
Harnessing γδ T Cells against Human Gynecologic Cancers

 ,
,
Abstract
:1. Introduction
1.1. γδ T Cell Subsets: Humans Are Not Mice
1.2. γδ T Cells in the Healthy Human Female Reproductive Tract
1.3. Activation of Cytotoxic and Non-Cytotoxic Functions of γδ T Cells: The Key Role of Butyrophilins
1.4. γδ T Cells in Human Ovarian Cancer
1.5. γδ T Cells in Other Human Ovarian Cancer Gynecologic Cancers and Non-Gynecological Malignancies
1.6. Potential of γδ T Cells in Anti-Cancer Cellular Therapies in Gynecologic Cancers
1.7. Modulating the Phenotype of γδ T Cells in Cancer Patients with Drugs or Antibodies
2. Concluding Remarks

{kind=link}
{kind=link}
| Tumor | Intervention | Trials | References |
|---|---|---|---|
| Multiple advanced-stage | BTN3A1 agonistic Abs | NCT04243499 NCT05307874 | [9] |
| CLL/MM/AML | Vγ9 TCR × CD1d bispecific Abs | NCT04887259 | [85] |
| Lung, liver, AML post-BM transplant | Allogeneic Vγ9Vδ2 T cells | NCT03183219 NCT03183232 NCT05015426 | [86] |
| MRD+ AML | Allogeneic Vδ1 T cells | NCT05001451 | [87] |
| Glioblastoma | Temozolomide-resistant γδ T cells | NCT04165941 | [88] |
| Relapsed/refractory solid tumors | NKG2D CAR Vγ9Vδ2 T cells | NCT04107142 | [89] |
| B cell malignancies | Anti-CD20 CAR Vδ1 T cells | NCT04735471 | [77] |
Author Contributions
Funding
Conflicts of Interest
References
- Nielsen, M.M.; Witherden, D.A.; Havran, W.L. γδ T cells in homeostasis and host defence of epithelial barrier tissues. Nat. Rev. Immunol. 2017, 17, 733–745. [Google Scholar] [CrossRef]
- Costa, G.P.; Mensurado, S.; Silva-Santos, B. Therapeutic avenues for γδ T cells in cancer. J. Immunother. Cancer 2023, 11, e007955. [Google Scholar] [CrossRef]
- Payne, K.K.; Mine, J.A.; Biswas, S.; Chaurio, R.A.; Perales-Puchalt, A.; Anadon, C.M.; Costich, T.L.; Harro, C.M.; Walrath, J.; Ming, Q.; et al. BTN3A1 governs antitumor responses by coordinating αβ and γδ T cells. Science 2020, 369, 942–949. [Google Scholar] [CrossRef]
- Conejo-Garcia, J.R.; Innamarato, P. γδ T cells share the spotlight in cancer. Nat. Cancer 2022, 3, 657–658. [Google Scholar] [CrossRef]
- Wu, Y.; Biswas, D.; Usaite, I.; Angelova, M.; Boeing, S.; Karasaki, T.; Veeriah, S.; Czyzewska-Khan, J.; Morton, C.; Joseph, M.; et al. A local human V 1 T cell population is associated with survival in non-small 1 cell lung cancer. Nat. Cancer 2022, in press.
- Rancan, C.; Arias-Badia, M.; Dogra, P.; Chen, B.; Aran, D.; Yang, H.; Luong, D.; Ilano, A.; Li, J.; Chang, H.; et al. Exhausted intratumoral Vδ2− γδ T cells in human kidney cancer retain effector function. Nat. Immunol. 2023, 24, 612–624. [Google Scholar] [CrossRef]
- Mensurado, S.; Blanco-Dominguez, R.; Silva-Santos, B. The emerging roles of γδ T cells in cancer immunotherapy. Nat. Rev. Clin. Oncol. 2023, 20, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Zhang, Q.; Ye, J.; Wang, F.; Zhang, Y.; Wevers, E.; Schwartz, T.; Hunborg, P.; Varvares, M.A.; Hoft, D.F.; et al. Tumor-infiltrating γδ T lymphocytes predict clinical outcome in human breast cancer. J. Immunol. 2012, 189, 5029–5036. [Google Scholar] [CrossRef] [PubMed]
- De Gassart, A.; Le, K.S.; Brune, P.; Agaugue, S.; Sims, J.; Goubard, A.; Castellano, R.; Joalland, N.; Scotet, E.; Collette, Y.; et al. Development of ICT01, a first-in-class, anti-BTN3A antibody for activating Vγ9Vδ2 T cell-mediated antitumor immune response. Sci. Transl. Med. 2021, 13, eabj0835. [Google Scholar] [CrossRef] [PubMed]
