
Role of IL-4 and IL-13 in Cutaneous T Cell Lymphoma

 , , , and
, , , and 
Abstract
1. Introduction: Cutaneous T Cell Lymphoma
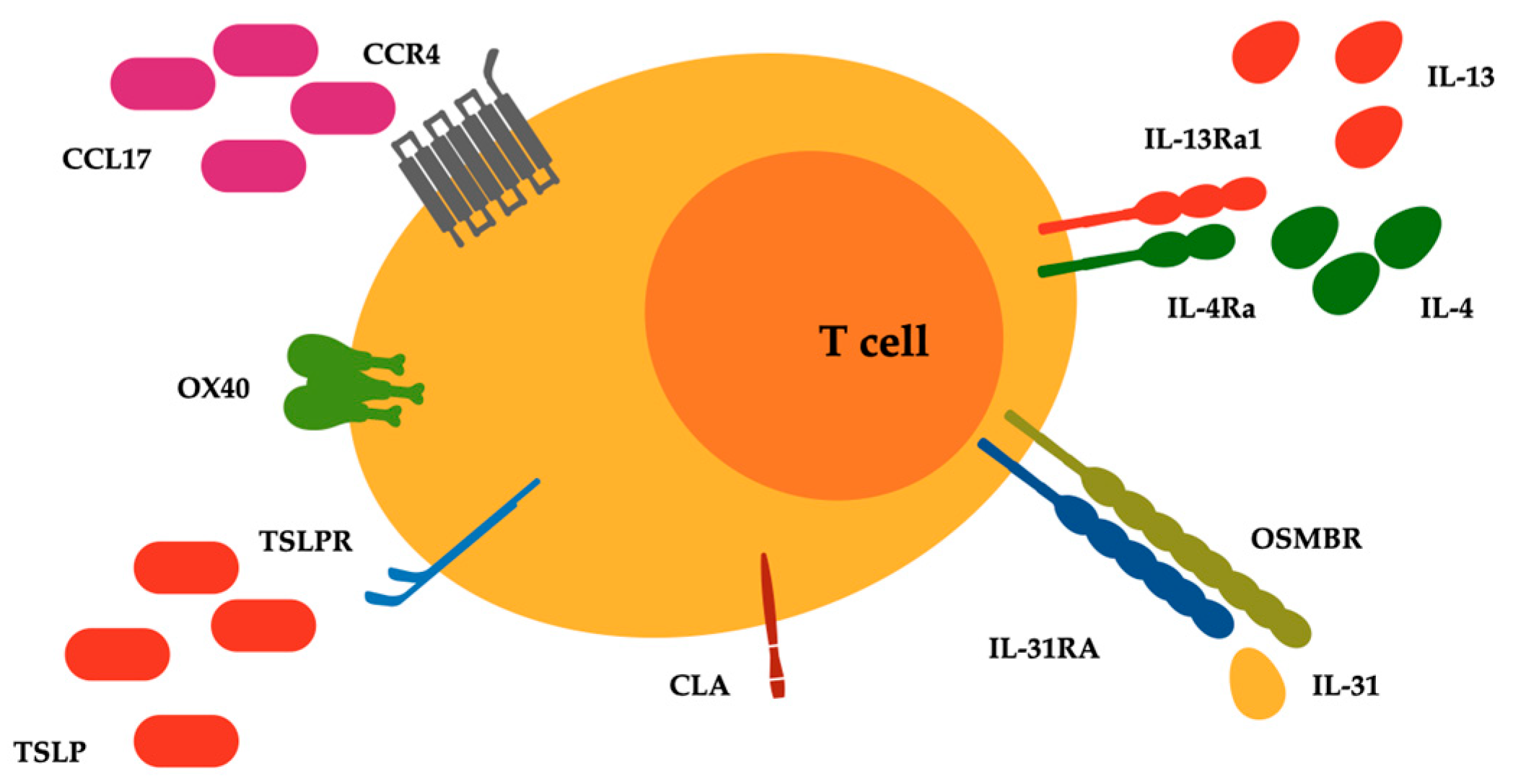
2. IL-4 and IL-13 in the Development and Progression of Cutaneous T Cell Lymphoma
3. Targeting IL-4 and IL-13 in Cutaneous T Cell Lymphoma
4. Targeting IL-13 in Cutaneous T Cell Lymphoma
5. Targeting IL-31 in Cutaneous T Cell Lymphoma
6. Targeting Thymic Stromal Lymphopoietin in Cutaneous T Cell Lymphoma
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gaydosik, A.M.; Queen, D.S.; Trager, M.H.; Akilov, O.E.; Geskin, L.J.; Fuschiotti, P. Genome-Wide Transcriptome Analysis of the STAT6-Regulated Genes in Advanced-Stage Cutaneous T-Cell Lymphoma. Blood 2020, 136, 1748–1759. [Google Scholar] [CrossRef] [PubMed]
- Dobos, G.; Pohrt, A.; Ram-Wolff, C.; Lebbé, C.; Bouaziz, J.-D.; Battistella, M.; Bagot, M.; De Masson, A. Epidemiology of Cutaneous T-Cell Lymphomas: A Systematic Review and Meta-Analysis of 16,953 Patients. Cancers 2020, 12, 2921. [Google Scholar] [CrossRef] [PubMed]
- Willemze, R.; Cerroni, L.; Kempf, W.; Berti, E.; Facchetti, F.; Swerdlow, S.H.; Jaffe, E.S. The 2018 Update of the WHO-EORTC Classification for Primary Cutaneous Lymphomas. Blood 2019, 133, 1703–1714. [Google Scholar] [CrossRef] [PubMed]
- Jawed, S.I.; Myskowski, P.L.; Horwitz, S.; Moskowitz, A.; Querfeld, C. Primary Cutaneous T-Cell Lymphoma (Mycosis Fungoides and Sézary Syndrome). J. Am. Acad. Dermatol. 2014, 70, 205.e1–205.e16. [Google Scholar] [CrossRef] [PubMed]
- Ottevanger, R.; Feenstra, J.S.; van Vliet, L.M.; van Beugen, S.; Evers, A.W.M.; Kennedy, C.; Willemze, R.; Vermeer, M.H.; Quint, K.D. Unveiling the Hidden Struggles: Exploring the Profound Impact of Advanced Stage Cutaneous T-Cell Lymphoma on Quality of Life. Ski. Health Dis. 2023, 3, e300. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.J.; Clark, R.A.; Watanabe, R.; Kupper, T.S. Sézary Syndrome and Mycosis Fungoides Arise from Distinct T-Cell Subsets: A Biologic Rationale for Their Distinct Clinical Behaviors. Blood 2010, 116, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Rübben, A.; Kempf, W.; Kadin, M.E.; Zimmermann, D.R.; Burg, G. Multilineage Progression of Genetically Unstable Tumor Subclones in Cutaneous T-cell Lymphoma. Exp. Dermatol. 2004, 13, 472–483. [Google Scholar] [CrossRef] [PubMed]
- Saulite, I.; Hoetzenecker, W.; Weidinger, S.; Cozzio, A.; Guenova, E.; Wehkamp, U. Sézary Syndrome and Atopic Dermatitis: Comparison of Immunological Aspects and Targets. BioMed Res. Int. 2016, 2016, 9717530. [Google Scholar] [CrossRef]
- Krejsgaard, T.; Lindahl, L.M.; Mongan, N.P.; Wasik, M.A.; Litvinov, I.V.; Iversen, L.; Langhoff, E.; Woetmann, A.; Odum, N. Malignant Inflammation in Cutaneous T-cell Lymphoma—A Hostile Takeover. Semin. Immunopathol. 2017, 39, 269–282. [Google Scholar] [CrossRef]
