
Antioxidant and Anticancer Activity of Vitis vinifera Extracts in Breast Cell Lines


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction Methodology
2.3. Total Polyphenol Content
2.4. Oligomeric Proanthocyanidins (OPC) Content
2.5. Cell Culture and Reagents
2.6. Extract Dilution
2.7. MTT Assay
2.8. DCFH-DA Assay
2.9. Cell Cycle Analysis
2.10. Annexin V/Propidium Iodide Staining
2.11. Total RNA Preparation and Real-Time Quantitative PCR (q-PCR)
2.12. Western Blot
2.13. Statistical Analysis
3. Results
3.1. Composition of Vitis vinifera Extracts
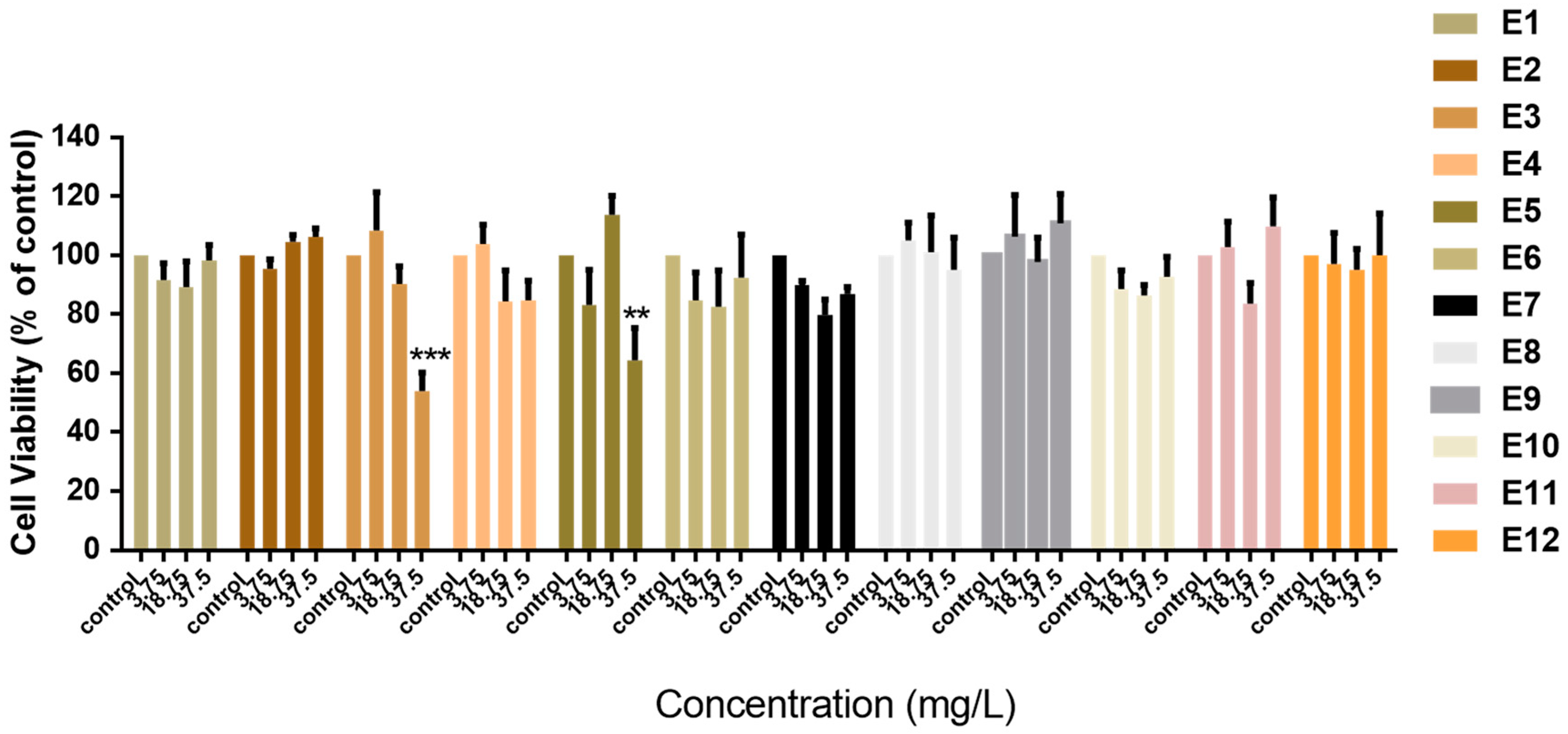
3.2. Effect of Vitis vinifera Extracts on the Viability of Normal Breast Cells
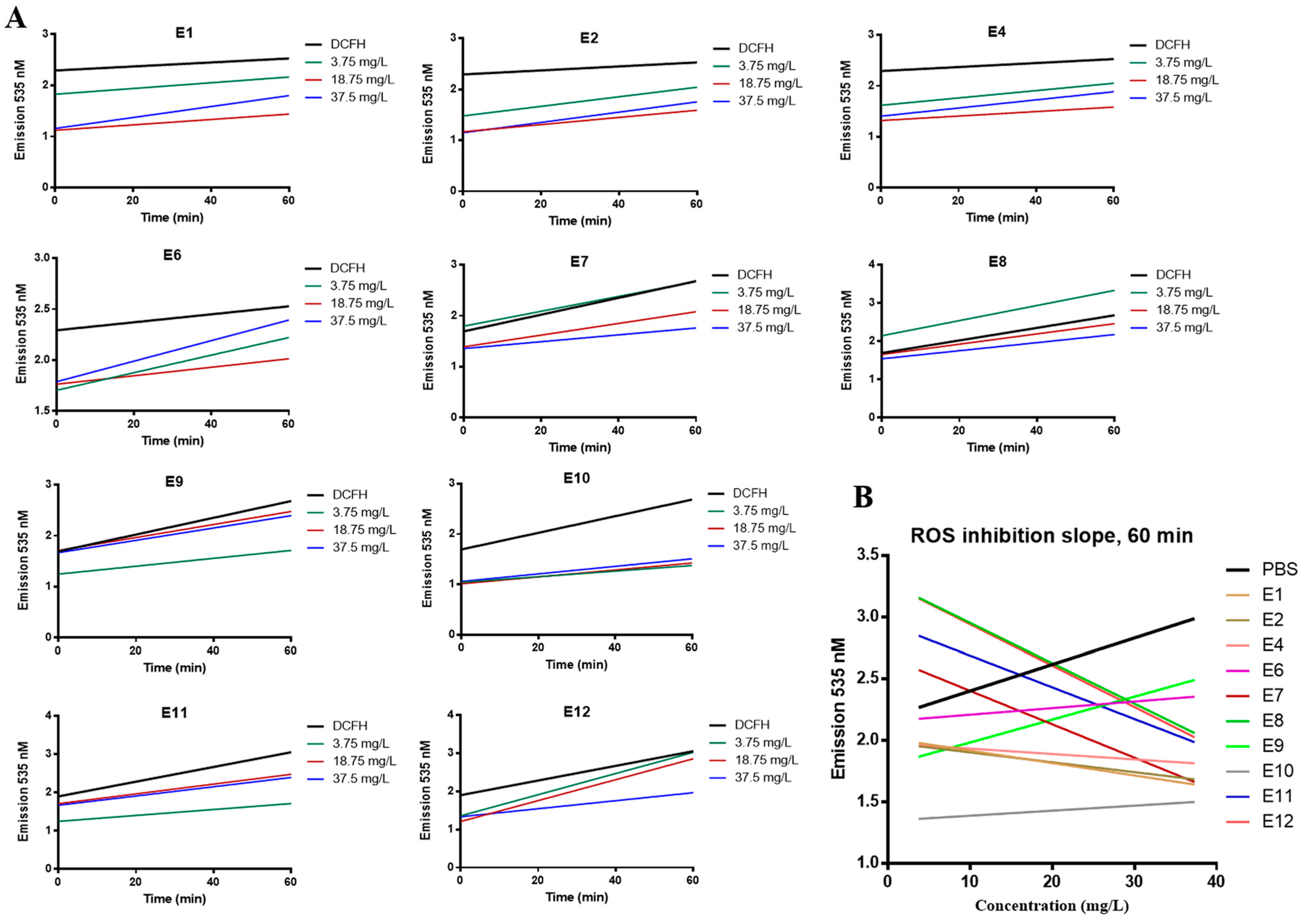
3.3. Antioxidant Activity of Vitis vinifera Extracts
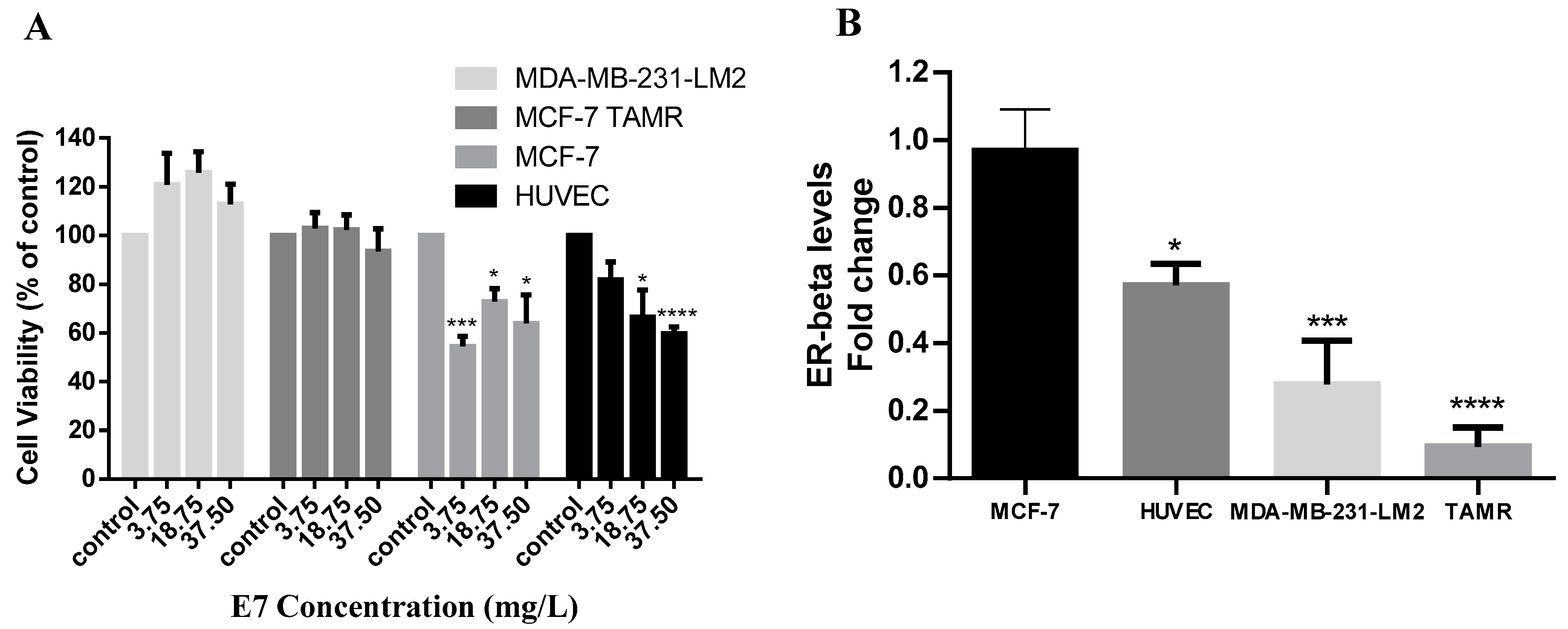
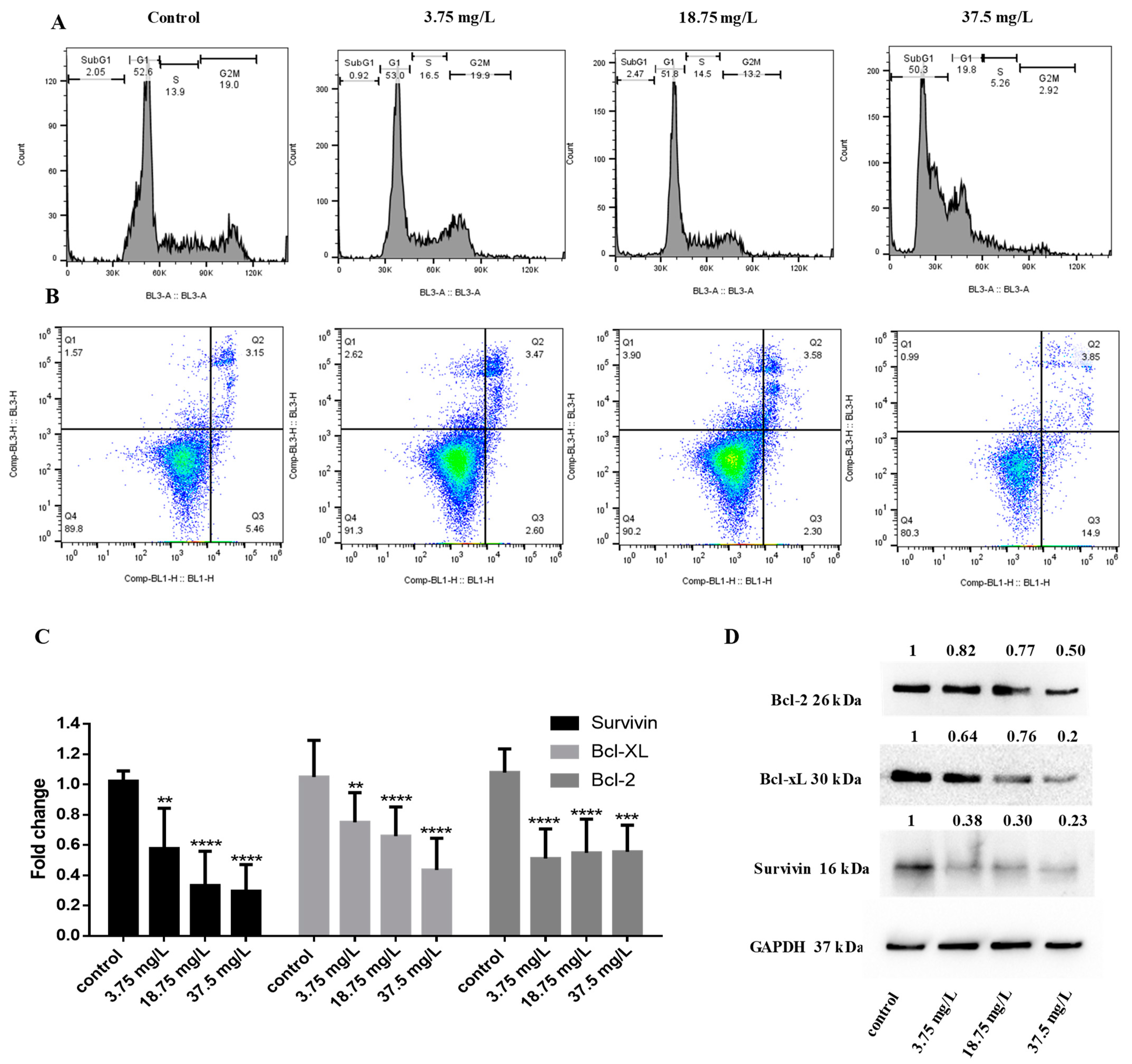
3.4. Investigation of the Anticancer Activity of Whole Seeds Extract from Vitis vinifera in Breast Cancer Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Neophytou, C.M.; Gregoriou, Y.; Constantinou, A.I. Pro-apoptotic Properties of Chemopreventive Agents. In Natural Products for Cancer Chemoprevention: Single Compounds and Combinations; Pezzuto, J.M., Vang, O., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 517–559. [Google Scholar]
- Nassiri-Asl, M.; Hosseinzadeh, H. Review of the Pharmacological Effects of Vitis vinifera (Grape) and its Bioactive Constituents: An Update. Phytother. Res. 2016, 30, 1392–1403. [Google Scholar] [CrossRef]
- Shrotriya, S.; Deep, G.; Gu, M.; Kaur, M.; Jain, A.K.; Inturi, S.; Agarwal, R.; Agarwal, C. Generation of reactive oxygen species by grape seed extract causes irreparable DNA damage leading to G2/M arrest and apoptosis selectively in head and neck squamous cell carcinoma cells. Carcinogenesis 2012, 33, 848–858. [Google Scholar] [CrossRef]
- Liang, Z.; Cheng, L.; Zhong, G.Y.; Liu, R.H. Antioxidant and antiproliferative activities of twenty-four Vitis vinifera grapes. PLoS ONE 2014, 9, e105146. [Google Scholar] [CrossRef]
- Yadav, D.; Kumar, A.; Kumar, P.; Mishra, D. Antimicrobial properties of black grape (Vitis vinifera L.) peel extracts against antibiotic-resistant pathogenic bacteria and toxin producing molds. Indian. J. Pharmacol. 2015, 47, 663–667. [Google Scholar] [CrossRef]
- Kantsadi, A.L.; Apostolou, A.; Theofanous, S.; Stravodimos, G.A.; Kyriakis, E.; Gorgogietas, V.A.; Chatzileontiadou, D.S.; Pegiou, K.; Skamnaki, V.T.; Stagos, D.; et al. Biochemical and biological assessment of the inhibitory potency of extracts from vinification byproducts of Vitis vinifera extracts against glycogen phosphorylase. Food Chem. Toxicol. 2014, 67, 35–43. [Google Scholar] [CrossRef]