- Conejo-Garcia, J.R.; Benencia, F.; Courreges, M.C.; Gimotty, P.A.; Khang, E.; Buckanovich, R.J.; Frauwirth, K.A.; Zhang, L.; Katsaros, D.; Thompson, C.B.; et al. Ovarian carcinoma expresses the NKG2D ligand Letal and promotes the survival and expansion of CD28- antitumor T cells. Cancer Res. 2004, 64, 2175–2182. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, M.R.; Stephen, T.L.; Svoronos, N.; Allegrezza, M.J.; Tesone, A.J.; Perales-Puchalt, A.; Brencicova, E.; Escovar-Fadul, X.; Nguyen, J.M.; Cadungog, M.G.; et al. Microbially driven TLR5-dependent signaling governs distal malignant progression through tumor-promoting inflammation. Cancer Cell 2015, 27, 27–40. [Google Scholar] [CrossRef]
- Ribot, J.C.; Ribeiro, S.T.; Correia, D.V.; Sousa, A.E.; Silva-Santos, B. Human γδ thymocytes are functionally immature and differentiate into cytotoxic type 1 effector T cells upon IL-2/IL-15 signaling. J. Immunol. 2014, 192, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Kyle-Cezar, F.; Woolf, R.T.; Naceur-Lombardelli, C.; Owen, J.; Biswas, D.; Lorenc, A.; Vantourout, P.; Gazinska, P.; Grigoriadis, A.; et al. An innate-like Vδ1+ γδ T cell compartment in the human breast is associated with remission in triple-negative breast cancer. Sci. Transl. Med. 2019, 11, eaax9364. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.L.; Thomas, P.G. A Cell for the Ages: Human γδ T Cells across the Lifespan. Int. J. Mol. Sci. 2020, 21, 8903. [Google Scholar] [CrossRef] [PubMed]
- Fleming, C.; Morrissey, S.; Cai, Y.; Yan, J. γδ T Cells: Unexpected Regulators of Cancer Development and Progression. Trends Cancer 2017, 3, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Di Marco Barros, R.; Roberts, N.A.; Dart, R.J.; Vantourout, P.; Jandke, A.; Nussbaumer, O.; Deban, L.; Cipolat, S.; Hart, R.; Iannitto, M.L.; et al. Epithelia Use Butyrophilin-like Molecules to Shape Organ-Specific γδ T Cell Compartments. Cell 2016, 167, 203–218.e17. [Google Scholar] [CrossRef]
- Suzuki, T.; Hayman, L.; Kilbey, A.; Edwards, J.; Coffelt, S.B. Gut γδ T cells as guardians, disruptors, and instigators of cancer. Immunol. Rev. 2020, 298, 198–217. [Google Scholar] [CrossRef]
- Pitard, V.; Roumanes, D.; Lafarge, X.; Couzi, L.; Garrigue, I.; Lafon, M.E.; Merville, P.; Moreau, J.F.; Dechanet-Merville, J. Long-term expansion of effector/memory Vδ2− γδ T cells is a specific blood signature of CMV infection. Blood 2008, 112, 1317–1324. [Google Scholar] [CrossRef]
- Davey, M.S.; Willcox, C.R.; Baker, A.T.; Hunter, S.; Willcox, B.E. Recasting Human Vδ1 Lymphocytes in an Adaptive Role. Trends Immunol. 2018, 39, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Spada, F.M.; Grant, E.P.; Peters, P.J.; Sugita, M.; Melian, A.; Leslie, D.S.; Lee, H.K.; van Donselaar, E.; Hanson, D.A.; Krensky, A.M.; et al. Self-recognition of CD1 by γ/δ T cells: Implications for innate immunity. J. Exp. Med. 2000, 191, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Luoma, A.M.; Castro, C.D.; Mayassi, T.; Bembinster, L.A.; Bai, L.; Picard, D.; Anderson, B.; Scharf, L.; Kung, J.E.; Sibener, L.V.; et al. Crystal structure of Vδ1 T cell receptor in complex with CD1d-sulfatide shows MHC-like recognition of a self-lipid by human γδ T cells. Immunity 2013, 39, 1032–1042. [Google Scholar] [CrossRef]
