

Antivirulence Effects of Trans-Resveratrol and Curcumin on Methicillin-Resistant Staphylococcus aureus (MRSA) from Saudi Arabia


Abstract
1. Introduction
2. Materials and Methods
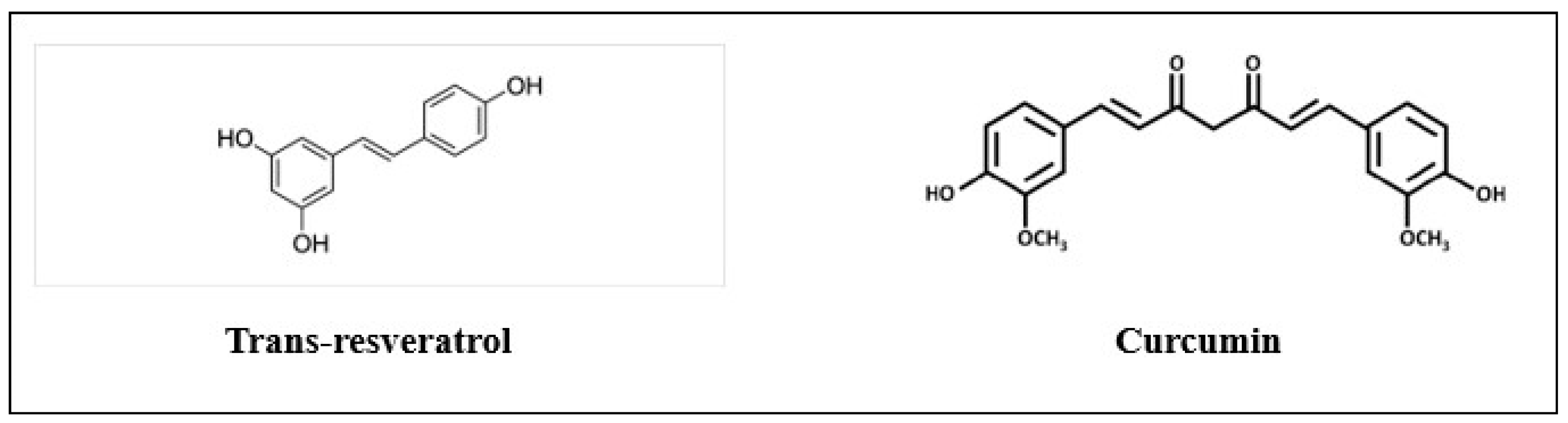
2.1. Trans-Resveratrol and Curcumin
2.2. Isolation and Identification of Bacterial Isolates
2.3. Minimum Inhibitory Concentration Assay of Trans-Resveratrol and Curcumin and Influence of Subinhibitory Concentrations on Bacterial Count
2.4. Effect of Trans-Resveratrol and Curcumin on Virulence Factors of MRSA
2.4.1. Biofilm Assay
2.4.2. Hemolysin Assay
2.4.3. Hemagglutination Assay
2.4.4. Total Protease Production
2.4.5. Assay of Lecithinase Activity
2.5. Statistical Analysis
3. Results
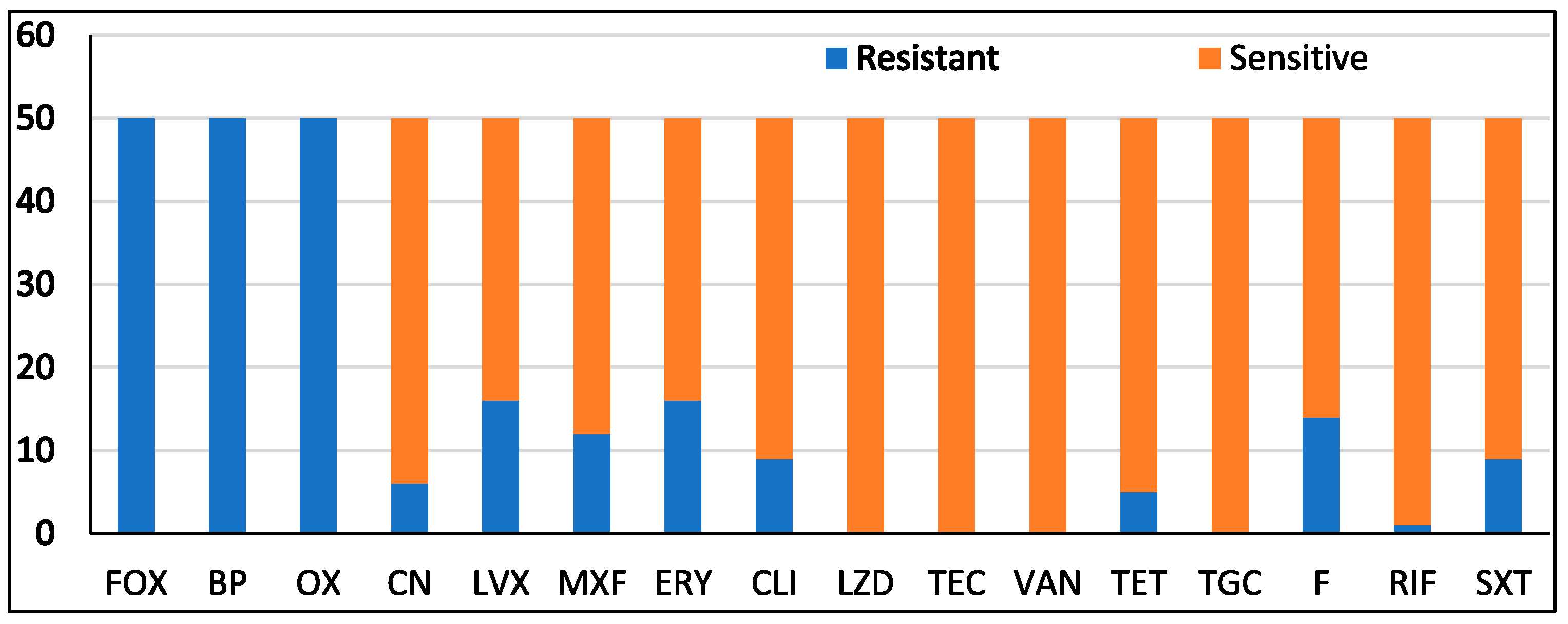
3.1. Isolation, Identification, and Antimicrobial Susceptibility Pattern of Bacterial Isolates
3.2. Determination of MICs of Trans-Resveratrol and Curcumin and Effect of Their Subinhibitory Concentrations on Bacterial Growth
3.3. Detection of Virulence Factors of MRSA Isolates
3.4. Virulence Profile Pattern of Studied Virulence Factors
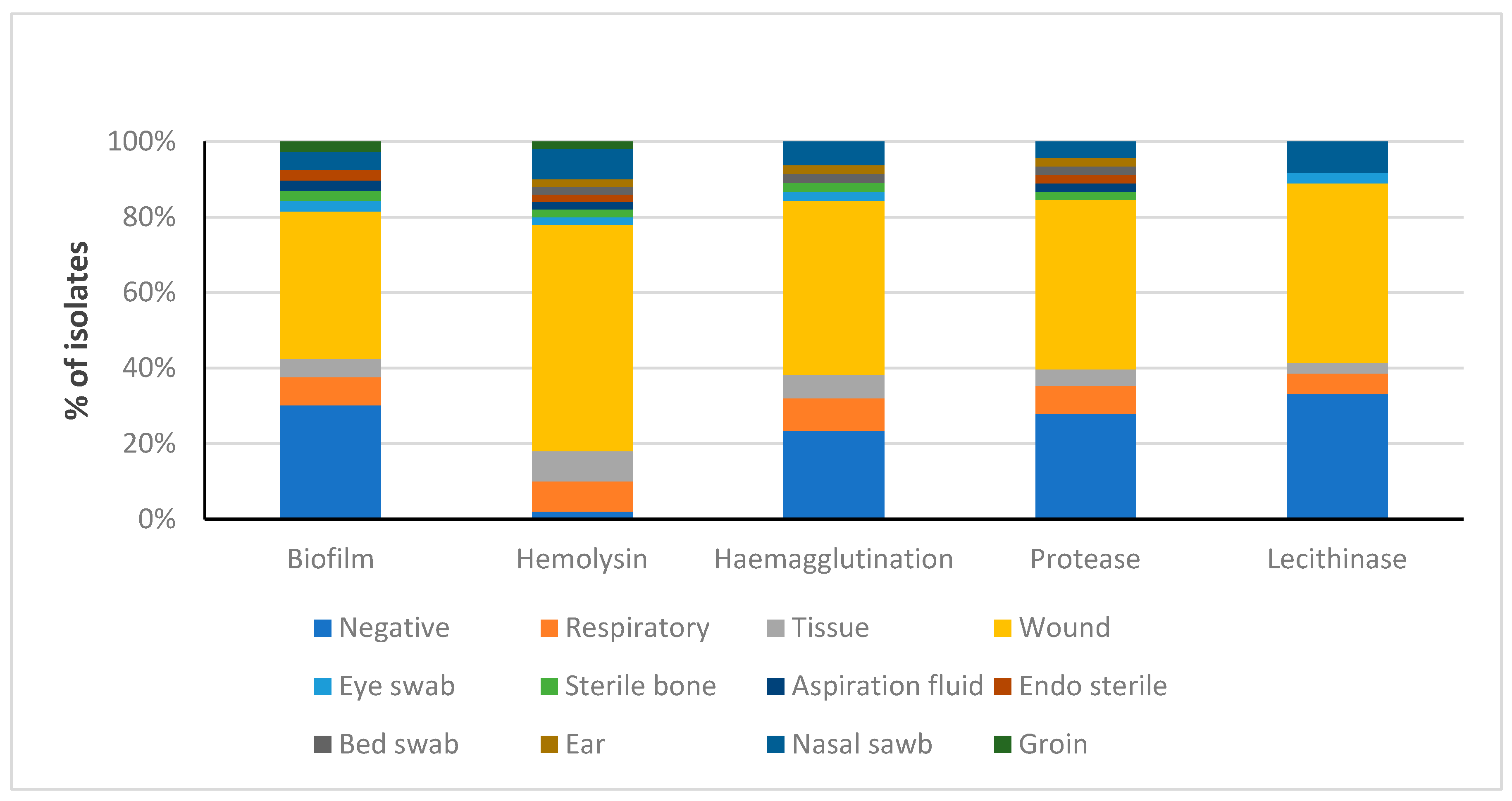
3.5. Distribution of Detected Virulence Factors among MRSA Obtained from Several Clinical Sources
3.6. Effect of Sub-MICs of Both Resveratrol and Curcumin on Virulence Factors of MRSA Isolates
3.6.1. Effect on Biofilm Production
3.6.2. Effect on Hemolysin
3.6.3. Effect on Hemagglutination
3.6.4. Effect on Total Protease Production
3.6.5. Effect on Lecithinase Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hoyert, D.L.; Xu, J. Deaths: Preliminary data for 2011. In National Vital Statistics Reports: From the Centers for Disease Control and Prevention, National Center for Health Statistics; National Vital Statistics System (US): Washington, DC, USA, 2012. [Google Scholar]
- WHO. 2017. Available online: https://www.who.int/publications/i/item/WHO-EMP-IAU-2017.12 (accessed on 25 February 2024).
- Batabyal, B.; Kundu, G.K.; Biswas, S. Methicillin-resistant Staphylococcus aureus: A brief review. Int. Res. J. Biol. Sci. 2012, 1, 65–71. [Google Scholar]