- Chung, C.G.; Poligone, B. Cutaneous T cell Lymphoma: An Update on Pathogenesis and Systemic Therapy. Curr. Hematol. Malig. Rep. 2015, 10, 468–476. [Google Scholar] [CrossRef]
- Vieyra-Garcia, P.; Crouch, J.D.; O’Malley, J.T.; Seger, E.W.; Yang, C.H.; Teague, J.E.; Vromans, A.M.; Gehad, A.; Win, T.S.; Yu, Z.; et al. Benign T Cells Drive Clinical Skin Inflammation in Cutaneous T Cell Lymphoma. JCI Insight 2019, 4, e124233. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, B.R.; Zain, J.; Rosen, S.T.; Querfeld, C. Tumor Microenvironment in Mycosis Fungoides and Sézary Syndrome. Curr. Opin. Oncol. 2016, 28, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Lebas, E.; Collins, P.; Somja, J.; Nikkels, A.F. Causes of Death in Cutaneous T-Cell Lymphoma Patients. Dermatology 2023, 239, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wu, X.; Hwang, S.T.; Liu, J. The Role of Tumor Microenvironment in Mycosis Fungoides and Sézary Syndrome. Ann. Dermatol. 2021, 33, 487. [Google Scholar] [CrossRef] [PubMed]
- Pals, S.T.; De Gorter, D.J.J.; Spaargaren, M. Lymphoma Dissemination: The Other Face of Lymphocyte Homing. Blood 2007, 110, 3102–3111. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Shin, D.B.; Syed, M.N.; Abuabara, K.; Lemeshow, A.R.; Fuxench, Z.C.C.; Gelfand, J.M. Malignancy Risk in Patients with Atopic Dermatitis: A Population-Based Cohort Study. Br. J. Dermatol. 2023, 189, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Josephs, D.H.; Spicer, J.F.; Corrigan, C.J.; Gould, H.J.; Karagiannis, S.N. Epidemiological Associations of Allergy, IgE and Cancer. Clin. Exp. Allergy 2013, 43, 1110–1123. [Google Scholar] [CrossRef]
- Patil, K.; Kuttikrishnan, S.; Khan, A.Q.; Ahmad, F.; Alam, M.; Buddenkotte, J.; Ahmad, A.; Steinhoff, M.; Uddin, S. Molecular Pathogenesis of Cutaneous T Cell Lymphoma: Role of Chemokines, Cytokines, and Dysregulated Signaling Pathways. Semin. Cancer Biol. 2022, 86, 382–399. [Google Scholar] [CrossRef]
- Pérez, C.; Mondéjar, R.; García-Díaz, N.; Cereceda, L.; León, A.; Montes, S.; Durán Vian, C.; Pérez Paredes, M.G.; González-Morán, A.; Miguel, V.; et al. Advanced-stage Mycosis Fungoides: Role of the Signal Transducer and Activator of Transcription 3, Nuclear factor-κB and Nuclear Factor of Activated T Cells Pathways. Br. J. Dermatol. 2019, 182, 147–155. [Google Scholar] [CrossRef]
- Dulmage, B.O.; Geskin, L.J. Lessons Learned from Gene Expression Profiling of Cutaneous T-Cell Lymphoma. Br. J. Dermatol. 2013, 169, 1188–1197. [Google Scholar] [CrossRef]
