.; Murota, Y.; Nishida, K.M.; Yamashiro, H.; Fujii, K.; Ogai, A.; Yamanaka, S.; Negishi, L.; Siomi, H.; Siomi, M.C. Piwi Nuclear Localization and Its Regulatory Mechanism in Drosophila Ovarian Somatic Cells. Cell Rep. 2018, 23, 3647–3657. [Google Scholar] [CrossRef]
- Yamanaka, S.; Siomi, M.C.; Siomi, H. PiRNA Clusters and Open Chromatin Structure. Mob. DNA 2014, 5, 22. [Google Scholar] [CrossRef]
- Schnabl, J.; Wang, J.; Hohmann, U.; Gehre, M.; Batki, J.; Andreev, V.I.; Purkhauser, K.; Fasching, N.; Duchek, P.; Novatchkova, M.; et al. Molecular Principles of Piwi-Mediated Cotranscriptional Silencing through the Dimeric SFiNX Complex. Genes Dev. 2021, 35, 392–409. [Google Scholar] [CrossRef]
- Onishi, R.; Sato, K.; Murano, K.; Negishi, L.; Siomi, H.; Siomi, M.C. Piwi Suppresses Transcription of Brahma-Dependent Transposons via Maelstrom in Ovarian Somatic Cells. Sci. Adv. 2020, 6, eaaz7420. [Google Scholar] [CrossRef]
- Mohn, F.; Sienski, G.; Handler, D.; Brennecke, J. The rhino-deadlock-cutoff complex licenses noncanonical transcription of dual-strand piRNA clusters in Drosophila. Cell 2014, 157, 1364–1379. [Google Scholar] [CrossRef] [PubMed]
- Aravin, A.A.; Hannon, G.J.; Brennecke, J. The Piwi-PiRNA Pathway Provides an Adaptive Defense in the Transposon Arms Race. Science 2007, 318, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete Small RNA-Generating Loci as Master Regulators of Transposon Activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [PubMed]
- Siomi, M.C.; Sato, K.; Pezic, D.; Aravin, A.A. PIWI-Interacting Small RNAs: The Vanguard of Genome Defence. Nat. Rev. Mol. Cell Biol. 2011, 12, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Siomi, M.C. Two Distinct Transcriptional Controls Triggered by Nuclear Piwi-PiRISCs in the Drosophila PiRNA Pathway. Curr. Opin. Struct. Biol. 2018, 53, 69–76. [Google Scholar] [CrossRef]
- Hur, J.K.; Luo, Y.; Moon, S.; Ninova, M.; Marinov, G.K.; Chung, Y.D.; Aravin, A.A. Splicing-Independent Loading of TREX on Nascent RNA Is Required for Efficient Expression of Dual-Strand PiRNA Clusters in Drosophila. Genes Dev. 2016, 30, 840–855. [Google Scholar] [CrossRef]
- Kneuss, E.; Munafò, M.; Eastwood, E.L.; Deumer, U.-S.; Preall, J.B.; Hannon, G.J.; Czech, B. Specialization of the Drosophila Nuclear Export Family Protein Nxf3 for PiRNA Precursor Export. Genes Dev. 2019, 33, 1208–1220. [Google Scholar] [CrossRef]
- Vieira, C.; Aubry, P.; Lepetit, D.; Bié Mont, C. A Temperature Cline in Copy Number for 412 but Not Roo/B104 Retrotransposons in Populations of Drosophila simulans. Proc. R. Soc. Lond. B 1998, 265, 1161–1165. [Google Scholar] [CrossRef]
- Oliveira, D.S.; Rosa, M.T.; Vieira, C.; Loreto, E.L.S. Oxidative and Radiation Stress Induces Transposable Element Transcription in Drosophila melanogaster. J. Evol. Biol. 2021, 34, 628–638. [Google Scholar] [CrossRef]
- McClintock, B. The Significance of Responses of the Genome to Challenge. Science 1984, 226, 792–801. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, Oxidative Stress and the Biology of Ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Meadows, S.; Sharp, L.; Jan, L.Y.; Jan, Y.N. Genome-Wide Study of Aging and Oxidative Stress Response in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2000, 97, 13726–13731. [Google Scholar] [CrossRef] [PubMed]