- Sergazy, S.; Shulgau, Z.; Fedotovskikh, G.; Chulenbayeva, L.; Nurgozhina, A.; Nurgaziyev, M.; Krivyh, E.; Kamyshanskiy, Y.; Kushugulova, A.; Gulyayev, A.; et al. Cardioprotective effect of grape polyphenol extract against doxorubicin induced cardiotoxicity. Sci. Rep. 2020, 10, 14720. [Google Scholar] [CrossRef]
- Madrigal-Santillan, E.; Madrigal-Bujaidar, E.; Alvarez-Gonzalez, I.; Sumaya-Martinez, M.T.; Gutierrez-Salinas, J.; Bautista, M.; Morales-Gonzalez, A.; Garcia-Luna y Gonzalez-Rubio, M.; Aguilar-Faisal, J.L.; Morales-Gonzalez, J.A. Review of natural products with hepatoprotective effects. World J. Gastroenterol. 2014, 20, 14787–14804. [Google Scholar] [CrossRef] [PubMed]
- Micheli, L.; Mattoli, L.; Maidecchi, A.; Pacini, A.; Ghelardini, C.; Di Cesare Mannelli, L. Effect of Vitis vinifera hydroalcoholic extract against oxaliplatin neurotoxicity: In vitro and in vivo evidence. Sci. Rep. 2018, 8, 14364. [Google Scholar] [CrossRef] [PubMed]
- Kubyshkin, A.; Shevandova, A.; Petrenko, V.; Fomochkina, I.; Sorokina, L.; Kucherenko, A.; Gordienko, A.; Khimich, N.; Zyablitskaya, E.; Makalish, T.; et al. Anti-inflammatory and antidiabetic effects of grape-derived stilbene concentrate in the experimental metabolic syndrome. J. Diabetes Metab. Disord. 2020, 19, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Anatolii, K.; Yury, O.; Iryna, F.; Inna, C.; Georgiy, Z.; Yuliana, S. Polyphenols of Red Grape Wines and Alcohol-Free Food Concentrates in Rehabilitation Technologies. In Polyphenols; Janica, W., Ed.; IntechOpen: Rijeka, Yugoslavia, 2018; Chapter 5. [Google Scholar]
- Ben Khadher, T.; Aydi, S.; Mars, M.; Bouajila, J. Study on the Chemical Composition and the Biological Activities of Vitis vinifera Stem Extracts. Molecules 2022, 27, 3109. [Google Scholar] [CrossRef] [PubMed]
- Dopico-Garcia, M.S.; Fique, A.; Guerra, L.; Afonso, J.M.; Pereira, O.; Valentao, P.; Andrade, P.B.; Seabra, R.M. Principal components of phenolics to characterize red Vinho Verde grapes: Anthocyanins or non-coloured compounds? Talanta 2008, 75, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Novak, I.; Janeiro, P.; Seruga, M.; Oliveira-Brett, A.M. Ultrasound extracted flavonoids from four varieties of Portuguese red grape skins determined by reverse-phase high-performance liquid chromatography with electrochemical detection. Anal. Chim. Acta 2008, 630, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Spacil, Z.; Novakova, L.; Solich, P. Analysis of phenolic compounds by high performance liquid chromatography and ultra performance liquid chromatography. Talanta 2008, 76, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.D.; Luo, M.; Huang, S.Y.; Saimaiti, A.; Shang, A.; Gan, R.Y.; Li, H.B. Effects and Mechanisms of Resveratrol on Aging and Age-Related Diseases. Oxid. Med. Cell Longev. 2021, 2021, 9932218. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.Q.; Deng, G.F.; Guo, Y.J.; Li, H.B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef] [PubMed]