- Reijneveld, J.F.; Ocampo, T.A.; Shahine, A.; Gully, B.S.; Vantourout, P.; Hayday, A.C.; Rossjohn, J.; Moody, D.B.; Van Rhijn, I. Human γδ T cells recognize CD1b by two distinct mechanisms. Proc. Natl. Acad. Sci. USA 2020, 117, 22944–22952. [Google Scholar] [CrossRef]
- Zeng, X.; Wei, Y.L.; Huang, J.; Newell, E.W.; Yu, H.; Kidd, B.A.; Kuhns, M.S.; Waters, R.W.; Davis, M.M.; Weaver, C.T.; et al. γδ T cells recognize a microbial encoded B cell antigen to initiate a rapid antigen-specific interleukin-17 response. Immunity 2012, 37, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Harly, C.; Joyce, S.P.; Domblides, C.; Bachelet, T.; Pitard, V.; Mannat, C.; Pappalardo, A.; Couzi, L.; Netzer, S.; Massara, L.; et al. Human γδ T cell sensing of AMPK-dependent metabolic tumor reprogramming through TCR recognition of EphA2. Sci. Immunol. 2021, 6, eaba9010. [Google Scholar] [CrossRef]
- Kenna, T.; Golden-Mason, L.; Norris, S.; Hegarty, J.E.; O’Farrelly, C.; Doherty, D.G. Distinct subpopulations of γδ T cells are present in normal and tumor-bearing human liver. Clin. Immunol. 2004, 113, 56–63. [Google Scholar] [CrossRef]
- Rice, M.T.; von Borstel, A.; Chevour, P.; Awad, W.; Howson, L.J.; Littler, D.R.; Gherardin, N.A.; Le Nours, J.; Giles, E.M.; Berry, R.; et al. Recognition of the antigen-presenting molecule MR1 by a Vδ3+ γδ T cell receptor. Proc. Natl. Acad. Sci. USA 2021, 118, e2110288118. [Google Scholar] [CrossRef]
- Le Nours, J.; Gherardin, N.A.; Ramarathinam, S.H.; Awad, W.; Wiede, F.; Gully, B.S.; Khandokar, Y.; Praveena, T.; Wubben, J.M.; Sandow, J.J.; et al. A class of γδ T cell receptors recognize the underside of the antigen-presenting molecule MR1. Science 2019, 366, 1522–1527. [Google Scholar] [CrossRef] [PubMed]
- Flynn, L.; Byrne, B.; Carton, J.; Kelehan, P.; O’Herlihy, C.; O’Farrelly, C. Menstrual cycle dependent fluctuations in NK and T-lymphocyte subsets from non-pregnant human endometrium. Am. J. Reprod. Immunol. 2000, 43, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gilman-Sachs, A.; Kwak-Kim, J. Ovarian and endometrial immunity during the ovarian cycle. J. Reprod. Immunol. 2019, 133, 7–14. [Google Scholar] [CrossRef]
- Strbo, N.; Romero, L.; Alcaide, M.; Fischl, M. Isolation and Flow Cytometric Analysis of Human Endocervical Gamma Delta T Cells. J. Vis. Exp. 2017, e55038. [Google Scholar] [CrossRef]
- Kang, S.; Wu, Q.; Huang, J.; Yang, B.; Liang, C.; Chi, P.; Wu, C. Tissue Resident Memory γδ T Cells in Murine Uterus Expressed High Levels of IL-17 Promoting the Invasion of Trophocytes. Front. Immunol. 2020, 11, 588227. [Google Scholar] [CrossRef] [PubMed]
- Mincheva-Nilsson, L. Pregnancy and gamma/delta T cells: Taking on the hard questions. Reprod. Biol. Endocrinol. 2003, 1, 120. [Google Scholar] [CrossRef] [PubMed]
- Monin, L.; Ushakov, D.S.; Arnesen, H.; Bah, N.; Jandke, A.; Munoz-Ruiz, M.; Carvalho, J.; Joseph, S.; Almeida, B.C.; Green, M.J.; et al. γδ T cells compose a developmentally regulated intrauterine population and protect against vaginal candidiasis. Mucosal Immunol. 2020, 13, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Deseke, M.; Prinz, I. Ligand recognition by the γδ TCR and discrimination between homeostasis and stress conditions. Cell Mol. Immunol. 2020, 17, 914–924. [Google Scholar] [CrossRef]
- Luoma, A.M.; Castro, C.D.; Adams, E.J. γδ T cell surveillance via CD1 molecules. Trends Immunol. 2014, 35, 613–621. [Google Scholar] [CrossRef]