- Tettweiler, G.; Miron, M.; Jenkins, M.; Sonenberg, N.; Lasko, P.F. Starvation and Oxidative Stress Resistance in Drosophila Are Mediated through the EIF4E-Binding Protein, D4E-BP. Genes Dev. 2005, 19, 1840–1843. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Cañas, J.L.; Horvath, V.; Aguilera, L.; González, J. Diverse Families of Transposable Elements Affect the Transcriptional Regulation of Stress-Response Genes in Drosophila melanogaster. Nucleic Acids Res. 2019, 47, 6842–6857. [Google Scholar] [CrossRef] [PubMed]
- Moschetti, R.; Palazzo, A.; Lorusso, P.; Viggiano, L.; Massimiliano Marsano, R. “What You Need, Baby, I Got It”: Transposable Elements as Suppliers of Cis-Operating Sequences in Drosophila. Biology 2020, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.I.; Belyaeva, E.S.; Larkina, Z.G.; Aslanyan, M.M. Genetic Instability and Transposition of the Mobile Element MDG4 in the Drosophila melanogaster Mutator Line. Sov. Genet. 1989, 25, 1747–1756. [Google Scholar]
- Zhang, Z.; Wang, J.; Schultz, N.; Zhang, F.; Parhad, S.S.; Tu, S.; Vreven, T.; Zamore, P.D.; Weng, Z.; Theurkauf, W.E. The HP1 Homolog Rhino Anchors a Nuclear Complex That Suppresses PiRNA Precursor Splicing. Cell 2014, 157, 1353–1363. [Google Scholar] [CrossRef]
- Nefedova, L.N.; Urusov, F.A.; Romanova, N.I.; Shmel’kova, A.O.; Kim, A.I. Study of the Transcriptional and Transpositional Activities of the Tirant Retrotransposon in Drosophila melanogaster Strains Mutant for the flamenco Locus. Russ. J. Genet. 2012, 48, 1089–1096. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]
- Ewing, A.D.; Smits, N.; Sanchez-Luque, F.J.; Faivre, J.; Brennan, P.M.; Richardson, S.R.; Cheetham, S.W.; Faulkner, G.J. Nanopore Sequencing Enables Comprehensive Transposable Element Epigenomic Profiling. Mol. Cell 2020, 80, 915–928.e5. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Huang, C.-H. LASAGNA-Search: An integrated web tool for transcription factor binding site search and visualization. BioTechniques 2013, 54, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Gramates, L.S.; Agapite, J.; Attrill, H.; Calvi, B.R.; Crosby, M.A.; dos Santos, G.; Goodman, J.L.; Goutte-Gattat, D.; Jenkins, V.K.; Kaufman, T.; et al. FlyBase: A guided tour of highlighted features. Genetics 2022, 220, iyac035. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Makhnovskii, P.; Balakireva, Y.; Nefedova, L.; Lavrenov, A.; Kuzmin, I.; Kim, A. Domesticated Gag Gene of Drosophila LTR Retrotransposons Is Involved in Response to Oxidative Stress. Genes 2020, 11, 396. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Bai, Y.; Li, K.; Rong, Y.S. Taming Active Transposons at Drosophila Telomeres: The Interconnection between HipHop’s Roles in Capping and Transcriptional Silencing. PLoS Genet. 2021, 17, e1009925. [Google Scholar] [CrossRef] [PubMed]