- Netchiporouk, E.; Litvinov, I.V.; Moreau, L.; Gilbert, M.; Sasseville, D.; Duvic, M. Deregulation in STAT Signaling Is Important for Cutaneous T-Cell Lymphoma (CTCL) Pathogenesis and Cancer Progression. Cell Cycle 2014, 13, 3331–3335. [Google Scholar] [CrossRef] [PubMed]
- Karpathiou, G.; Papoudou-Bai, A.; Ferrand, E.; Dumollard, J.M.; Peoc’h, M. STAT6: A Review of a Signaling Pathway Implicated in Various Diseases with a Special Emphasis in Its Usefulness in Pathology. Pathol.-Res. Pract. 2021, 223, 153477. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, Y. Thymus and Activation-regulated Chemokine as a Clinical Biomarker in Atopic Dermatitis. J. Dermatol. 2014, 41, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Kadono, T. New Aspects of the Clinicopathological Features and Treatment of Mycosis Fungoides and Sézary Syndrome. J. Dermatol. 2015, 42, 941–944. [Google Scholar] [CrossRef] [PubMed]
- Perera, L.P.; Zhang, M.; Nakagawa, M.; Petrus, M.N.; Maeda, M.; Kadin, M.E.; Waldmann, T.A.; Perera, P. Chimeric Antigen Receptor Modified T Cells That Target Chemokine Receptor CCR4 as a Therapeutic Modality for T-cell Malignancies. Am. J. Hematol. 2017, 92, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K. New Therapies and Immunological Findings in Cutaneous T-Cell Lymphoma. Front. Oncol. 2018, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Sugaya, M.; Suga, H.; Oka, T.; Kawaguchi, M.; Miyagaki, T.; Fujita, H.; Sato, S. Thymic Stromal Chemokine TSLP Acts through Th2 Cytokine Production to Induce Cutaneous T-Cell Lymphoma. Cancer Res. 2016, 76, 6241–6252. [Google Scholar] [CrossRef] [PubMed]
- Urosevic, M.; Willers, J.; Mueller, B.; Kempf, W.; Burg, G.; Dummer, R. HLA-G Protein up-Regulation in Primary Cutaneous Lymphomas Is Associated with Interleukin-10 Expression in Large Cell T-Cell Lymphomas and Indolent B-Cell Lymphomas. Blood 2002, 99, 609–617. [Google Scholar] [CrossRef]
- Scala, E.; Abeni, D.; Palazzo, P.; Liso, M.; Pomponi, D.; Lombardo, G.; Picchio, M.C.; Narducci, M.G.; Russo, G.; Mari, A. Specific IgE toward Allergenic Molecules Is a New Prognostic Marker in Patients with Sézary Syndrome. Int. Arch. Allergy Immunol. 2012, 157, 159–167. [Google Scholar] [CrossRef]
- Wolk, K.; Mitsui, H.; Witte, K.; Gellrich, S.; Gulati, N.; Humme, D.; Witte, E.; Gonsior, M.; Beyer, M.; Kadin, M.E.; et al. Deficient Cutaneous Antibacterial Competence in Cutaneous T-Cell Lymphomas: Role of Th2-Mediated Biased Th17 Function. Clin. Cancer Res. 2014, 20, 5507–5516. [Google Scholar] [CrossRef]
- Ferretti, E.; Corcione, A.; Pistoia, V. The IL-31/IL-31 Receptor Axis: General Features and Role in Tumor Microenvironment. J. Leukoc. Biol. 2017, 102, 711–717. [Google Scholar] [CrossRef]