- Sharma, A.K.; Dhasmana, N.; Dubey, N.; Kumar, N.; Gangwal, A.; Gupta, M.; Singh, Y. Bacterial virulence factors: Secreted for survival. Indian J. Microbiol. 2017, 57, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jevon, M.; Guo, C.; Ma, B.; Mordan, N.; Nair, S.P.; Harris, M.; Henderson, B.; Bentley, G.; Meghji, S. Mechanisms of internalization of Staphylococcus aureus by cultured human osteoblasts. Infect. Immun. 1999, 67, 2677–2681. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, M.L.; Richardson, K.; Vaughan Sarrazin, M.S.; Goto, M.; Livorsi, D.J.; Nair, R.; Alexander, B.; Beck, B.F.; Jones, M.P.; Puig-Asensio, M. Comparative effectiveness of switching to daptomycin versus remaining on vancomycin among patients with methicillin-resistant Staphylococcus aureus (MRSA) bloodstream infections. Clin. Infect. Dis. 2021, 72 (Suppl. S1), S68–S73. [Google Scholar] [CrossRef]
- Wu, S.-C.; Liu, F.; Zhu, K.; Shen, J.-Z. Natural products that target virulence factors in antibiotic-resistant Staphylococcus aureus. J. Agric. Food Chem. 2019, 67, 13195–13211. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Li, M.; Hao, Z.; Shen, X.; Liu, L.; Jin, Y.; Wang, S.; Guo, Y.; Yang, L.; Wang, L. Subinhibitory concentrations of resveratrol reduce alpha-hemolysin production in Staphylococcus aureus isolates by downregulating saeRS. Emerg. Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-C.; Tseng, C.-H.; Wang, P.-W.; Lu, P.-L.; Weng, Y.-H.; Yen, F.-L.; Fang, J.-Y. Pterostilbene, a methoxylated resveratrol derivative, efficiently eradicates planktonic, biofilm, and intracellular MRSA by topical application. Front. Microbiol. 2017, 8, 1103. [Google Scholar] [CrossRef]
- Silva, L.N.; Zimmer, K.R.; Macedo, A.J.; Trentin, D.S. Plant natural products targeting bacterial virulence factors. Chem. Rev. 2016, 116, 9162–9236. [Google Scholar] [CrossRef]
- Zacchino, S.A.; Butassi, E.; Di Liberto, M.; Raimondi, M.; Postigo, A.; Sortino, M. Plant phenolics and terpenoids as adjuvants of antibacterial and antifungal drugs. Phytomedicine 2017, 37, 27–48. [Google Scholar] [CrossRef]
- El-Mahdy, A.M. Effect of resveratrol on quorum sensing and some virulence characteristics of pseudomonas aeruginosa isolated from manoura university hospitals. J. Microbiol 2017, 46, 98–111. [Google Scholar]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Cha, S.-H.; Cho, I.; Park, S.; Park, Y.; Cho, S.; Park, Y. Antibacterial nanocarriers of resveratrol with gold and silver nanoparticles. Mater. Sci. Eng. C 2016, 58, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.S.; Lee, J.