- Weidner, S.; Rybarczyk, A.; Karamac, M.; Krol, A.; Mostek, A.; Grebosz, J.; Amarowicz, R. Differences in the phenolic composition and antioxidant properties between Vitis coignetiae and Vitis vinifera seeds extracts. Molecules 2013, 18, 3410–3426. [Google Scholar] [CrossRef]
- Souza, E.L.; Nascimento, T.S.; Magalhaes, C.M.; Barreto, G.A.; Leal, I.L.; Dos Anjos, J.P.; Machado, B.A.S. Development and Characterization of Powdered Antioxidant Compounds Made from Shiraz (Vitis vinifera L.) Grape Peels and Arrowroot (Maranta arundinacea L.). Sci. World J. 2022, 2022, 7664321. [Google Scholar] [CrossRef]
- Weidner, S.; Kordala, E.; Brosowska-Arendt, W.; Karamac, M.; Kosinska, A.; Amarowicz, R. Phenolic compounds and properties of antioxidants in grapevine roots [Vitis vinifera L.] under low-temperature stress followed by recovery. Acta Soc. Bot. Pol. 2009, 78, 279–286. [Google Scholar] [CrossRef]
- Kapcsandi, V.; Hanczne Lakatos, E.; Sik, B.; Linka, L.A.; Szekelyhidi, R. Antioxidant and polyphenol content of different Vitis vinifera seed cultivars and two facilities of production of a functional bakery product. Chem. Zvesti 2021, 75, 5711–5717. [Google Scholar] [CrossRef] [PubMed]
- Recinella, L.; Chiavaroli, A.; Veschi, S.; Cama, A.; Acquaviva, A.; Libero, M.L.; Leone, S.; Di Simone, S.C.; Pagano, E.; Zengin, G.; et al. A grape (Vitis vinifera L.) pomace water extract modulates inflammatory and immune response in SW-480 cells and isolated mouse colon. Phytother. Res. 2022, 36, 4620–4630. [Google Scholar] [CrossRef]
- You, D.; Jeong, Y.; Yoon, S.Y.; Kim, S.A.; Lo, E.; Kim, S.W.; Lee, J.E.; Nam, S.J.; Kim, S. Entelon((R)) (Vitis vinifera Seed Extract) Prevents Cancer Metastasis via the Downregulation of Interleukin-1 Alpha in Triple-Negative Breast Cancer Cells. Molecules 2021, 26, 3644. [Google Scholar] [CrossRef]
- Sei-Ichi, K.; Toda, K.; Matsumoto, K.; Ishihara, C.; Nonobe, S.; Matsunaga, C.; Gomi, Y.K.; Senga, S.; Kawaguchi, K.; Yamamoto, A.; et al. Isolation and characterization of a novel oligomeric proanthocyanidin with significant anti-cancer activities from grape stems (Vitis vinifera). Sci. Rep. 2019, 9, 12046. [Google Scholar] [CrossRef]
- Lantzouraki, D.Z.; Tsiaka, T.; Soteriou, N.; Asimomiti, G.; Spanidi, E.; Natskoulis, P.; Gardikis, K.; Sinanoglou, V.J.; Zoumpoulakis, P. Antioxidant Profiles of Vitis vinifera L. and Salvia triloba L. Leaves Using High-Energy Extraction Methodologies. J. AOAC Int. 2020, 103, 413–421. [Google Scholar] [CrossRef]