- Ohmatsu, H.; Sugaya, M.; Suga, H.; Morimura, S.; Miyagaki, T.; Kai, H.; Kagami, S.; Fujita, H.; Asano, Y.; Tada, Y.; et al. Serum IL-31 Levels Are Increased in Patients with Cutaneous T-Cell Lymphoma. Acta Derm. Venerol. 2012, 92, 282–283. [Google Scholar] [CrossRef] [PubMed]
- Belmesk, L.; Muntyanu, A.; Cantin, E.; AlHalees, Z.; Jack, C.S.; Le, M.; Sasseville, D.; Iannattone, L.; Ben-Shoshan, M.; Litvinov, I.V.; et al. Prominent Role of Type 2 Immunity in Skin Diseases: Beyond Atopic Dermatitis. J. Cutan. Med. Surg. 2022, 26, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J. Immunopathogenesis and Therapy of Cutaneous T Cell Lymphoma. J. Clin. Investig. 2005, 115, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Renzo, M.D. Extracorporeal Photochemotherapy Restores Thl/Th2 Imbalance in Patients with Early Stage Cutaneous T-Cell Lymphoma. Immunology 1997, 92, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Lazaridou, I.; Ram-Wolff, C.; Bouaziz, J.; Bégon, E.; Battistella, M.; Rivet, J.; Jachiet, M.; Bagot, M.; Masson, A. Dupilumab Treatment in Two Patients with Cutaneous T-Cell Lymphomas. Acta Derm. Venereol. 2020, 100, adv00271. [Google Scholar] [CrossRef] [PubMed]
- Talmon, A.; Elias, S.; Rubin, L.; Ribak, Y.; Ben Dori, E.; Shamriz, O.; Lotem, M.; Adini, I.; Tal, Y. Dupilumab for Cancer-Associated Refractory Pruritus. J. Allergy Clin. Immunol. Glob. 2023, 2, 100128. [Google Scholar] [CrossRef] [PubMed]
- Steck, O.; Bertschi, N.L.; Luther, F.; van den Berg, J.; Winkel, D.J.; Holbro, A.; Schlapbach, C. Rapid and Sustained Control of Itch and Reduction in Th2 Bias by Dupilumab in a Patient with Sézary Syndrome. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- Kychygina, A.; Cassagne, M.; Tauber, M.; Galiacy, S.; Paul, C.; Fournié, P.; Simon, M. Dupilumab-Associated Adverse Events During Treatment of Allergic Diseases. Clin. Rev. Allergy Immunol. 2022, 62, 519–533. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Di Nardo, L.; Gori, N.; Antonelli, F.; Pinto, L.; Cuffaro, G.; Piro, G.; Savino, G.; Tortora, G.; Peris, K. Dupilumab-Associated Ocular Adverse Events Are Predicted by Low Tear Break-up Time and Correlate with High IL-33 Tear Concentrations in Patients with Atopic Dermatitis. Exp. Dermatol. 2023, 32, 1531–1537. [Google Scholar] [CrossRef]
- Tran, J.; Morris, L.; Vu, A.; Duvic, M. Development of Sézary Syndrome Following the Administration of Dupilumab. Dermatol. Online J. 2020, 26, 17. [Google Scholar] [CrossRef]
- Jfri, A.; Smith, J.S.; Larocca, C. Diagnosis of Mycosis Fungoides or Sézary Syndrome after Dupilumab Use: A Systematic Review. J. Am. Acad. Dermatol. 2023, 88, 1164–1166. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, M.L.; Nguyen, M.T.; Aguirre, A.S.; Martinez-Escala, M.E.; Kim, J.; Walker, C.J.; Pontes, D.S.; Silverberg, J.I.; Choi, J.; Pro, B.; et al. Progression of Cutaneous T-Cell Lymphoma after Dupilumab: Case Review of 7 Patients. J. Am. Acad. Dermatol. 2020, 83, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Hamp, A.; Hanson, J.; Schwartz, R.A.; Lambert, W.C.; Alhatem, A. Dupilumab-Associated Mycosis Fungoides: A Cross-Sectional Study. Arch. Dermatol. Res. 2023, 315, 2561–2569. [Google Scholar] [CrossRef] [PubMed]