-H.; Cho, M.H.; Lee, J. Red wines and flavonoids diminish Staphylococcus aureus virulence with anti-biofilm and anti-hemolytic activities. Biofouling 2015, 31, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Surh, Y.-J.; Shishodia, S. The Molecular Targets and Therapeutic Uses of Curcumin in Health and Disease; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007; Volume 595. [Google Scholar]
- Araujo, C.; Leon, L. Biological activities of Curcuma longa L. Memórias Do Inst. Oswaldo Cruz 2001, 96, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Zorofchian Moghadamtousi, S.; Abdul Kadir, H.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. BioMed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, X.; Li, W.; Deng, X.; Deng, Y.; Niu, X. Curcumin protects mice from Staphylococcus aureus pneumonia by interfering with the self-assembly process of α-hemolysin. Sci. Rep. 2016, 6, 28254. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Huang, C.; Huang, H.; Zhao, Y.; Khan, M.R.U.; Zhao, H.; Huang, L. Antibacterial mechanism of curcumin: A review. Chem. Biodivers. 2020, 17, e2000171. [Google Scholar] [CrossRef]
- El-Mahdy, A.M.; Alqahtani, M.; Almukainzi, M.; Alghoribi, M.F.; Abdel-Rhman, S.H. Effect of Resveratrol and Curcumin on Gene Expression of Methicillin-Resistant Staphylococcus aureus (MRSA) Toxins. J. Microbiol. Biotechnol. 2024, 34, 141. [Google Scholar] [CrossRef]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K. CLSI methods development and standardization working group best practices for evaluation of antimicrobial susceptibility tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef]
- AS 4276.3.1-1995; Water Microbiology–Heterotrophic Colony Count Methods–Pour Plate Method using Plate Count Agar. Standards Australia Store: Sydney, NSW, Australia, 1995.
- Gupta, R.K.; Setia, S.; Harjai, K. Expression of quorum sensing and virulence factors are interlinked in Pseudomonas aeruginosa: An in vitro approach. Am. J. Biomed. Sci. 2011, 3, 116–125. [Google Scholar] [CrossRef]
- Melo, P.d.C.; Ferreira, L.M.; Nader Filho, A.; Zafalon, L.F.; Vicente, H.I.G.; Souza, V.d. Comparison of methods for the detection of biofilm formation by Staphylococcus aureus isolated from bovine subclinical mastitis. Braz. J. Microbiol. 2013, 44, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Rupp, M.E.; Archer, G.L. Hemagglutination and adherence to plastic by Staphylococcus epidermidis. Infect. Immun. 1992, 60, 4322–4327. [Google Scholar] [CrossRef]