- Cappucci, U.; Noro, F.; Casale, A.M.; Fanti, L.; Berloco, M.; Alagia, A.A.; Grassi, L.; Le Pera, L.; Piacentini, L.; Pimpinelli, S. The Hsp70 Chaperone Is a Major Player in Stress-Induced Transposable Element Activation. Proc. Natl. Acad. Sci. USA 2019, 116, 17943–17950. [Google Scholar] [CrossRef] [PubMed]
- Perrat, P.N.; DasGupta, S.; Wang, J.; Theurkauf, W.; Weng, Z.; Rosbash, M.; Waddell, S. Transposition-Driven Genomic Heterogeneity in the Drosophila. Brain Sci. 2013, 340, 91–95. [Google Scholar] [CrossRef]
- Jedlicka, P.; Mortin, M.A.; Wu, C. Multiple Functions of Drosophila Heat Shock Transcription Factor In Vivo. EMBO J. 1997, 16, 2452–2462. [Google Scholar] [CrossRef]
- Wu, K.; Tang, Y.; Zhang, Q.; Zhuo, Z.; Sheng, X.; Huang, J.; Ye, J.; Li, X.; Liu, Z.; Chen, H. Aging-Related Upregulation of the Homeobox Gene Caudal Represses Intestinal Stem Cell Differentiation in Drosophila. PLoS Genet. 2021, 17, e1009649. [Google Scholar] [CrossRef]
- Ogienko, A.A.; Yarinich, L.A.; Fedorova, E.V.; Dorogova, N.V.; Bayborodin, S.I.; Baricheva, E.M.; Pindyurin, A.V. GAGA Regulates Border Cell Migration in Drosophila. Int. J. Mol. Sci. 2020, 21, 7468. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Mohammad, A.; Pazdernik, N.; Huang, H.; Bowman, B.; Tycksen, E.; Schedl, T. GLP-1 Notch—LAG-1 CSL Control of the Germline Stem Cell Fate Is Mediated by Transcriptional Targets Lst-1 and Sygl-1. PLoS Genet. 2020, 16, e1008650. [Google Scholar] [CrossRef] [PubMed]
- Van Oosten-Hawle, P. Organismal Roles of Hsp90. Biomolecules 2023, 13, 251. [Google Scholar] [CrossRef] [PubMed]
- Stączek, S.; Cytryńska, M.; Zdybicka-Barabas, A. Unraveling the Role of Antimicrobial Peptides in Insects. Int. J. Mol. Sci. 2023, 24, 5753. [Google Scholar] [CrossRef] [PubMed]
- Loch, G.; Zinke, I.; Mori, T.; Carrera, P.; Schroer, J.; Takeyama, H.; Hoch, M. Antimicrobial Peptides Extend Lifespan in Drosophila. PLoS ONE 2017, 12, e0176689. [Google Scholar] [CrossRef] [PubMed]
- Salces-Ortiz, J.; Vargas-Chavez, C.; Guio, L.; Rech, G.E.; González, J. Transposable Elements Contribute to the Genomic Response to Insecticides in Drosophila melanogaster. Philos. Trans. R. Soc. B 2020, 375, 20190341. [Google Scholar] [CrossRef] [PubMed]
- Jachowicz, J.W.; Bing, X.; Pontabry, J.; Bošković, A.; Rando, O.J.; Torres-Padilla, M.-E. LINE-1 Activation after Fertilization Regulates Global Chromatin Accessibility in the Early Mouse Embryo. Nat. Genet. 2017, 49, 1502–1510. [Google Scholar] [CrossRef]
- Trizzino, M.; Kapusta, A.; Brown, C.D. Transposable Elements Generate Regulatory Novelty in a Tissue-Specific Fashion. BMC Genom. 2018, 19, 468. [Google Scholar] [CrossRef]
- Nampoothiri, S.S.; Rajanikant, G.K. Decoding the Ubiquitous Role of MicroRNAs in Neurogenesis. Mol. Neurobiol. 2017, 54, 2003–2011. [Google Scholar] [CrossRef]
- Mustafin, R.N.; Khusnutdinova, E.K. Involvement of Transposable Elements in Neurogenesis. Vestn. VOGiS 2020, 24, 209–218. [Google Scholar] [CrossRef]
- Kim, K.W. PIWI Proteins and PiRNAs in the Nervous System. Mol. Cells 2019, 42, 828–835. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward Primer | Reverse Primer |
|---|---|---|
| Genes | ||