- Nakazaki, K.; Yoshida, M.; Masamoto, Y.; Shinozaki-Ushiku, A.; Ikemura, M.; Hisamoto, T.; Yasunaga, M.; Sato, S.; Kurokawa, M. Discordant Lymphomas of Classic Hodgkin Lymphoma and Peripheral T-Cell Lymphoma Following Dupilumab Treatment for Atopic Dermatitis. Int. J. Hematol. 2022, 116, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Andrews, A.-L.; Holloway, J.W.; Puddicombe, S.M.; Holgate, S.T.; Davies, D.E. Kinetic Analysis of the Interleukin-13 Receptor Complex. J. Biol. Chem. 2002, 277, 46073–46078. [Google Scholar] [CrossRef] [PubMed]
- Skinnider, B.F.; Elia, A.J.; Gascoyne, R.D.; Trümper, L.H.; von Bonin, F.; Kapp, U.; Patterson, B.; Snow, B.E.; Mak, T.W. Interleukin 13 and Interleukin 13 Receptor Are Frequently Expressed by Hodgkin and Reed-Sternberg Cells of Hodgkin Lymphoma. Blood 2001, 97, 250–255. [Google Scholar] [CrossRef]
- Chaouchi, N.; Wallon, C.; Goujard, C.; Tertian, G.; Rudent, A.; Caput, D.; Ferrera, P.; Minty, A.; Vazquez, A.; Delfraissy, J.F. Interleukin-13 Inhibits Interleukin-2-Induced Proliferation and Protects Chronic Lymphocytic Leukemia B Cells from in Vitro Apoptosis. Blood 1996, 87, 1022–1029. [Google Scholar] [CrossRef]
- Kelly-Welch, A.E.; Hanson, E.M.; Boothby, M.R.; Keegan, A.D. Interleukin-4 and Interleukin-13 Signaling Connections Maps. Science 2003, 300, 1527–1528. [Google Scholar] [CrossRef]
- Terabe, M.; Park, J.M.; Berzofsky, J.A. Role of IL-13 in Regulation of Anti-Tumor Immunity and Tumor Growth. Cancer Immunol. Immunother. 2004, 53, 79–85. [Google Scholar] [CrossRef]
- Kioi, M.; Shimamura, T.; Nakashima, H.; Hirota, M.; Tohnai, I.; Husain, S.R.; Puri, R.K. IL-13 Cytotoxin Has Potent Antitumor Activity and Synergizes with Paclitaxel in a Mouse Model of Oral Squamous Cell Carcinoma. Int. J. Cancer 2009, 124, 1440–1448. [Google Scholar] [CrossRef]
- Wills-Karp, M.; Chiaramonte, M. Interleukin-13 in Asthma. Curr. Opin. Pulm. Med. 2003, 9, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Geskin, L.J.; Viragova, S.; Stolz, D.B.; Fuschiotti, P. Interleukin-13 Is Overexpressed in Cutaneous T-Cell Lymphoma Cells and Regulates Their Proliferation. Blood 2015, 125, 2798–2805. [Google Scholar] [CrossRef] [PubMed]
- Sernicola, A.; Russo, I.; Alaibac, M. Small-Molecule-Based Immunotherapy for Immunologically Mediated Skin Conditions. Immunotherapy 2020, 12, 417–429. [Google Scholar] [CrossRef]