- El-Mowafy, S.A.; Abd El Galil, K.H.; El-Messery, S.M.; Shaaban, M.I. Aspirin is an efficient inhibitor of quorum sensing, virulence and toxins in Pseudomonas aeruginosa. Microb. Pathog. 2014, 74, 25–32. [Google Scholar] [CrossRef] [PubMed]
- El-baz, R.; Rizk, D.E.; Barwa, R.; Hassan, R. Virulence factors profile of Staphylococcus aureus isolated from different clinical sources. J. Microbiol. 2016, 43, 126–144. [Google Scholar]
- Su, Y.; Ma, L.; Wen, Y.; Wang, H.; Zhang, S. Studies of the in vitro antibacterial activities of several polyphenols against clinical isolates of methicillin-resistant Staphylococcus aureus. Molecules 2014, 19, 12630–12639. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.-H.; Joung, D.-K.; Kim, Y.-S.; Kang, O.-H.; Kim, S.-B.; Seo, Y.-S.; Kim, Y.-C.; Lee, D.-S.; Shin, D.-W.; Kweon, K.-T. Synergistic antibacterial effect of curcumin against methicillin-resistant Staphylococcus aureus. Phytomedicine 2013, 20, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Kali, A.; Bhuvaneshwar, D.; Charles, P.M.; Seetha, K.S. Antibacterial synergy of curcumin with antibiotics against biofilm producing clinical bacterial isolates. J. Basic Clin. Pharm. 2016, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Paul, D.; Orwin, M.; Schlievert, P. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar]
- Mulcahy, M.E.; Geoghegan, J.A.; Monk, I.R.; O’Keeffe, K.M.; Walsh, E.J.; Foster, T.J.; McLoughlin, R.M. Nasal colonisation by Staphylococcus aureus depends upon clumping factor B binding to the squamous epithelial cell envelope protein loricrin. PLoS Pathog. 2012, 8, e1003092. [Google Scholar] [CrossRef]
- Craft, K.M.; Nguyen, J.M.; Berg, L.J.; Townsend, S.D. Methicillin-resistant Staphylococcus aureus (MRSA): Antibiotic-resistance and the biofilm phenotype. MedChemComm 2019, 10, 1231–1241. [Google Scholar] [CrossRef]
- Eng, S.-A.; Nathan, S. Curcumin rescues Caenorhabditis elegans from a Burkholderia pseudomallei infection. Front. Microbiol. 2015, 6, 290. [Google Scholar] [CrossRef][Green Version]
- Kebaier, C.; Chamberland, R.R.; Allen, I.C.; Gao, X.; Broglie, P.M.; Hall, J.D.; Jania, C.; Doerschuk, C.M.; Tilley, S.L.; Duncan, J.A. Staphylococcus aureus α-hemolysin mediates virulence in a murine model of severe pneumonia through activation of the NLRP3 inflammasome. J. Infect. Dis. 2012, 205, 807–817. [Google Scholar] [CrossRef]