- Cotoras, M.; Vivanco, H.; Melo, R.; Aguirre, M.; Silva, E.; Mendoza, L. In vitro and in vivo evaluation of the antioxidant and prooxidant activity of phenolic compounds obtained from grape (Vitis vinifera) pomace. Molecules 2014, 19, 21154–21167. [Google Scholar] [CrossRef]
- Hollands, W.J.; Voorspoels, S.; Jacobs, G.; Aaby, K.; Meisland, A.; Garcia-Villalba, R.; Tomas-Barberan, F.; Piskula, M.K.; Mawson, D.; Vovk, I.; et al. Development, validation and evaluation of an analytical method for the determination of monomeric and oligomeric procyanidins in apple extracts. J. Chromatogr. A 2017, 1495, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Minn, A.J.; Gupta, G.P.; Siegel, P.M.; Bos, P.D.; Shu, W.; Giri, D.D.; Viale, A.; Olshen, A.B.; Gerald, W.L.; Massague, J. Genes that mediate breast cancer metastasis to lung. Nature 2005, 436, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, M.; Turnbull, T.; Sebastian, S.; Kempson, I. The MTT Assay: Utility, Limitations, Pitfalls, and Interpretation in Bulk and Single-Cell Analysis. Int. J. Mol. Sci. 2021, 22, 12827. [Google Scholar] [CrossRef] [PubMed]
- Papageorgis, P.; Ozturk, S.; Lambert, A.W.; Neophytou, C.M.; Tzatsos, A.; Wong, C.K.; Thiagalingam, S.; Constantinou, A.I. Targeting IL13Ralpha2 activates STAT6-TP63 pathway to suppress breast cancer lung metastasis. Breast Cancer Res. 2015, 17, 98. [Google Scholar] [CrossRef] [PubMed]
- Savva, C.G.; Totokotsopoulos, S.; Nicolaou, K.C.; Neophytou, C.M.; Constantinou, A.I. Selective activation of TNFR1 and NF-kappaB inhibition by a novel biyouyanagin analogue promotes apoptosis in acute leukemia cells. BMC Cancer 2016, 16, 279. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B.; Darley-Usmar, V.; Davies, K.J.; Dennery, P.A.; Forman, H.J.; Grisham, M.B.; Mann, G.E.; Moore, K.; Roberts, L.J., 2nd; Ischiropoulos, H. Measuring reactive oxygen and nitrogen species with fluorescent probes: Challenges and limitations. Free Radic. Biol. Med. 2012, 52, 1–6. [Google Scholar] [CrossRef]
- Al-Warhi, T.; Zahran, E.M.; Selim, S.; Al-Sanea, M.M.; Ghoneim, M.M.; Maher, S.A.; Mostafa, Y.A.; Alsenani, F.; Elrehany, M.A.; Almuhayawi, M.S.; et al. Antioxidant and Wound Healing Potential of Vitis vinifera Seeds Supported by Phytochemical Characterization and Docking Studies. Antioxidants 2022, 11, 881. [Google Scholar] [CrossRef]
- Ma, Z.F.; Zhang, H. Phytochemical Constituents, Health Benefits, and Industrial Applications of Grape Seeds: A Mini-Review. Antioxidants 2017, 6, 71. [Google Scholar] [CrossRef] [PubMed]
- Somkuwar, R.G.; Bhange, M.A.; Oulkar, D.P.; Sharma, A.K.; Ahammed Shabeer, T.P. Estimation of polyphenols by using HPLC-DAD in red and white wine grape varieties grown under tropical conditions of India. J. Food Sci. Technol. 2018, 55, 4994–5002. [Google Scholar] [CrossRef] [PubMed]