- Guenova, E.; Watanabe, R.; Teague, J.E.; Desimone, J.A.; Jiang, Y.; Dowlatshahi, M.; Schlapbach, C.; Schaekel, K.; Rook, A.H.; Tawa, M.; et al. TH2 Cytokines from Malignant Cells Suppress TH1 Responses and Enforce a Global TH2 Bias in Leukemic Cutaneous T-Cell Lymphoma. Clin. Cancer Res. 2013, 19, 3755–3763. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. IL-13 Effector Functions. Annu. Rev. Immunol. 2003, 21, 425–456. [Google Scholar] [CrossRef] [PubMed]
- Tollenaere, M.A.X.; Mølck, C.; Henderson, I.; Pollack, S.; Addis, P.; Petersen, H.H.; Norsgaard, H. Tralokinumab Effectively Disrupts the IL-13/IL-13Rα1/IL-4Rα Signaling Complex but Not the IL-13/IL-13Rα2 Complex. JID Innov. 2023, 3, 100214. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, K.; Irie, H. Interleukin-31 as a Clinical Target for Pruritus Treatment. Front. Med. 2021, 8, 638325. [Google Scholar] [CrossRef] [PubMed]
- Miake, S.; Tsuji, G.; Takemura, M.; Hashimoto-Hachiya, A.; Vu, Y.H.; Furue, M.; Nakahara, T. IL-4 Augments IL-31/IL-31 Receptor Alpha Interaction Leading to Enhanced Ccl 17 and Ccl 22 Production in Dendritic Cells: Implications for Atopic Dermatitis. Int. J. Mol. Sci. 2019, 20, 4053. [Google Scholar] [CrossRef]
- Kashem, S.W.; Fassett, M.S. Can Serum Biomarkers for Prurigo Nodularis Expose Pathophysiology or Just Treatment Response? JAMA Dermatol. 2023, 159, 915–917. [Google Scholar] [CrossRef]
- Datsi, A.; Steinhoff, M.; Ahmad, F.; Alam, M.; Buddenkotte, J. Interleukin-31: The “Itchy” Cytokine in Inflammation and Therapy. Allergy 2021, 76, 2982–2997. [Google Scholar] [CrossRef]
- Cayrol, C.; Girard, J.-P. Interleukin-33 (IL-33): A Nuclear Cytokine from the IL-1 Family. Immunol. Rev. 2018, 281, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Rook, A.H.; Rook, K.A.; Lewis, D.J. Interleukin-31, a Potent Pruritus-Inducing Cytokine and Its Role in Inflammatory Disease and in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2021, 1290, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Olszewska, B.; Sokołowska-Wojdyło, M.; Lakomy, J.; Nowicki, R.J. The Ambiguous Pruritogenic Role of Interleukin-31 in Cutaneous T-Cell Lymphomas in Comparison to Atopic Dermatitis: A Review. Postep. Dermatol. Alergol. 2020, 37, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Cedeno-Laurent, F.; Singer, E.M.; Wysocka, M.; Benoit, B.M.; Vittorio, C.C.; Kim, E.J.; Yosipovitch, G.; Rook, A.H. Improved Pruritus Correlates with Lower Levels of IL-31 in CTCL Patients under Different Therapeutic Modalities. Clin. Immunol. 2015, 158, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nattkemper, L.A.; Martinez-Escala, M.-E.; Gelman, A.B.; Singer, E.M.; Rook, A.H.; Guitart, J.; Yosipovitch, G. Cutaneous T-Cell Lymphoma and Pruritus: The Expression of IL-31 and Its Receptors in the Skin. Acta Derm. Venereol. 2016, 96, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.J.; Huang, S.; Duvic, M. Inflammatory Cytokines and Peripheral Mediators in the Pathophysiology of Pruritus in Cutaneous T-Cell Lymphoma. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1652–1656. [Google Scholar] [CrossRef] [PubMed]