- Türkyilmaz, S.; Kaya, O. Determination of some virulence factors in Staphylococcus spp. isolated from various clinical samples. Turk. J. Vet. Anim. Sci. 2006, 30, 127–132. [Google Scholar]
- Darmani, H.; Smadi, E.A.; Bataineh, S.M. Blue light emitting diodes enhance the antivirulence effects of Curcumin against Helicobacter pylori. J. Med. Microbiol. 2020, 69, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, A.; Arvidson, S. Variation in extracellular protease production among clinical isolates of Staphylococcus aureus due to different levels of expression of the protease repressor sarA. Infect. Immun. 2002, 70, 4239–4246. [Google Scholar] [CrossRef]
- Saising, J.; Singdam, S.; Ongsakul, M.; Voravuthikunchai, S.P. Lipase, protease, and biofilm as the major virulence factors in staphylococci isolated from acne lesions. Biosci. Trends 2012, 6, 160–164. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rudrappa, T.; Bais, H.P. Curcumin, a known phenolic from Curcuma longa, attenuates the virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models. J. Agric. Food Chem. 2008, 56, 1955–1962. [Google Scholar] [CrossRef]
- Anacarso, I.; Condò, C.; Sabia, C.; Messi, P.; Niederhausern, S.d.; Bondi, M.; Iseppi, R. Antimicrobial resistance and other related virulence factors in staphylococcus spp. isolated from food, environmental and humans in Italy. Univers. J. Microbiol. Res. 2013, 1, 1–9. [Google Scholar] [CrossRef]
- Chung, I.-M.; Subramanian, U.; Thirupathi, P.; Venkidasamy, B.; Samynathan, R.; Gangadhar, B.H.; Rajakumar, G.; Thiruvengadam, M. Resveratrol nanoparticles: A promising therapeutic advancement over native resveratrol. Processes 2020, 8, 458. [Google Scholar] [CrossRef]
- Tian, F.; Conde, J.; Bao, C.; Chen, Y.; Curtin, J.; Cui, D. Gold nanostars for efficient in vitro and in vivo real-time SERS detection and drug delivery via plasmonic-tunable Raman/FTIR imaging. Biomaterials 2016, 106, 87–97. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virulence Profile Pattern | Virulence Factors | VFs | Riyadh | Jeddah | Total |
|---|---|---|---|---|---|
| P1 | Bio, He, HA, Pro, Li | 5 | 0 | 5 | 5 |
| P2 | Bio, He, HA, Li | 4 | 3 | 1 | 4 |
| P3 | Bio, He, Pro, Li | 4 | 3 | 2 | 5 |