- Brandes, M.; Willimann, K.; Bioley, G.; Levy, N.; Eberl, M.; Luo, M.; Tampe, R.; Levy, F.; Romero, P.; Moser, B. Cross-presenting human γδ T cells induce robust CD8+ αβ T cell responses. Proc. Natl. Acad. Sci. USA 2009, 106, 2307–2312. [Google Scholar] [CrossRef]
- Couzi, L.; Pitard, V.; Sicard, X.; Garrigue, I.; Hawchar, O.; Merville, P.; Moreau, J.F.; Dechanet-Merville, J. Antibody-dependent anti-cytomegalovirus activity of human γδ T cells expressing CD16 (FcγRIIIa). Blood 2012, 119, 1418–1427. [Google Scholar] [CrossRef]
- Biswas, S.; Mandal, G.; Payne, K.K.; Anadon, C.M.; Gatenbee, C.D.; Chaurio, R.A.; Costich, T.L.; Moran, C.; Harro, C.M.; Rigolizzo, K.E.; et al. IgA transcytosis and antigen recognition govern ovarian cancer immunity. Nature 2021, 591, 464–470. [Google Scholar] [CrossRef]
- Conejo-Garcia, J.R.; Biswas, S.; Chaurio, R.; Rodriguez, P.C. Neglected no more: B cell-mediated anti-tumor immunity. Semin. Immunol. 2023, 65, 101707. [Google Scholar] [CrossRef]
- Mandal, G.; Biswas, S.; Anadon, C.M.; Yu, X.; Gatenbee, C.D.; Prabhakaran, S.; Payne, K.K.; Chaurio, R.A.; Martin, A.; Innamarato, P.; et al. IgA-Dominated Humoral Immune Responses Govern Patients’ Outcome in Endometrial Cancer. Cancer Res. 2022, 82, 859–871. [Google Scholar] [CrossRef]
- Arnett, H.A.; Escobar, S.S.; Gonzalez-Suarez, E.; Budelsky, A.L.; Steffen, L.A.; Boiani, N.; Zhang, M.; Siu, G.; Brewer, A.W.; Viney, J.L. BTNL2, a butyrophilin/B7-like molecule, is a negative costimulatory molecule modulated in intestinal inflammation. J. Immunol. 2007, 178, 1523–1533. [Google Scholar] [CrossRef]
- Abeler-Dorner, L.; Swamy, M.; Williams, G.; Hayday, A.C.; Bas, A. Butyrophilins: An emerging family of immune regulators. Trends Immunol. 2012, 33, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.A.; Reith, W.; Trowsdale, J. Regulation of Immunity by Butyrophilins. Annu. Rev. Immunol. 2016, 34, 151–172. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, C.C.; Nguyen, K.; Jin, Y.; Vinogradova, O.; Wiemer, A.J. Ligand-induced interactions between butyrophilin 2A1 and 3A1 internal domains in the HMBPP receptor complex. Cell Chem. Biol. 2022, 29, 985–995.e5. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, M.M.; Willcox, C.R.; Salim, M.; Paletta, D.; Fichtner, A.S.; Noll, A.; Starick, L.; Nohren, A.; Begley, C.R.; Berwick, K.A.; et al. Butyrophilin-2A1 Directly Binds Germline-Encoded Regions of the Vγ9Vδ2 TCR and Is Essential for Phosphoantigen Sensing. Immunity 2020, 52, 487–498.e6. [Google Scholar] [CrossRef]
- Rigau, M.; Ostrouska, S.; Fulford, T.S.; Johnson, D.N.; Woods, K.; Ruan, Z.; McWilliam, H.E.G.; Hudson, C.; Tutuka, C.; Wheatley, A.K.; et al. Butyrophilin 2A1 is essential for phosphoantigen reactivity by γδ T cells. Science 2020, 367, eaay5516. [Google Scholar] [CrossRef]
- Willcox, C.R.; Vantourout, P.; Salim, M.; Zlatareva, I.; Melandri, D.; Zanardo, L.; George, R.; Kjaer, S.; Jeeves, M.; Mohammed, F.; et al. Butyrophilin-like 3 Directly Binds a Human Vγ4+ T Cell Receptor Using a Modality Distinct from Clonally-Restricted Antigen. Immunity 2019, 51, 813–825.e4. [Google Scholar] [CrossRef]
- Zhang, L.; Conejo-Garcia, J.R.; Katsaros, D.; Gimotty, P.A.; Massobrio, M.; Regnani, G.; Makrigiannakis, A.; Gray, H.; Schlienger, K.; Liebman, M.N.; et al. Intratumoral T cells, recurrence, and survival in epithelial ovarian cancer. N. Engl. J. Med. 2003, 348, 203–213. [Google Scholar] [CrossRef]