- Feld, M.; Shpacovitch, V.M.; Fastrich, M.; Cevikbas, F.; Steinhoff, M. Interferon-γ Induces Upregulation and Activation of the Interleukin-31 Receptor in Human Dermal Microvascular Endothelial Cells. Exp. Dermatol. 2010, 19, 921–923. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.J. Nemolizumab: First Approval. Drugs 2022, 82, 1143–1150. [Google Scholar] [CrossRef]
- Kwatra, S.G.; Yosipovitch, G.; Legat, F.J.; Reich, A.; Paul, C.; Simon, D.; Naldi, L.; Lynde, C.; De Bruin-Weller, M.S.; Nahm, W.K.; et al. Phase 3 Trial of Nemolizumab in Patients with Prurigo Nodularis. N. Engl. J. Med. 2023, 389, 1579–1589. [Google Scholar] [CrossRef]
- Sofen, H.; Bissonnette, R.; Yosipovitch, G.; Silverberg, J.I.; Tyring, S.; Loo, W.J.; Zook, M.; Lee, M.; Zou, L.; Jiang, G.-L.; et al. Efficacy and Safety of Vixarelimab, a Human Monoclonal Oncostatin M Receptor β Antibody, in Moderate-to-Severe Prurigo Nodularis: A Randomised, Double-Blind, Placebo-Controlled, Phase 2a Study. EClinicalMedicine 2023, 57, 101826. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, H.; Masuoka, M.; Ohta, S.; Suzuki, S.; Arima, K.; Taniguchi, K.; Aoki, S.; Toda, S.; Yoshimoto, T.; Inagaki, N.; et al. Periostin Contributes to the Pathogenesis of Atopic Dermatitis by Inducing TSLP Production from Keratinocytes. Allergol. Int. 2012, 61, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, H.; Yu, J.; Su, Y.; Cao, S.; An, X.; Ren, X. Increased Prevalence of Regulatory T Cells in the Lung Cancer Microenvironment: A Role of Thymic Stromal Lymphopoietin. Cancer Immunol. Immunother. 2011, 60, 1587–1596. [Google Scholar] [CrossRef] [PubMed]
- Demehri, S.; Turkoz, A.; Manivasagam, S.; Yockey, L.J.; Turkoz, M.; Kopan, R. Elevated Epidermal Thymic Stromal Lymphopoietin Levels Establish an Antitumor Environment in the Skin. Cancer Cell 2012, 22, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Rochman, I.; Watanabe, N.; Arima, K.; Liu, Y.-J.; Leonard, W.J. Cutting Edge: Direct Action of Thymic Stromal Lymphopoietin on Activated Human CD4+ T Cells. J. Immunol. 2007, 178, 6720–6724. [Google Scholar] [CrossRef] [PubMed]
- Tuzova, M.; Richmond, J.; Wolpowitz, D.; Curiel-Lewandrowski, C.; Chaney, K.; Kupper, T.; Cruikshank, W. CCR4+T Cell Recruitment to the Skin in Mycosis Fungoides: Potential Contributions by Thymic Stromal Lymphopoietin and Interleukin-16. Leuk. Lymphoma 2015, 56, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Miyagaki, T.; Sugaya, M.; Fujita, H.; Saeki, H.; Tamaki, K. Increased Serum Thymic Stromal Lymphopoietin Levels in Patients with Cutaneous T Cell Lymphoma. Clin. Exp. Dermatol. 2009, 34, 539–540. [Google Scholar] [CrossRef] [PubMed]