| P4 | He, HA, Pro, Li | 4 | 2 | 3 | 5 |
| P5 | Bio, He, HA, Pro | 4 | 1 | 4 | 5 |
| P6 | HA, Pro, Li | 3 | 1 | 0 | 1 |
| P7 | He, HA, Pro | 3 | 2 | 3 | 5 |
| P8 | Bio, He, HA | 3 | 2 | 0 | 2 |
| P9 | He, HA, Li | 3 | 3 | 0 | 3 |
| P10 | Bio, He, Pro | 3 | 1 | 3 | 4 |
| P11 | He, Pro, Li | 3 | 0 | 1 | 1 |
| P12 | Bio, He, Li | 3 | 0 | 2 | 2 |
| P13 | He, HA | 2 | 6 | 0 | 6 |
| P14 | Bio, He | 2 | 1 | 0 | 1 |
| P15 | He, Li | 2 | 0 | 1 | 1 |
| Isolate No. | Control | Trans-Resveratrol | % Reduction | Curcumin | % Reduction |
|---|---|---|---|---|---|
| 17 | 0.14 +/− 0.017 | 0.07 +/− 0.002 | 50.7 | - | - |
| 29 | 0.34 +/− 0.001 | 0.11 +/− 0.001 | 66.8 | 0.11 +/− 0.001 | 68.2 |
| 36 | 0.21 +/− 0.005 | - | - | 0.10 +/− 0.001 | 50.1 |
| 40 | 0.22 +/− 0.001 | 0.08 +/− 0.002 | 65 | 0.10 +/− 0.002 | 54.7 |
| 50 | 0.29 +/− 0.001 | 0.08 +/− 0.001 | 70.7 | 0.07 +/− 0.001 | 72.6 |
| Isolate No. | % Hemoglobin Release | ||||
|---|---|---|---|---|---|
| Control | Trans-Resveratrol | % Reduction | Curcumin | % Reduction | |
| 2 | 53.85 +/− 0.031 | 7.05 +/− 0.065 | 87 | - | - |
| 4 | 24.07 +/− 0.035 | 11.84 +/− 0.046 | 50.8 | 6.09 +/− 0.014 | 74.7 |
| 5 | 7.07 +/− 0.025 | 3.24 +/− 0.072 | 54.2 | - | - |
| 6 | 5.76 +/− 0.02 | 2.24 +/− 0.025 | 61.2 | - | - |
| 7 | 13.14 +/− 0.06 | 3.21 +/− 0.021 | 75.6 | 3.20 +/− 0.025 | 75.6 |
| 9 | 21.17 +/− 0.025 | 3.81 +/− 0.026 | 82 | 5.14 +/− 0.021 | 75.7 |
| 10 | 9.94 +/− 0.032 | 2.55 +/− 0.031 | 74.3 | 3.22 +/− 0.017 | 67.6 |
| 11 | 9.66 +/− 0.119 | 3.81 +/− 0.026 | 60.6 | 3.52 +/− 0.015 | 63.5 |
| 14 | 3.82 +/− 0.025 | 1.25 +/− 0.031 | 67.2 | - | - |
| 15 | 21.47 +/− 0.015 | 3.52 +/− 0.015 | 83.6 | 2.88 +/− 0.01 | 86.6 |
| 17 | 3.52 +/− 0.012 | 1.52 +/− 0.02 | 56.8 | 1.65 +/− 0.02 | 53.2 |
| 18 | 9.26 +/− 0.061 | - | - | 3.21 +/− 0.01 | 65.3 |
| 19 | 21.78 +/− 0.021 | 3.20 +/− 0.015 | 85.3 | 4.81 +/− 0.015 | 78 |
| 20 | 13.47 +/− 0.031 | 3.53 +/− 0.021 | 73.8 | 4.22 +/− 0.072 | 68.7 |
| 21 | 6.73 +/− 0.040 | 2.56 +/− 0.006 | 62 | - | - |
| 22 | 8.04 +/− 0.074 | 3.83 +/− 0.010 | 52.3 | 3.19 +/− 0.010 | 60.3 |
| 23 | 13.48 +/− 0.020 | 3.19 +/− 0.015 | 76.3 | 4.80 +/− 0.010 | 64.4 |
| 24 | 13.10 +/− 0.096 | 2.87 +/− 0.006 | 78 | 4.16 +/− 0.006 | 68.2 |
| 25 | 50.00 +/− 0.10 | 9.29 +/− 0.010 | 81.4 | - | - |
| 27 | 27.55 +/− 0.021 | 4.18 +/− 0.020 | 84.8 | 5.76 +/− 0.006 | 79 |
| 28 | 3.20 +/− 0.015 | 1.12 +/− 0.012 | 64.8 | 1.41 +/− 0.017 | 56 |
