
Vitamin D in Melanoma: Potential Role of Cytochrome P450 Enzymes

 , , and
, , and 
Abstract
:1. Background
2. Literature Search Strategy
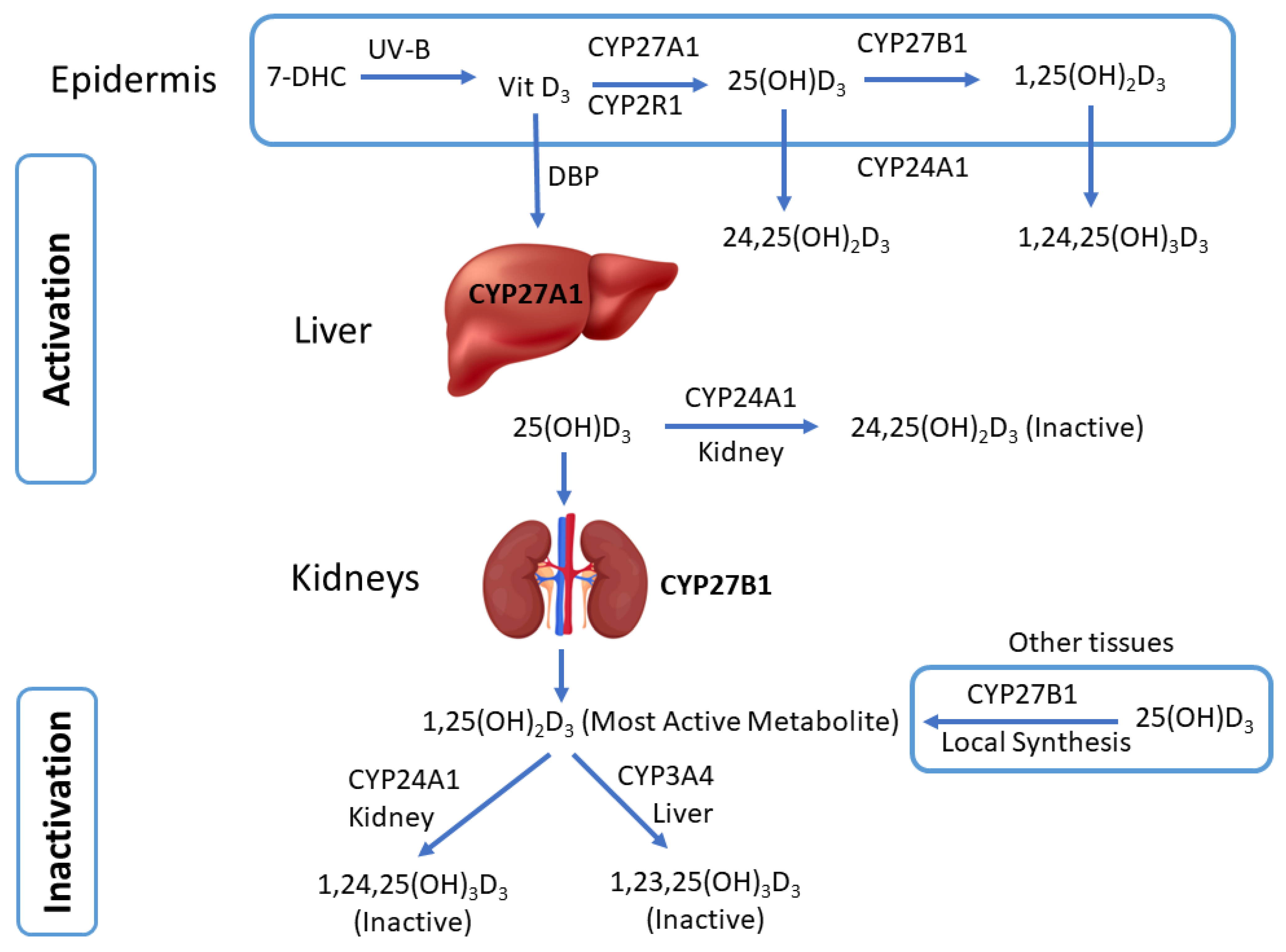
3. Cytochrome P450-Mediated Vitamin D Metabolism
4. Dermal Expression of Vitamin D-Related CYP Enzymes
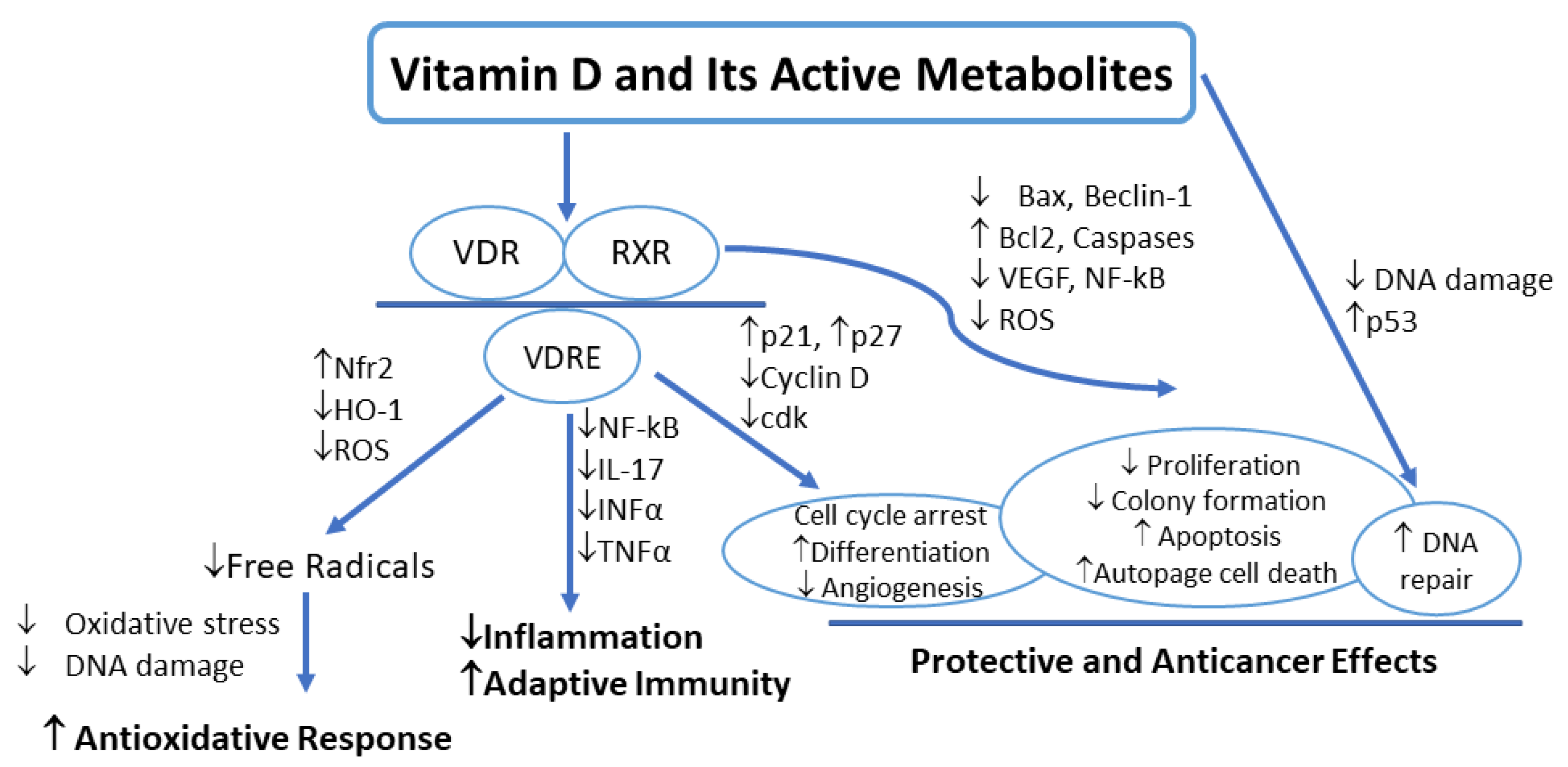
5. Vitamin D Signaling in Melanoma
6. Vitamin D Deficiency and Melanoma

{kind=link}
{kind=link}
| Author | Study Design | Sample Size | Location | Study Population | Duration of Follow Up | Mean Age | Objective | Intervention | Study Findings |
|---|---|---|---|---|---|---|---|---|---|
| Asgari et al., 2009 [11] | Cohort study | Male: 37,382, Female: 40,337 | USA | Men and women from western Washington 50–76 years | 10 years | 62 years (range, 50–76 years | To examine the association between vitamin D intake and melanoma risk | Average intake in µg/day over 10 years from multi- vitamin and individual supplement sources | No association between total intake of vitamin D and melanoma risk |
| De Smedt et al., 2022 [82] | Multicentre randomized double-blind placebo-controlled phase III trial | Male: 78, Female: 77 | Belgium, Hungary | Patients with stage IB to III Cutaneous melanoma (according to the 7th AJCC staging), age 18–80 | NA | Group A 56 (47–66), Group B 54 (45–64) | To investigate the connection between 25(OH)D3 levels, tumor-node-metastasis (TNM) stage and histopathological parameters | 25(OH)D3 plasma levels | Low levels of 25(OH)D3 were associated with a worse outcome in patients with melanoma |
| Liyanage et al., 2020 [6] | Mendelian randomization study | Cases: 12,874 and Controls: 23,203 | Australia | Summary data from a large genome-wide association study | NA | NA | To investigate a causal association between genetically predicted 25(OH)D3 concentrations and melanoma using a Mendelian randomization (MR) approach | Genetically predicted 25(OH)D3 concentrations | Low levels of 25(OH)D3 were not associated with the risk of melanoma [(OR) 1.06, 95% confidence interval (CI) 0.95–1.19] |
| Reichrath et al., 2022 [63] | Prospective, comparative, non-interventional side-by-side study | 83 | Germany | Treatment with CTLA-4i, PD-1i, or BRAFi/MEKi for histologically and clinically confirmed advanced melanoma | 2013–until death | 63.43 years | To investigate whether vitamin D status is predictive of efficacy and safety in patients treated with immune checkpoint or BRAF/MEK inhibitors | Vitamin D levels in plasma | Vitamin D deficiency was associated with poor clinical outcomes in patients treated for metastasized melanoma with BRAF/MEK inhibitors or immunotherapy |