- Bocsan, I.C.; Magureanu, D.C.; Pop, R.M.; Levai, A.M.; Macovei, S.O.; Patrasca, I.M.; Chedea, V.S.; Buzoianu, A.D. Antioxidant and Anti-Inflammatory Actions of Polyphenols from Red and White Grape Pomace in Ischemic Heart Diseases. Biomedicines 2022, 10, 2337. [Google Scholar] [CrossRef] [PubMed]
- Trost, K.; Klancnik, A.; Mozetic Vodopivec, B.; Sternad Lemut, M.; Jug Novsak, K.; Raspor, P.; Smole Mozina, S. Polyphenol, antioxidant and antimicrobial potential of six different white and red wine grape processing leftovers. J. Sci. Food Agric. 2016, 96, 4809–4820. [Google Scholar] [CrossRef] [PubMed]
- Serino, A.; Salazar, G. Protective Role of Polyphenols against Vascular Inflammation, Aging and Cardiovascular Disease. Nutrients 2018, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, L.; Giampieri, F.; Alvarez Suarez, J.M.; Gasparrini, M.; Mezzetti, B.; Forbes Hernandez, T.Y.; Battino, M.A. Isolation of strawberry anthocyanin-rich fractions and their mechanisms of action against murine breast cancer cell lines. Food Funct. 2019, 10, 7103–7120. [Google Scholar] [CrossRef] [PubMed]
- Tenore, G.C.; Manfra, M.; Stiuso, P.; Coppola, L.; Russo, M.; Gomez Monterrey, I.M.; Campiglia, P. Antioxidant profile and in vitro cardiac radical-scavenging versus pro-oxidant effects of commercial red grape juices (Vitis vinifera L. cv. Aglianico N.). J. Agric. Food Chem. 2012, 60, 9680–9687. [Google Scholar] [CrossRef]
- Pepe, G.; Salviati, E.; Rapa, S.F.; Ostacolo, C.; Cascioferro, S.; Manfra, M.; Autore, G.; Marzocco, S.; Campiglia, P. Citrus sinensis and Vitis vinifera Protect Cardiomyocytes from Doxorubicin-Induced Oxidative Stress: Evaluation of Onconutraceutical Potential of Vegetable Smoothies. Antioxidants 2020, 9, 378. [Google Scholar] [CrossRef]
- Gregoriou, G.; Neophytou, C.M.; Vasincu, A.; Gregoriou, Y.; Hadjipakkou, H.; Pinakoulaki, E.; Christodoulou, M.C.; Ioannou, G.D.; Stavrou, I.J.; Christou, A.; et al. Anti-Cancer Activity and Phenolic Content of Extracts Derived from Cypriot Carob (Ceratonia siliqua L.) Pods Using Different Solvents. Molecules 2021, 26, 5017. [Google Scholar] [CrossRef]
- Velez, M.D.; Llano-Ramirez, M.A.; Ramon, C.; Rojas, J.; Bedoya, C.; Arango-Varela, S.; Santa-Gonzalez, G.A.; Gil, M. Antioxidant capacity and cytotoxic effect of an optimized extract of isabella grape (Vitis labrusca) on breast cancer cells. Heliyon 2023, 9, e16540. [Google Scholar] [CrossRef]
- Costa, V.M.; Carvalho, F.; Duarte, J.A.; Bastos Mde, L.; Remiao, F. The heart as a target for xenobiotic toxicity: The cardiac susceptibility to oxidative stress. Chem. Res. Toxicol. 2013, 26, 1285–1311. [Google Scholar] [CrossRef]