| 29 | 77.88 +/− 0.02 | 15.70 +/− 0.015 | 79.8 | 35.24 +/− 0.012 | 54.7 |
| 31 | 87.56 +/− 0.122 | 15.70 +/− 0.010 | 82 | - | - |
| 32 | 81.74 +/− 0.010 | 12.81 +/− 0.010 | 84.3 | - | - |
| 33 | 27.26 +/− 0.032 | 7.05 +/− 0.050 | 74 | 13.22 +/− 0.020 | 51.5 |
| 34 | 14.73 +/− 0.010 | 5.43 +/− 0.012 | 63 | - | - |
| 35 | 34.60 +/− 0.010 | 6.41 +/− 0.006 | 81.5 | 5.11 +/− 0.015 | 85.2 |
| 36 | 61.53 +/− 0.006 | 9.91 +/− 0.015 | 84 | 19.67 +/− 0.058 | 68 |
| 37 | 85.89 +/− 0.010 | 12.49 +/− 0.006 | 85.5 | 24.37 +/− 0.025 | 71.6 |
| 38 | 21.15 +/− 0.010 | 8.99 +/− 0.015 | 57.5 | - | = |
| 39 | 76.29 +/− 0.010 | 6.41 +/− 0.020 | 91.6 | 10.58 +/− 0.015 | 86 |
| 40 | 81.42 +/− 0.006 | 23.69 +/− 0.015 | 71 | - | - |
| 41 | 68.27 +/− 0.026 | 17.62 +/− 0.006 | 74.2 | - | - |
| 42 | 89.73 +/− 0.010 | 21.09 +/− 0.081 | 76.5 | 25.35 +/− 0.132 | 71.7 |
| 44 | 3.20 +/− 0.010 | 1.13 +/− 0.144 | 64.6 | 0.97 +/− 0.010 | 69.7 |
| 45 | 4.18 +/− 0.020 | 1.78 +/− 0.015 | 57.3 | 2.07 +/− 0.026 | 50.5 |
| 46 | 15.38 +/− 0.015 | 4.16 +/− 0.010 | 73 | 3.54 +/− 0.015 | 77 |
| 48 | 17.94 +/− 0.006 | 5.76 +/− 0.010 | 68 | 3.83 +/− 0.006 | 78.6 |
| 49 | 20.86 +/− 0.038 | 4.80 +/− 0.010 | 77 | 6.73 +/− 0.010 | 67.7 |
| 50 | 99.32 +/− 0.015 | 20.33 +/− 0.101 | 79.5 | - | - |
| Isolate No. | Protease Production | ||
|---|---|---|---|
| Control | Trans-Resveratrol | Curcumin | |
| 33 | 67.97 +/− 0.021 | 21.27 +/− 0.007 | - |
| 34 | 60.14 +/− 0.015 | 28.44 +/− 0.015 | - |
| 36 | 49.28 +/− 0.006 | 24.35 +/− 0.010 | 25.27 +/− 0.002 |
| 38 | 50.39 +/− 0.015 | 17.96 +/− 0.058 | 7.65 +/− 0.001 |
| 42 | 60.18 +/− 0.015 | 29.45 +/− 0.010 | - |
| Isolate No. | Lecithinase Production | ||||
|---|---|---|---|---|---|
| Control | Trans-Resveratrol | % Reduction | Curcumin | % Reduction | |
| 1 | 1.37 +/− 0.0006 | 1.18 +/− 0.006 | 14.2 | 1.0.6 +/− 0.001 | 22.7 |
| 9 | 1.35 +/− 0.0006 | 1.28 +/− 0.001 | 4.9 | 1.24 +/− 0.001 | 8.5 |
| 11 | 1.27 +/− 0.001 | 1.13 +/− 0.001 | 11.5 | 0.99 +/− 0.001 | 22 |
| 13 | 1.53 +/− 0.0006 | 1.51 +/− 0.001 | 1.3 | 1.41 +/− 0.002 | 7.9 |
| 14 | 1.55 +/− 0.0006 | 1.40 +/− 0.01 | 9.7 | 1.43 +/− 0.02 | 7.7 |
| 16 | 1.88 +/− 0.0006 | 1.83 +/− 0.006 | 2.4 | 1.63 +/− 0.001 | 13.3 |
| 17 | 1.87 +/− 0.0006 | 1.49 +/− 0.001 | 19.7 | 1.38 +/− 0.001 | 25.9 |
| 21 | 1.82 +/− 0.002 | 1.78 +/− 0.001 | 2.6 | 1.70 +/− 0.001 | 6.5 |