| αTub84D | 5′-GTGCATGTTGTCCAACACCAC-3′ | 5′-AGAACTCTCCCTCCTCCATA-3′ |
| EloB | 5′-GCACAAACATACACACTCACG-3′ | 5′-TTTCCTACTTCGCTTGCACC-3′ |
| RpL40 | 5′-CTGCGTGGTGGTATCATTG-3′ | 5′-CAGGTTGTTGGTGTGTCC-3′ |
| hsp22 | 5′-CTTTTCACGCCTTCTTCCAC-3′ | 5′-GTGAGTTTGTAGCCATCCTTG-3′ |
| sid | 5′-GGAAGTGTTCAAGCGATTG-3′ | 5′-AGCAGATACAACGTCTGGTG-3′ |
| upd3 | 5′-AACGGCCAGAACCAGGAATC-3′ | 5′-GAGAGGGCAAACTGGGACAT-3′ |
| cactus | 5′-GATCTCAGCGAGGAGATAGTC-3′ | 5′-CTCCCTCCTCTTTCTCCTGC-3′ |
| Nuak | 5′-GGTTCCTGTTTCCCAGTTACTC-3′ | 5′-ATCACTTTGGTGGCATCCTTTT-3′ |
| BoYb | 5′-CCCAAGTTTCTCATGGTTTC-3′ | 5′-ACGAACTGCTCCCGAATATG-3′ |
| Clusters | ||
| flanemco-unspliced | 5′-CATCAGCTCAGCAGCAGTGTA-3′ | 5′-GACTTAACACTTACCGCTTGAAA-3′ |
| 42AB-spliced | 5′-GCAGTTGCCGTCTCTCCT-3′ | 5′-TGGGTCAAAGTGCAGCAGTT-3′ |
| 42AB unspliced | 5′-TACGGGAATATAATCGCAGCAGTT-3′ | |
| 38C1 | 5′-AATGGCTAGTTCGCTACCAGACAG-3′ | 5′-CGGGTCTTCTCTCAAACGCAATC-3′ |
| 38C2 | 5′-CACAAAATGGCCCGCTGGAAA-3′ | 5′-AGCCAAACCCTGTGTTGTGA-3′ |
| 20A | 5′-GCCTACGCAGAGGCCTAAGT-3′ | 5′-CAGATGTGGTCCAGTTGTGC-3′ |
| Retrotransposons | ||
| gypsy | 5′-CTGCTGAAAGACGGCATTATC-3′ | 5′-AGAACTTTGCCTTGCCCAGAT-3′ |
| Tirant | 5′-AACGCTATTCATTCTGCAAC-3′ | 5′-AGGGTTCTCCTAACTACGTC-3′ |
| copia | 5′-CTTCAGTGATGGACAACTG-3′ | 5′-CAGTGTAATCTCATGGTCAT-3′ |
| springer | 5′-CTAAATTCGCCATGGTACAGC-3′ | 5′-AGCCGAGGAGTAAATGAGTA-3′ |
| Tirant in cactus | 5′-GGGACCTTTCGCCTCAAC-3′ | 5′-GCCACTATTGTCTGCGATTT-3′ |
| Tirant in Nuak | 5′-GCCTGAGTTAACAAAGGTGAAC-3′ | 5′-GTAGGAATCAAAAAACTACACAACC-3′ |
| Strain | gypsy | copia | Tirant | springer | |
|---|---|---|---|---|---|
| Transposable Element | |||||
| SS7K | 3 | 44 | 5 | 6 | |
| CantonS | 3 | 55 | 4 | 6 | |
| D32 | 2 | 46 | 11 | 6 | |
| LTR Retrotransposon | Gene, Contaning TE Insertion | Insertion Position in the Gene | Collinearity with the Direction of the Gene Transcription | Response of Genes to Oxidative Stress and Heat Shock (According FlyBase) |
|---|---|---|---|---|
| SS7K | ||||
| copia | lr42a | Intron | No | No |
| IRSp53 | Intron | No | No | |
| bnl | Intron | No | No | |
| CG34353 | Intron | Yes | No | |
| Pde9 | Intron | No | Moderate to both stresses | |
| Tirant | cactus | 3′-UTR | No | High to both stresses |
| Nuak | Intron | No | No | |
| Canton-S | ||||
| copia | CG15431 | Intron | Yes | No |
| CG17684 | Intron | Yes | No | |
| SLO2 | Intron | No | No | |
| Pdk1 | Intron | No | Moderately high to stresses | |
| dpr6 | Intron | Yes | No | |
| CG45782 | Intron | No | No | |
| gypsy | CG42346 | Intron | No | No |
| springer | eyes | Intron | Yes | No |
| dnc | Intron | No | No | |
| CG30389 | Intron | No | High to oxidative stress, moderately high to heat shock | |
| CG33970 | Intron | No | No | |
| LTR Retrotransposon Insertion | Expression in Stress Conditions According to FlyBase | Strains | |||
|---|---|---|---|---|---|
| Position in the Gene | Collinearity with the Direction of the Gene Transcription | Oxidative Stress | Heat Shock | SS7K | Canton-S |
| copia | |||||
| CG3726 (intron) | No | Moderate | + | + | |
| toc (intron) | No | Moderately high | Moderate | + | + |
| for (intron) | No | Moderately high | Moderately high | + | + |