| Befon et al., 2020 [93] | Prospective cohort study | 105, Male: 46, Female: 53 | Greece | Consecutive patients diagnosed with primary invasive CMM of any stage were enrolled. | 42–78 months | ≤40 years: 31 ± 31.31 | Association of serum total 25(OH)D3 levels at melanoma diagnosis with known risk and prognostic factors for CMM | Serum 25-hydroxy vitamin D levels of melanoma patients | Low levels of 25(OH)D3 were found in Greek cohort of melanoma patients vs. healthy controls |
| Stenehjem et al., 2020 [5] | Case-control study | 1416, Case: 708, Control: 708 | Norway | Cases were histologically verified melanomas < 75 years of age | Year 1972–2003 to 31 December 2009 | Cases: 42 (22–67), Control: 42 (22–67 | To study 25(OH)D3 and melanoma risk in the population-based Janus Serum Bank Cohort | Concentrations of 25(OH)D3 were measured during 2016–2017 | Adequate vitamin D levels were associated with a reduced risk of melanoma |
| Fearfield et al., 2019 [74] | Retrospective study | 104 patients | UK | Patients with primary and metastatic melanoma | May 2016 to October 2017. | Range: 23–85 years | To determine levels of 25(OH)D3 in patients with primary and metastatic melanoma | Plasma levels of vitamin D | 80% of patients with melanoma had vitamin D deficiency |
| Timerman et al., 2017 [75] | retrospective, single-center study | 252, Male: 144, Female: 108 | USA | Patients with melanoma and at least one serum 25(OH)D3 measurement within one year after the diagnosis | January 2007 to June 2013 | 55.4 ± 14.7 | The initial serum 25(OH)D3 concentrations and the change in 25(OH)D3 levels were associated with melanoma prognosis | Serum 25(OH)D3 concentrations | Vitamin D deficiency was associated with a worse prognosis in patients with metastatic melanoma |
| Lombardo et al., 2021 [94] | retrospective, single-center study | 154, Male: 78, Female: 76 | Italy | Patients with melanoma with low Vitamin D serum levels. Control groups with a negative melanoma history | 3-year period 2016 to 2019 | 59.7 ± 15.5 | To study the association between vitamin D serum levels and germane prognostic factors/pathological parameters in melanoma | 25(OH)D3 serum levels were measured during 2016–2019 | Vitamin D deficiency is a possible predisposing factor for the development of melanoma |