- Sato, E.; Olson, S.H.; Ahn, J.; Bundy, B.; Nishikawa, H.; Qian, F.; Jungbluth, A.A.; Frosina, D.; Gnjatic, S.; Ambrosone, C.; et al. Intraepithelial CD8+ tumor-infiltrating lymphocytes and a high CD8+/regulatory T cell ratio are associated with favorable prognosis in ovarian cancer. Proc. Natl. Acad. Sci. USA 2005, 102, 18538–18543. [Google Scholar] [CrossRef]
- Groh, V.; Rhinehart, R.; Secrist, H.; Bauer, S.; Grabstein, K.H.; Spies, T. Broad tumor-associated expression and recognition by tumor-derived γδ T cells of MICA and MICB. Proc. Natl. Acad. Sci. USA 1999, 96, 6879–6884. [Google Scholar] [CrossRef]
- Chen, X.; Shang, W.; Xu, R.; Wu, M.; Zhang, X.; Huang, P.; Wang, F.; Pan, S. Distribution and functions of γδ T cells infiltrated in the ovarian cancer microenvironment. J. Transl. Med. 2019, 17, 144. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, J.; Wang, D.; Cai, M.; Xu, Y.; Hu, Y.; Chen, H.; He, W.; Zhang, J. Anti-PD-1 antibody armored γδ T cells enhance anti-tumor efficacy in ovarian cancer. Signal Transduct. Target. Ther. 2023, 8, 399. [Google Scholar] [CrossRef] [PubMed]
- Harmon, C.; Zaborowski, A.; Moore, H.; St Louis, P.; Slattery, K.; Duquette, D.; Scanlan, J.; Kane, H.; Kunkemoeller, B.; McIntyre, C.L.; et al. γδ T cell dichotomy with opposing cytotoxic and wound healing functions in human solid tumors. Nat. Cancer 2023, 4, 1122–1137. [Google Scholar] [CrossRef]
- Lheureux, S.; Matei, D.E.; Konstantinopoulos, P.A.; Wang, B.X.; Gadalla, R.; Block, M.S.; Jewell, A.; Gaillard, S.L.; McHale, M.; McCourt, C.; et al. Translational randomized phase II trial of cabozantinib in combination with nivolumab in advanced, recurrent, or metastatic endometrial cancer. J. Immunother. Cancer 2022, 10, e004233. [Google Scholar] [CrossRef]
- Wu, Y.; Ye, S.; Goswami, S.; Pei, X.; Xiang, L.; Zhang, X.; Yang, H. Clinical significance of peripheral blood and tumor tissue lymphocyte subsets in cervical cancer patients. BMC Cancer 2020, 20, 173. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Zhou, F. Effect of ex vivo-expanded γδ-T cells combined with galectin-1 antibody on the growth of human cervical cancer xenografts in SCID mice. Clin. Investig. Med. 2010, 33, E280–E289. [Google Scholar] [CrossRef]
- Zakeri, N.; Hall, A.; Swadling, L.; Pallett, L.J.; Schmidt, N.M.; Diniz, M.O.; Kucykowicz, S.; Amin, O.E.; Gander, A.; Pinzani, M.; et al. Characterisation and induction of tissue-resident gamma delta T-cells to target hepatocellular carcinoma. Nat. Commun. 2022, 13, 1372. [Google Scholar] [CrossRef]
- Wang, J.; Lin, C.; Li, H.; Li, R.; Wu, Y.; Liu, H.; Zhang, H.; He, H.; Zhang, W.; Xu, J. Tumor-infiltrating γδ T cells predict prognosis and adjuvant chemotherapeutic benefit in patients with gastric cancer. Oncoimmunology 2017, 6, e1353858. [Google Scholar] [CrossRef]
- Lu, H.; Dai, W.; Guo, J.; Wang, D.; Wen, S.; Yang, L.; Lin, D.; Xie, W.; Wen, L.; Fang, J.; et al. High Abundance of Intratumoral γδ T Cells Favors a Better Prognosis in Head and Neck Squamous Cell Carcinoma: A Bioinformatic Analysis. Front. Immunol. 2020, 11, 573920. [Google Scholar] [CrossRef]
- Nguyen, S.; Chevalier, M.F.; Benmerzoug, S.; Cesson, V.; Schneider, A.K.; Rodrigues-Dias, S.C.; Dartiguenave, F.; Lucca, I.; Jichlinski, P.; Roth, B.; et al. Vδ2 T cells are associated with favorable clinical outcomes in patients with bladder cancer and their tumor reactivity can be boosted by BCG and zoledronate treatments. J. Immunother. Cancer 2022, 10, e004880. [Google Scholar] [CrossRef]