| Ir40a (intron) | Yes | + | + | ||
| CadN2 (intron) | Yes | + | + | ||
| CG44623 (3′UTR) | No | ||||
| mbl (intron) | No | ||||
| Nipped-A (intron) | No | ||||
| sallimus (intron) | Yes | ||||
| bbg (intron) | No | ||||
| Snap25 (intron) | No | Moderate | + | + | |
| Lasp, CG43954 (intron) | No | High | High | + | + |
| Eip63E (intron) | Yes | Moderate | + | + | |
| Myo81F (intron) | No | ||||
| beat-Vb (intron) | No | ||||
| CG34347 (intron) | Yes | ||||
| Gprk2 (intron) | No | Moderate | Moderate | + | + |
| Tirant | |||||
| CG42534(intron) | No | ||||
| CG42238(intron) | No | ||||
| Cipc (intron) | Yes | ||||
| SNF4Aγ (intron) | No | Moderately high | Moderate | + | + |
| Raf (intron) | No | Moderate | + | ||
| CG34417 (intron) | No | Moderately high | Moderate | ||
| CG32486 (intron) | No | Moderately high | Moderately high | + | |
| skd (intron) | No | Moderate | Moderate | + | + |
| Shab (intron) | No | Moderate | |||
| psq (intron) | No | Moderately high | Moderately high | + | |
| unc-5 (intron) | Yes | ||||
| chn (intron) | Yes | Moderate | Moderately high | ||
| CG11360 (intron) | No | ||||
| raw (intron) | Yes | ||||
| Fs(2)Ket (intron) | No | ||||
| LTR Retrotransposons | Transcription Factors | Binding Site |
|---|---|---|
| LTR | ||
| copia | HSF(M00164) | AGAGTGGTATTCTCT |
| gypsy | HSF(M00166) | CCTCGAGCAATGCAT |
| Bcd(T00063) | ACCTAATCTCCA | |
| Lag1(MA0193.1) | CTACTGG | |
| Tirant | Cad(T00079) | AGCATAATGA |
| Cad(T00079) | GGCCGAATG | |
| Deaf1(MA0185.1) | AGCATTCGGCCGGAA | |
| h(MA0449.1) | GCCACATGCC | |
| slbo(MA0244.1) | ATTACACA | |
| springer | Cad(T00079) | GGCCAAATGC |
| 5′UTR | ||
| copia | Zeste(M00283) | GAATTTGAGTGAAAA |
| Tirant | Cad(T00079) | GTCATAATTT |
| dl(M00043) | CGGTATGTCCA | |
| CF2-II(M00012) | CTATATACA | |
| CF2-II(M00012) | ATATATGTG | |
| slbo(MA0244.1) | ATTGCACA | |
| slbo(MA0244.1) | ATTGCAGA | |
| Cad(T00079) | GTACTAAAGA | |
| tll(MA0459.1) | TAAAGCCAAA | |
| tll(MA0459.1) | AGAAGTCGAC | |
| CF2-II(M00012) | CTATATGCA | |
| Dll(MA0187.1) | TAATTAC | |
| springer | cad(MA0216.1) | CTTATTG |
| ovo(MA0126.1) | TGTTACTGT | |
| dl(M00120) | TAAGAAAATCG | |
| dl(M00043) | CGGCATTTCCA | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milyaeva, P.A.; Kukushkina, I.V.; Kim, A.I.; Nefedova, L.N. Stress Induced Activation of LTR Retrotransposons in the Drosophila melanogaster Genome. Life 2023, 13, 2272. https://doi.org/10.3390/life13122272
Milyaeva PA, Kukushkina IV, Kim AI, Nefedova LN. Stress Induced Activation of LTR Retrotransposons in the Drosophila melanogaster Genome. Life. 2023; 13(12):2272. https://doi.org/10.3390/life13122272
Chicago/Turabian StyleMilyaeva, Polina A., Inna V. Kukushkina, Alexander I. Kim, and Lidia N. Nefedova. 2023. "Stress Induced Activation of LTR Retrotransposons in the Drosophila melanogaster Genome" Life 13, no. 12: 2272. https://doi.org/10.3390/life13122272
APA StyleMilyaeva, P. A., Kukushkina, I. V., Kim, A. I., & Nefedova, L. N. (2023). Stress Induced Activation of LTR Retrotransposons in the Drosophila melanogaster Genome. Life, 13(12), 2272. https://doi.org/10.3390/life13122272