| Moreno-Arrones et al.,2019 [78] | cross-sectional, single-center study | 204 patients | Spain | Patients with primary invasive melanoma | 2013 to 2017 | - | To investigate the association between vitamin D serum levels and pathological parameters in melanoma | Serum 25(OH)D3 concentrations | There was a significant association between low vitamin D levels and an increased risk of developing aggressive melanoma |
| Bade et al., 2014 [76] | retrospective, single-center study | 324 melanoma patients, 141 healthy controls | Germany | Melanoma patients | February 2000 and April 2004 | 56.3 | To study the association between low vitamin D serum levels and risk for melanoma prognosis | Serum 25(OH)D3 concentrations | Low serum 25(OH)D3 concentrations were associated with poor outcomes in melanoma patients, predicting risk and prognosis of melanoma |
| Gambichler et al., 2013 [77] | Prospective study | 764 patients | Germany | Malignant melanoma | - | - | To study the association between vitamin D serum levels and clinical parameters in melanoma patients | Serum 25(OH)D3 concentrations | Low levels of vitamin D were associated with advanced tumor stage |
| Nurnberg et al., 2009 [79] | Prospective study | 205 Patients, 141 health controls | Germany | Stage 4 melanoma patients | December 1997 to March 2007 | - | To study the association between vitamin D levels and clinical and histopathological parameters among melanoma patients | Serum 25(OH)D3 concentrations | Patients with metastasized melanoma stage 4 had significantly lower vitamin D levels |
| Moro et al., 2022 [80] | Retrospective, observational, longitudinal | 286 patients with history with melanoma; | Spain | Patients with a histological melanoma | Follow-up period of 39.4 months | - | To study the prognostic value of vitamin D serum levels in melanoma patients | Serum 25(OH)D3 concentrations | Low vitamin D levels were associated with ulceration in melanoma |
7. Factors Affecting CYP-Mediated Vitamin D Metabolism and Vitamin D Effects
7.1. Xenobiotics as Inducers of Vitamin D Metabolism
7.2. Xenobiotics as Inhibitors of Vitamin D Metabolism
7.3. Comorbidities
7.4. Polymorphism
8. Therapeutic Prevention of Melanoma by Vitamin D Supplementation and Modulation of CYPs
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Brozyna, A.A.; Zmijewski, M.A.; Jozwicki, W.; Jetten, A.M.; Mason, R.S.; Tuckey, R.C.; Elmets, C.A. Vitamin D signaling and melanoma: Role of vitamin D and its receptors in melanoma progression and management. Lab. Investig. 2017, 97, 706–724. [Google Scholar] [CrossRef]