- Davies, D.; Kamdar, S.; Woolf, R.; Zlatareva, I.; Iannitto, M.L.; Morton, C.; Haque, Y.; Martin, H.; Biswas, D.; Ndagire, S.; et al. PD-1 defines a distinct, functional, tissue-adapted state in Vδ1+ T cells with implications for cancer immunotherapy. Nat. Cancer 2024, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bruni, E.; Cimino, M.M.; Donadon, M.; Carriero, R.; Terzoli, S.; Piazza, R.; Ravens, S.; Prinz, I.; Cazzetta, V.; Marzano, P.; et al. Intrahepatic CD69+ Vδ1 T cells re-circulate in the blood of patients with metastatic colorectal cancer and limit tumor progression. J. Immunother. Cancer 2022, 10, e004579. [Google Scholar] [CrossRef] [PubMed]
- de Vries, N.L.; van de Haar, J.; Veninga, V.; Chalabi, M.; Ijsselsteijn, M.E.; van der Ploeg, M.; van den Bulk, J.; Ruano, D.; van den Berg, J.G.; Haanen, J.B.; et al. γδ T cells are effectors of immunotherapy in cancers with HLA class I defects. Nature 2023, 613, 743–750. [Google Scholar] [CrossRef]
- Anadon, C.M.; Yu, X.; Hanggi, K.; Biswas, S.; Chaurio, R.A.; Martin, A.; Payne, K.K.; Mandal, G.; Innamarato, P.; Harro, C.M.; et al. Ovarian cancer immunogenicity is governed by a narrow subset of progenitor tissue-resident memory T cells. Cancer Cell 2022, 40, 545–557.e513. [Google Scholar] [CrossRef]
- June, C.H.; O’Connor, R.S.; Kawalekar, O.U.; Ghassemi, S.; Milone, M.C. CAR T cell immunotherapy for human cancer. Science 2018, 359, 1361–1365. [Google Scholar] [CrossRef]
- Locke, F.L.; Miklos, D.B.; Jacobson, C.A.; Perales, M.A.; Kersten, M.J.; Oluwole, O.O.; Ghobadi, A.; Rapoport, A.P.; McGuirk, J.; Pagel, J.M.; et al. Axicabtagene Ciloleucel as Second-Line Therapy for Large B-Cell Lymphoma. N. Engl. J. Med. 2022, 386, 640–654. [Google Scholar] [CrossRef]
- Neelapu, S.S.; Dickinson, M.; Munoz, J.; Ulrickson, M.L.; Thieblemont, C.; Oluwole, O.O.; Herrera, A.F.; Ujjani, C.S.; Lin, Y.; Riedell, P.A.; et al. Axicabtagene ciloleucel as first-line therapy in high-risk large B-cell lymphoma: The phase 2 ZUMA-12 trial. Nat. Med. 2022, 28, 735–742. [Google Scholar] [CrossRef]
- Neelapu, S.S.; Locke, F.L.; Bartlett, N.L.; Lekakis, L.J.; Miklos, D.B.; Jacobson, C.A.; Braunschweig, I.; Oluwole, O.O.; Siddiqi, T.; Lin, Y.; et al. Axicabtagene Ciloleucel CAR T-Cell Therapy in Refractory Large B-Cell Lymphoma. N. Engl. J. Med. 2017, 377, 2531–2544. [Google Scholar] [CrossRef]
- Grupp, S.A.; Kalos, M.; Barrett, D.; Aplenc, R.; Porter, D.L.; Rheingold, S.R.; Teachey, D.T.; Chew, A.; Hauck, B.; Wright, J.F.; et al. Chimeric antigen receptor-modified T cells for acute lymphoid leukemia. N. Engl. J. Med. 2013, 368, 1509–1518. [Google Scholar] [CrossRef]
- Porter, D.L.; Levine, B.L.; Kalos, M.; Bagg, A.; June, C.H. Chimeric antigen receptor-modified T cells in chronic lymphoid leukemia. N. Engl. J. Med. 2011, 365, 725–733. [Google Scholar] [CrossRef]
- Raje, N.; Berdeja, J.; Lin, Y.; Siegel, D.; Jagannath, S.; Madduri, D.; Liedtke, M.; Rosenblatt, J.; Maus, M.V.; Turka, A.; et al. Anti-BCMA CAR T-Cell Therapy bb2121 in Relapsed or Refractory Multiple Myeloma. N. Engl. J. Med. 2019, 380, 1726–1737. [Google Scholar] [CrossRef]
- Conejo-Garcia, J.R.; Guevara-Patino, J.A. Barriers and Opportunities for CAR T-Cell Targeting of Solid Tumors. Immunol. Investig. 2022, 51, 2215–2225. [Google Scholar] [CrossRef]