- Giovannelli, L.; Innocenti, M.; Santamaria, A.R.; Bigagli, E.; Pasqua, G.; Mulinacci, N. Antitumoural activity of viniferin-enriched extracts from Vitis vinifera L. cell cultures. Nat. Prod. Res. 2014, 28, 2006–2016. [Google Scholar] [CrossRef] [PubMed]
- Pitta, C.A.; Papageorgis, P.; Charalambous, C.; Constantinou, A.I. Reversal of ER-beta silencing by chromatin modifying agents overrides acquired tamoxifen resistance. Cancer Lett. 2013, 337, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Toth, B.; Saadat, G.; Geller, A.; Scholz, C.; Schulze, S.; Friese, K.; Jeschke, U. Human umbilical vascular endothelial cells express estrogen receptor beta (ERbeta) and progesterone receptor A (PR-A), but not ERalpha and PR-B. Histochem. Cell Biol. 2008, 130, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Comsa, S.; Cimpean, A.M.; Raica, M. The Story of MCF-7 Breast Cancer Cell Line: 40 years of Experience in Research. Anticancer. Res. 2015, 35, 3147–3154. [Google Scholar] [PubMed]
- Yusuf, R.; Frenkel, K. Morphologic transformation of human breast epithelial cells MCF-10A: Dependence on an oxidative microenvironment and estrogen/epidermal growth factor receptors. Cancer Cell Int. 2010, 10, 30. [Google Scholar] [CrossRef]
- Cipolletti, M.; Solar Fernandez, V.; Montalesi, E.; Marino, M.; Fiocchetti, M. Beyond the Antioxidant Activity of Dietary Polyphenols in Cancer: The Modulation of Estrogen Receptors (ERs) Signaling. Int. J. Mol. Sci. 2018, 19, 2624. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic Galovic, A.; Jovanovic Ljeskovic, N.; Vidovic, S.; Vladic, J.; Jojic, N.; Ilic, M.; Srdic Rajic, T.; Kojic, V.; Jakimov, D. The Effects of Resveratrol-Rich Extracts of Vitis vinifera Pruning Waste on HeLa, MCF-7 and MRC-5 Cells: Apoptosis, Autophagia and Necrosis Interplay. Pharmaceutics 2022, 14, 2017. [Google Scholar] [CrossRef]
- Leone, A.; Longo, C.; Gerardi, C.; Trosko, J.E. Pro-Apoptotic Effect of Grape Seed Extract on MCF-7 Involves Transient Increase of Gap Junction Intercellular Communication and Cx43 Up-Regulation: A Mechanism of Chemoprevention. Int. J. Mol. Sci. 2019, 20, 3244. [Google Scholar] [CrossRef]
- Grace Nirmala, J.; Evangeline Celsia, S.; Swaminathan, A.; Narendhirakannan, R.T.; Chatterjee, S. Cytotoxicity and apoptotic cell death induced by Vitis vinifera peel and seed extracts in A431 skin cancer cells. Cytotechnology 2018, 70, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, V.; Singh, T.; Katiyar, S.K. Multi-targeted prevention and therapy of cancer by proanthocyanidins. Cancer Lett. 2008, 269, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Mantena, S.K.; Baliga, M.S.; Katiyar, S.K. Grape seed proanthocyanidins induce apoptosis and inhibit metastasis of highly metastatic breast carcinoma cells. Carcinogenesis 2006, 27, 1682–1691. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Perez, F.; Belchi-Navarro, S.; Almagro, L.; Bru, R.; Pedreno, M.A.; Gomez-Ros, L.V. Cytotoxic effect of natural trans-resveratrol obtained from elicited Vitis vinifera cell cultures on three cancer cell lines. Plant Foods Hum. Nutr. 2012, 67, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Kiyozuka, Y.; Uemura, Y.; Senzaki, H.; Shikata, N.; Hioki, K.; Tsubura, A. Resveratrol inhibits human breast cancer cell growth and may mitigate the effect of linoleic acid, a potent breast cancer cell stimulator. J. Cancer Res. Clin. Oncol. 2001, 127, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, W.; Zhan, J. Grape Seed Proanthocyanidins Induce Autophagy and Modulate Survivin in HepG2 Cells and Inhibit Xenograft Tumor Growth in Vivo. Nutrients 2019, 11, 2983. [Google Scholar] [CrossRef]
- Martin, L.A.; Dowsett, M. BCL-2: A new therapeutic target in estrogen receptor-positive breast cancer? Cancer Cell 2013, 24, 7–9. [Google Scholar] [CrossRef]
- Nabilsi, N.H.; Broaddus, R.R.; McCampbell, A.S.; Lu, K.H.; Lynch, H.T.; Chen, L.M.; Loose, D.S. Sex hormone regulation of survivin gene expression. J. Endocrinol. 2010, 207, 237–243. [Google Scholar] [CrossRef]
- Pike, C.J. Estrogen modulates neuronal Bcl-xL expression and beta-amyloid-induced apoptosis: Relevance to Alzheimer’s disease. J. Neurochem. 1999, 72, 1552–1563. [Google Scholar] [CrossRef]
- Nirmala, J.G.; Narendhirakannan, R.T. Vitis vinifera peel and seed gold nanoparticles exhibit chemopreventive potential, antioxidant activity and induce apoptosis through mutant p53, Bcl-2 and pan cytokeratin down-regulation in experimental animals. Biomed. Pharmacother. 2017, 89, 902–917. [Google Scholar] [CrossRef] [PubMed]
- Balcer-Kubiczek, E.K.; Yin, J.; Lin, K.; Harrison, G.H.; Abraham, J.M.; Meltzer, S.J. p53 mutational status and survival of human breast cancer MCF-7 cell variants after exposure to X rays or fission neutrons. Radiat. Res. 1995, 142, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Salman, G.; Pehlivanoglu, S.; Aydin Acar, C.; Yesilot, S. Anticancer Effects of Vitis vinifera L. Mediated Biosynthesized Silver Nanoparticles and Cotreatment with 5 Fluorouracil on HT-29 Cell Line. Biol. Trace Elem. Res. 2022, 200, 3159–3170. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Description | OPC DP 1–8 g/100 g | Catechins DP 1 g/100 g | OPC DP 2–4 g/100 g | OPC DP 5–8 g/100 g | TPC as Pyrogallol Solvent H2O g/100 g | TPC as Catechin Solvent H2O g/100 g |
|---|---|---|---|---|---|---|---|
| E1 | Whole seeds dry-extract, extraction solvent EtOH 70%m/m | 19.79 | 6.72 | 6.50 | 6.57 | 28.89 | 38.92 |
| E2 | Whole seeds dry-extract, extraction solvent H2O | 15.48 | 4.74 | 5.84 | 4.91 | 23.53 | 31.74 |