| 22 | 1.85 +/− 0.002 | 1.79 +/− 0.002 | 3.02 | 1.65 +/− 0.002 | 10.6 |
| 23 | 1.87 +/− 0.0006 | 1.72 +/− 0.002 | 7.76 | 1.69 +/− 0.006 | 9.1 |
| 24 | 1.63 +/− 0.015 | 1.47 +/− 0.001 | 10 | 1.48 +/− 0.001 | 9.2 |
| 25 | 2.18 +/− 0.0006 | 1.85 +/− 0.002 | 15.2 | 1.75 +/− 0.001 | 19.8 |
| 28 | 2.03 +/− 0.0006 | 1.89 +/− 0.002 | 2.4 | 1.81 +/− 0.002 | 10.5 |
| 30 | 2.06 +/− 0.012 | 1.79 +/− 0.001 | 13 | 1.77 +/− 0.001 | 14 |
| 34 | 2.21 +/− 0.001 | 1.84 +/− 0.001 | 17.2 | 1.98 +/− 0.002 | 10.7 |
| 35 | 2.17 +/− 0.001 | 1.95 +/− 0.001 | 10.2 | 1.87 +/− 0.002 | 13.7 |
| 36 | 2.21 +/− 0.002 | 2.07 +/− 0.002 | 6.1 | 2.12 +/− 0.002 | 3.7 |
| 37 | 2.17+/− 0.0006 | 2.10 +/− 0.003 | 3 | 2.07 +/− 0.001 | 4.4 |
| 39 | 1.99 +/− 0.003 | 1.81 +/− 0.001 | 9.6 | 1.62 +/− 0.001 | 19 |
| 40 | 2.03 +/− 0.001 | 1.89 +/− 0.001 | 6.8 | 1.96 +/− 0.010 | 3.4 |
| 41 | 2.38 +/− 0.015 | 2.1 +/− 0.002 | 11.5 | 2.04 +/− 0.051 | 14 |
| 43 | 2.16 +/− 0.015 | 1.97 +/− 0.002 | 8.5 | 1.84 +/− 0.001 | 14.9 |
| 44 | 1.92 +/− 0.001 | 1.79 +/− 0.002 | 6.4 | 1.79 +/− 0.002 | 6.2 |
| 47 | 1.92 +/− 0.001 | 1.73 +/− 0.001 | 9.6 | 1.4 +/− 0.001 | 27 |
| 48 | 2.26 +/− 0.002 | 1.67 +/− 0.001 | 26 | 1.66 +/− 0.002 | 26.4 |
| 50 | 1.9 +/− 0.002 | 1.35 +/− 0.002 | 28.8 | 1.69 +/− 0.002 | 10.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqahtani, M.; Almukainzi, M.; Alghoribi, M.F.; El-Mahdy, A.M. Antivirulence Effects of Trans-Resveratrol and Curcumin on Methicillin-Resistant Staphylococcus aureus (MRSA) from Saudi Arabia. Life 2024, 14, 491. https://doi.org/10.3390/life14040491
Alqahtani M, Almukainzi M, Alghoribi MF, El-Mahdy AM. Antivirulence Effects of Trans-Resveratrol and Curcumin on Methicillin-Resistant Staphylococcus aureus (MRSA) from Saudi Arabia. Life. 2024; 14(4):491. https://doi.org/10.3390/life14040491
Chicago/Turabian StyleAlqahtani, Maisa, May Almukainzi, Majed F. Alghoribi, and Areej M. El-Mahdy. 2024. "Antivirulence Effects of Trans-Resveratrol and Curcumin on Methicillin-Resistant Staphylococcus aureus (MRSA) from Saudi Arabia" Life 14, no. 4: 491. https://doi.org/10.3390/life14040491
APA StyleAlqahtani, M., Almukainzi, M., Alghoribi, M. F., & El-Mahdy, A. M. (2024). Antivirulence Effects of Trans-Resveratrol and Curcumin on Methicillin-Resistant Staphylococcus aureus (MRSA) from Saudi Arabia. Life, 14(4), 491. https://doi.org/10.3390/life14040491