- Liu, X.F.; Onda, M.; Schlomer, J.; Bassel, L.; Kozlov, S.; Tai, C.H.; Zhou, Q.; Liu, W.; Tsao, H.E.; Hassan, R.; et al. Tumor resistance to anti-mesothelin CAR-T cells caused by binding to shed mesothelin is overcome by targeting a juxtamembrane epitope. Proc. Natl. Acad. Sci. USA 2024, 121, e2317283121. [Google Scholar] [CrossRef]
- Perales-Puchalt, A.; Svoronos, N.; Rutkowski, M.R.; Allegrezza, M.J.; Tesone, A.J.; Payne, K.K.; Wickramasinghe, J.; Nguyen, J.M.; O’Brien, S.W.; Gumireddy, K.; et al. Follicle-Stimulating Hormone Receptor Is Expressed by Most Ovarian Cancer Subtypes and Is a Safe and Effective Immunotherapeutic Target. Clin. Cancer Res. 2017, 23, 441–453. [Google Scholar] [CrossRef]
- Xu, Y.; Xiang, Z.; Alnaggar, M.; Kouakanou, L.; Li, J.; He, J.; Yang, J.; Hu, Y.; Chen, Y.; Lin, L.; et al. Allogeneic Vγ9Vδ2 T-cell immunotherapy exhibits promising clinical safety and prolongs the survival of patients with late-stage lung or liver cancer. Cell Mol. Immunol. 2021, 18, 427–439. [Google Scholar] [CrossRef]
- Frieling, J.S.; Tordesillas, L.; Bustos, X.E.; Ramello, M.C.; Bishop, R.T.; Cianne, J.E.; Snedal, S.A.; Li, T.; Lo, C.H.; de la Iglesia, J.; et al. γδ-Enriched CAR-T cell therapy for bone metastatic castrate-resistant prostate cancer. Sci. Adv. 2023, 9, eadf0108. [Google Scholar] [CrossRef]
- Nishimoto, K.P.; Barca, T.; Azameera, A.; Makkouk, A.; Romero, J.M.; Bai, L.; Brodey, M.M.; Kennedy-Wilde, J.; Shao, H.; Papaioannou, S.; et al. Allogeneic CD20-targeted γδ T cells exhibit innate and adaptive antitumor activities in preclinical B-cell lymphoma models. Clin. Transl. Immunol. 2022, 11, e1373. [Google Scholar] [CrossRef]
- Ng, J.W.K.; Tan, K.W.; Guo, D.Y.; Lai, J.J.H.; Fan, X.; Poon, Z.; Lim, T.H.; Lim, A.S.T.; Lim, T.K.H.; Hwang, W.Y.K.; et al. Cord blood-derived Vδ2+ and Vδ2− T cells acquire differential cell state compositions upon in vitro expansion. Sci. Adv. 2023, 9, eadf3120. [Google Scholar] [CrossRef]
- Mirza, M.R.; Chase, D.M.; Slomovitz, B.M.; dePont Christensen, R.; Novak, Z.; Black, D.; Gilbert, L.; Sharma, S.; Valabrega, G.; Landrum, L.M.; et al. Dostarlimab for Primary Advanced or Recurrent Endometrial Cancer. N. Engl. J. Med. 2023, 388, 2145–2158. [Google Scholar] [CrossRef]
- Eskander, R.N.; Sill, M.W.; Beffa, L.; Moore, R.G.; Hope, J.M.; Musa, F.B.; Mannel, R.; Shahin, M.S.; Cantuaria, G.H.; Girda, E.; et al. Pembrolizumab plus Chemotherapy in Advanced Endometrial Cancer. N. Engl. J. Med. 2023, 388, 2159–2170. [Google Scholar] [CrossRef]
- Borella, F.; Ghisoni, E.; Giannone, G.; Cosma, S.; Benedetto, C.; Valabrega, G.; Katsaros, D. Immune Checkpoint Inhibitors in Epithelial Ovarian Cancer: An Overview on Efficacy and Future Perspectives. Diagnostics 2020, 10, 146. [Google Scholar] [CrossRef]
- Mamedov, M.R.; Vedova, S.; Freimer, J.W.; Sahu, A.D.; Ramesh, A.; Arce, M.M.; Meringa, A.D.; Ota, M.; Chen, P.A.; Hanspers, K.; et al. CRISPR screens decode cancer cell pathways that trigger γδ T cell detection. Nature 2023, 621, 188–195. [Google Scholar] [CrossRef]
- Crome, S.Q.; Nguyen, L.T.; Lopez-Verges, S.; Yang, S.Y.; Martin, B.; Yam, J.Y.; Johnson, D.J.; Nie, J.; Pniak, M.; Yen, P.H.; et al. A distinct innate lymphoid cell population regulates tumor-associated T cells. Nat. Med. 2017, 23, 368–375. [Google Scholar] [CrossRef]