| E3 | Crushed seeds without oil extract dried precipitate extraction solvent EtOH 62%m/m | 9.89 | 2.47 | 3.56 | 3.86 | 12.44 | 17.53 |
| E4 | Crushed seeds without oil dry-extract after precipitation, dried liquid extraction solvent EtOH 62%m/m | 22.37 | 4.89 | 7.81 | 9.68 | 25.94 | 34.33 |
| E5 | Whole seeds, organic seeds from white grape dry-extract extraction solvent EtOH 62%m/m | 15.14 | 3.28 | 5.84 | 6.01 | 20.44 | 27.53 |
| E6 | Whole seeds, pre-washed, organic seeds, from white grape dried wash-water | 7.25 | 1.99 | 2.92 | 2.35 | 8.36 | 11.25 |
| E7 | Whole seeds, pre-washed, organic seeds from white grape dry-extract extraction solvent EtOH 62%m/m | 16.20 | 3.11 | 6.13 | 6.97 | 25.00 | 33.02 |
| E8 | Fruits without seeds dry-extract extraction solvent EtOH 62%m/m | 0.44 | 0.38 | 0.06 | <0.005 | 3.82 | 5.50 |
| E9 | Fruits without seeds prewashed, dried wash-water | 0.13 | 0.13 | <0.005 | <0.005 | 1.47 | 2.39 |
| E10 | Fruits without seeds pre-washed dry-extract extraction solvent EtOH 62%m/m | 0.93 | 0.55 | 0.31 | 0.08 | 6.90 | 9.33 |
| E11 | Leaves dry-extract extraction solvent EtOH 62%m/m | 0.49 | 0.47 | 0.01 | <0.005 | 2.97 | 4.74 |
| E12 | Wood dry-extract extraction solvent EtOH 62%m/m | 0.84 | 0.77 | 0.07 | 0.01 | 0.86 | 1.66 |
| Extract | Slope + | CAA50 * (mg/L) |
|---|---|---|
| E1 | −0.0100 ± 0.007 | 31.00 |
| E2 | −0.0080 ± 0.007 | 38.94 |
| E3 | Not tested ** | Not tested ** |
| E4 | −0.0043 ± 0.006 | 36.24 |
| E5 | Not tested ** | Not tested ** |
| E6 | 0.0053 ± 0.006 | Not evaluable *** |
| E7 | −0.0271 ± 0.007 | 53.82 |
| E8 | −0.0326 ± 0.012 | 57.82 |
| E9 | 0.0185 ± 0.008 | Not evaluable *** |
| E10 | 0.0040 ± 0.006 | Not evaluable *** |
| E11 | −0.0256 ± 0.008 | 54.46 |
| E12 | −0.0335 ± 0.020 | 66.33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsantila, E.M.; Esslinger, N.; Christou, M.; Papageorgis, P.; Neophytou, C.M. Antioxidant and Anticancer Activity of Vitis vinifera Extracts in Breast Cell Lines. Life 2024, 14, 228. https://doi.org/10.3390/life14020228
Tsantila EM, Esslinger N, Christou M, Papageorgis P, Neophytou CM. Antioxidant and Anticancer Activity of Vitis vinifera Extracts in Breast Cell Lines. Life. 2024; 14(2):228. https://doi.org/10.3390/life14020228
Chicago/Turabian StyleTsantila, Evgenia Maria, Nils Esslinger, Maria Christou, Panagiotis Papageorgis, and Christiana M. Neophytou. 2024. "Antioxidant and Anticancer Activity of Vitis vinifera Extracts in Breast Cell Lines" Life 14, no. 2: 228. https://doi.org/10.3390/life14020228
APA StyleTsantila, E. M., Esslinger, N., Christou, M., Papageorgis, P., & Neophytou, C. M. (2024). Antioxidant and Anticancer Activity of Vitis vinifera Extracts in Breast Cell Lines. Life, 14(2), 228. https://doi.org/10.3390/life14020228