- Hoy, S.M. Tezepelumab: First Approval. Drugs 2022, 82, 461–468. [Google Scholar] [CrossRef]
- Simpson, E.L.; Parnes, J.R.; She, D.; Crouch, S.; Rees, W.; Mo, M.; van der Merwe, R. Tezepelumab, an Anti-Thymic Stromal Lymphopoietin Monoclonal Antibody, in the Treatment of Moderate to Severe Atopic Dermatitis: A Randomized Phase 2a Clinical Trial. J. Am. Acad. Dermatol. 2019, 80, 1013–1021. [Google Scholar] [CrossRef]
- Luo, J.; Zhu, Z.; Zhai, Y.; Zeng, J.; Li, L.; Wang, D.; Deng, F.; Chang, B.; Zhou, J.; Sun, L. The Role of TSLP in Atopic Dermatitis: From Pathogenetic Molecule to Therapeutical Target. Mediat. Inflamm. 2023, 2023, 7697699. [Google Scholar] [CrossRef]
- Furue, M.; Furue, M. Interleukin-31 and Pruritic Skin. J. Clin. Med. 2021, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- Kawana, Y.; Suga, H.; Kamijo, H.; Miyagaki, T.; Sugaya, M.; Sato, S. Roles of OX40 and OX40 Ligand in Mycosis Fungoides and Sézary Syndrome. Int. J. Mol. Sci. 2021, 22, 12576. [Google Scholar] [CrossRef]
- Sernicola, A.; Ciolfi, C.; Miceli, P.; Alaibac, M. Cutaneous Lymphoma and Antibody-Directed Therapies. Antibodies 2023, 12, 21. [Google Scholar] [CrossRef]


{kind=link}
| Pathway | Drug | Target |
|---|---|---|
| IL-4 and IL-13 | Dupilumab | IL-4Ra |
| Lebrikizumab | IL-13 | |
| Tralokinumab | IL-13 | |
| IL-31 | BMS-981164 | IL-31 |
| Nemolizumab | IL-31RA | |
| Vixarelimab | OSMRB | |
| TSLP | Tezepelumab | TSLPR |
| MK-8226 | TSLP | |
| Ecleralimab | TSLP | |
| OX40-OX40L | Rocatinlimab | OX40 |
| Telazorlimab | OX40 | |
| Amlitelimab | OX40L | |
| IL-33 | Itepekimab | IL-33 |
| Etokimab | IL-33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzetto, R.; Miceli, P.; Tartaglia, J.; Ciolfi, C.; Sernicola, A.; Alaibac, M. Role of IL-4 and IL-13 in Cutaneous T Cell Lymphoma. Life 2024, 14, 245. https://doi.org/10.3390/life14020245
Mazzetto R, Miceli P, Tartaglia J, Ciolfi C, Sernicola A, Alaibac M. Role of IL-4 and IL-13 in Cutaneous T Cell Lymphoma. Life. 2024; 14(2):245. https://doi.org/10.3390/life14020245
Chicago/Turabian StyleMazzetto, Roberto, Paola Miceli, Jacopo Tartaglia, Christian Ciolfi, Alvise Sernicola, and Mauro Alaibac. 2024. "Role of IL-4 and IL-13 in Cutaneous T Cell Lymphoma" Life 14, no. 2: 245. https://doi.org/10.3390/life14020245
APA StyleMazzetto, R., Miceli, P., Tartaglia, J., Ciolfi, C., Sernicola, A., & Alaibac, M. (2024). Role of IL-4 and IL-13 in Cutaneous T Cell Lymphoma. Life, 14(2), 245. https://doi.org/10.3390/life14020245