- Melenhorst, J.J.; Chen, G.M.; Wang, M.; Porter, D.L.; Chen, C.; Collins, M.A.; Gao, P.; Bandyopadhyay, S.; Sun, H.; Zhao, Z.; et al. Decade-long leukaemia remissions with persistence of CD4+ CAR T cells. Nature 2022, 602, 503–509. [Google Scholar] [CrossRef]
- de Weerdt, I.; Lameris, R.; Ruben, J.M.; de Boer, R.; Kloosterman, J.; King, L.A.; Levin, M.D.; Parren, P.; de Gruijl, T.D.; Kater, A.P.; et al. A Bispecific Single-Domain Antibody Boosts Autologous Vγ9Vδ2-T Cell Responses Toward CD1d in Chronic Lymphocytic Leukemia. Clin. Cancer Res. 2021, 27, 1744–1755. [Google Scholar] [CrossRef]
- Boucher, J.C.; Yu, B.; Li, G.; Shrestha, B.; Sallman, D.; Landin, A.M.; Cox, C.; Karyampudi, K.; Anasetti, C.; Davila, M.L.; et al. Large Scale Ex Vivo Expansion of γδ T cells Using Artificial Antigen-presenting Cells. J. Immunother. 2023, 46, 5–13. [Google Scholar] [CrossRef]
- Almeida, A.R.; Correia, D.V.; Fernandes-Platzgummer, A.; da Silva, C.L.; da Silva, M.G.; Anjos, D.R.; Silva-Santos, B. Delta One T Cells for Immunotherapy of Chronic Lymphocytic Leukemia: Clinical-Grade Expansion/Differentiation and Preclinical Proof of Concept. Clin. Cancer Res. 2016, 22, 5795–5804. [Google Scholar] [CrossRef]
- Lamb, L.S.; Pereboeva, L.; Youngblood, S.; Gillespie, G.Y.; Nabors, L.B.; Markert, J.M.; Dasgupta, A.; Langford, C.; Spencer, H.T. A combined treatment regimen of MGMT-modified γδ T cells and temozolomide chemotherapy is effective against primary high grade gliomas. Sci. Rep. 2021, 11, 21133. [Google Scholar] [CrossRef]
- Ang, W.X.; Ng, Y.Y.; Xiao, L.; Chen, C.; Li, Z.; Chi, Z.; Tay, J.C.; Tan, W.K.; Zeng, J.; Toh, H.C.; et al. Electroporation of NKG2D RNA CAR Improves Vγ9Vδ2 T Cell Responses against Human Solid Tumor Xenografts. Mol. Ther. Oncolytics 2020, 17, 421–430. [Google Scholar] [CrossRef] [PubMed]
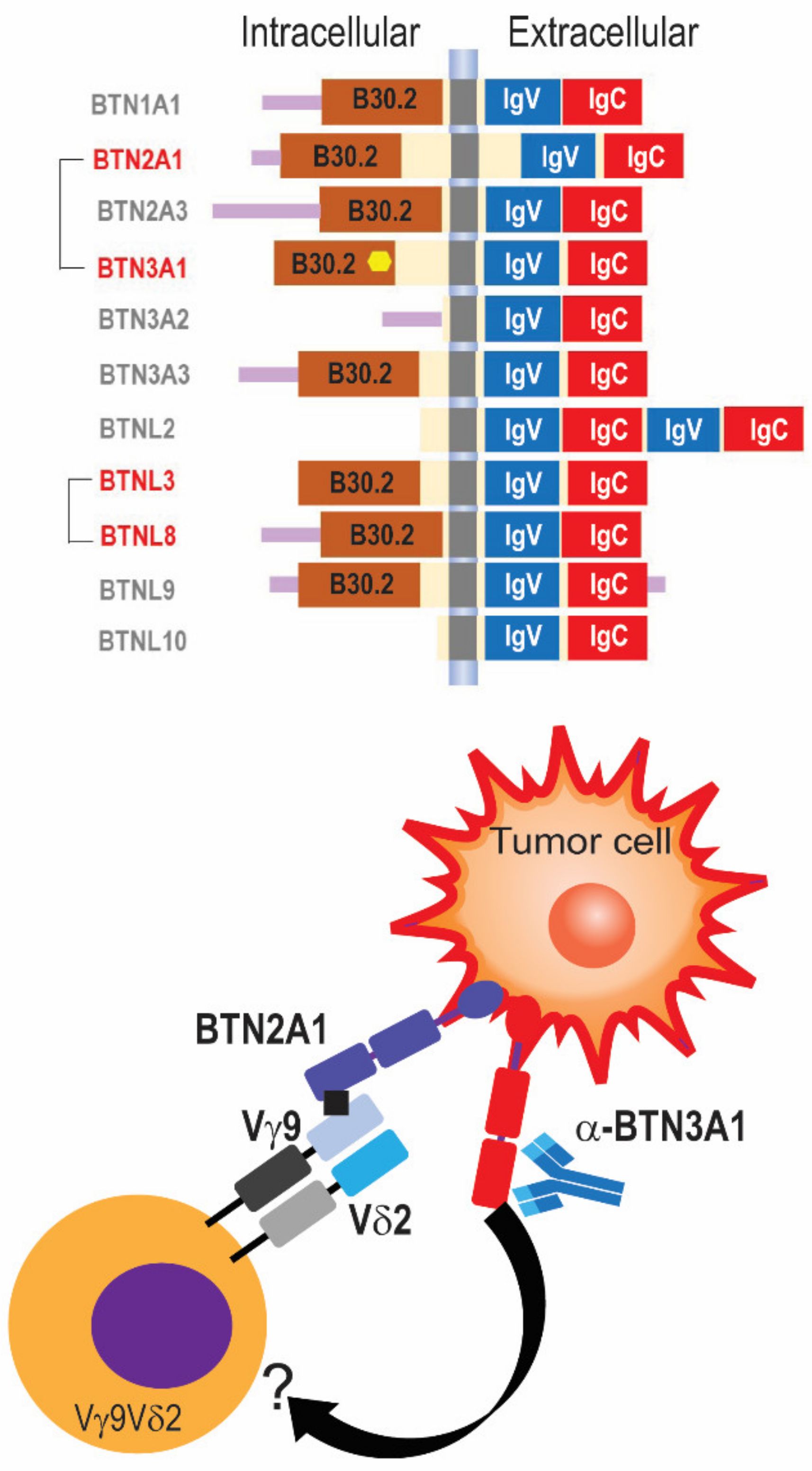
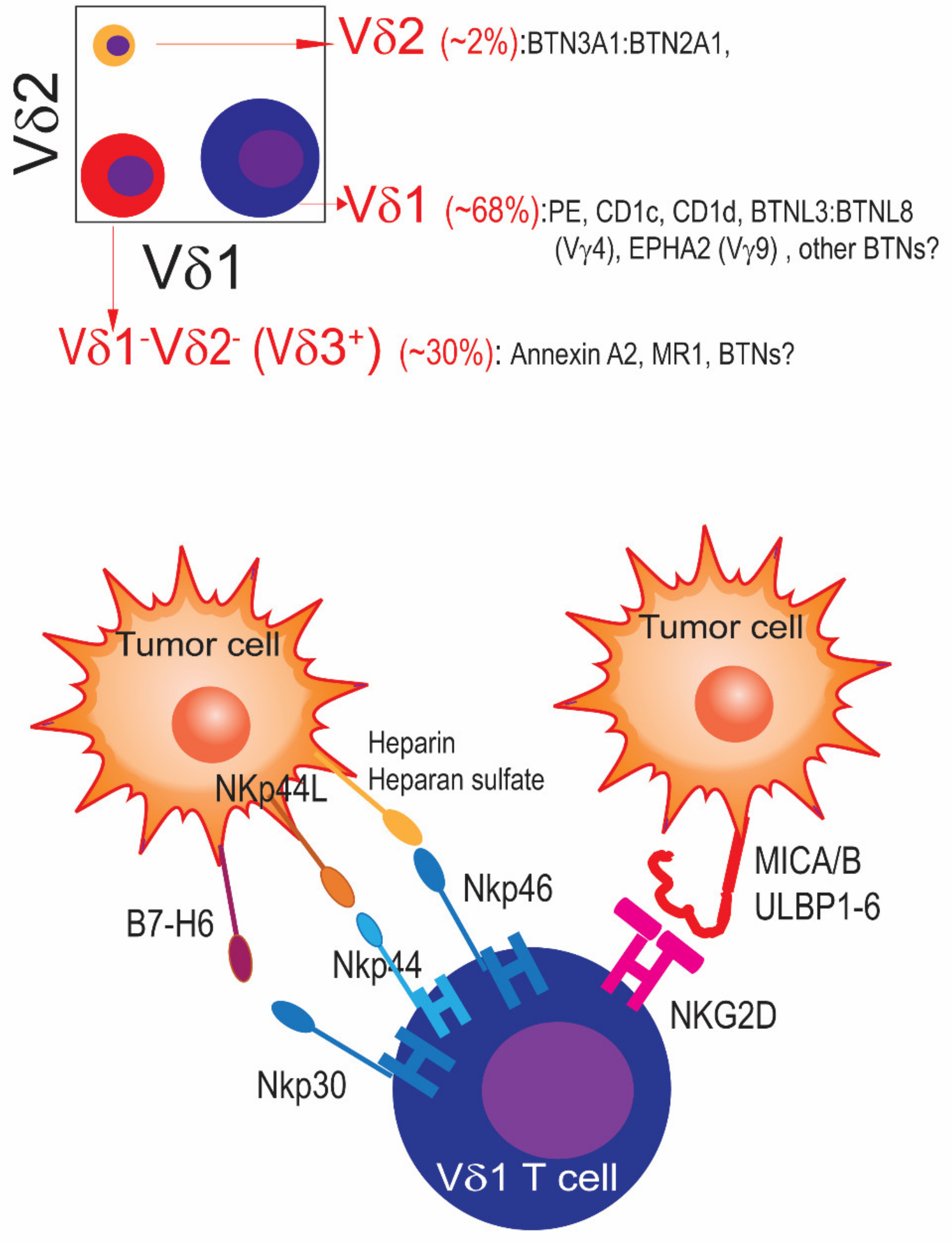
| Tumor | Marker/Subset | References |
|---|---|---|
| Ovarian carcinoma | Vδ1/Vδ3 γδ T cells | [3] |
| Endometrial carcinoma | TRD1 (Vδ1 marker) | [53] |
| Uterine carcinosarcoma | Tissue-resident memory γδ T cells | [54] |
| Cervical carcinoma | Total γδ T cells | [55] |
| Non-small-cell lung cancer | Tissue-resident memory Vδ1 γδ T cells | [5] |
| Breast cancer | Total γδ T cells | [8] |
| Renal cancer | PD-1+Vδ2neg γδ T cells | [6] |
| Hepatocarcinoma | Tissue-resident memory Vδ2neg γδ T cells | [56] |
| Gastric cancer | Total γδ T cells | [57] |
| Head and neck cancer | Total γδ T cells/butyrophilins | [58] |
| Bladder cancer | Vδ2 γδ T cells | [59] |
| Melanoma | PD-1+Vδ1+ γδ T cells | [60] |
| Colon carcinoma | CD69+Vδ1+ γδ T cells | [61] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conejo-Garcia, J.R.; Anadon, C.M.; Lopez-Bailon, L.U.; Chaurio, R.A. Harnessing γδ T Cells against Human Gynecologic Cancers. Life 2024, 14, 325. https://doi.org/10.3390/life14030325
Conejo-Garcia JR, Anadon CM, Lopez-Bailon LU, Chaurio RA. Harnessing γδ T Cells against Human Gynecologic Cancers. Life. 2024; 14(3):325. https://doi.org/10.3390/life14030325
Chicago/Turabian StyleConejo-Garcia, Jose R., Carmen M. Anadon, Luis U. Lopez-Bailon, and Ricardo A. Chaurio. 2024. "Harnessing γδ T Cells against Human Gynecologic Cancers" Life 14, no. 3: 325. https://doi.org/10.3390/life14030325
APA StyleConejo-Garcia, J. R., Anadon, C. M., Lopez-Bailon, L. U., & Chaurio, R. A. (2024). Harnessing γδ T Cells against Human Gynecologic Cancers. Life, 14(3), 325. https://doi.org/10.3390/life14030325