- Saw, R.P.; Armstrong, B.K.; Mason, R.S.; Morton, R.L.; Shannon, K.F.; Spillane, A.J.; Stretch, J.R.; Thompson, J.F. Adjuvant therapy with high dose vitamin D following primary treatment of melanoma at high risk of recurrence: A placebo controlled randomised phase II trial (ANZMTG 02.09 Mel-D). BMC Cancer 2014, 14, 780. [Google Scholar] [CrossRef] [PubMed]
- Ombra, M.N.; Paliogiannis, P.; Doneddu, V.; Sini, M.C.; Colombino, M.; Rozzo, C.; Stanganelli, I.; Tanda, F.; Cossu, A.; Palmieri, G. Vitamin D status and risk for malignant cutaneous melanoma: Recent advances. Eur. J. Cancer Prev. 2017, 26, 532–541. [Google Scholar] [CrossRef]
- Stenehjem, J.S.; Stoer, N.C.; Ghiasvand, R.; Grimsrud, T.K.; Babigumira, R.; Rees, J.R.; Nilsen, L.T.; Johnsen, B.; Thorsby, P.M.; Veierod, M.B.; et al. Prediagnostic serum 25-hydroxyvitamin D and melanoma risk. Sci. Rep. 2020, 10, 20129. [Google Scholar] [CrossRef]
- Liyanage, U.E.; Law, M.H.; Melanoma Meta-analysis, C.; Barrett, J.H.; Iles, M.M.; MacGregor, S. Is there a causal relationship between vitamin D and melanoma risk? A Mendelian randomization study. Br. J. Dermatol. 2020, 182, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Ben-Eltriki, M.; Deb, S.; Guns, E.S. Calcitriol in Combination Therapy for Prostate Cancer: Pharmacokinetic and Pharmacodynamic Interactions. J. Cancer 2016, 7, 391–407. [Google Scholar] [CrossRef]
- Deb, S.; Reeves, A.A.; Lafortune, S. Simulation of Physicochemical and Pharmacokinetic Properties of Vitamin D(3) and Its Natural Derivatives. Pharmaceuticals 2020, 13, 160. [Google Scholar] [CrossRef]
- Yao, L.; Zhang, N.; Ma, S.; Xie, X.H.; Xu, S.; Xiang, D.; Kang, L.; Chen, M.M.; Wang, P.; Liu, Z. The relationship between vitamin D levels in seasonal variations and Chinese patients with first-episode drug-naive depression. J. Psychosom. Res. 2023, 164, 111079. [Google Scholar] [CrossRef]
- Feng, C.; Song, X.; Chalamaiah, M.; Ren, X.; Wang, M.; Xu, B. Vitamin D Fortification and Its Effect on Athletes’ Physical Improvement: A Mini Review. Foods 2023, 12, 256. [Google Scholar] [CrossRef]
- Asgari, M.M.; Maruti, S.S.; Kushi, L.H.; White, E. A cohort study of vitamin D intake and melanoma risk. J. Investig. Dermatol. 2009, 129, 1675–1680. [Google Scholar] [CrossRef] [PubMed]
- Deeb, K.K.; Trump, D.L.; Johnson, C.S. Vitamin D signalling pathways in cancer: Potential for anticancer therapeutics. Nat. Rev. Cancer 2007, 7, 684–700. [Google Scholar] [CrossRef] [PubMed]
- Crowe, F.L.; Steur, M.; Allen, N.E.; Appleby, P.N.; Travis, R.C.; Key, T.J. Plasma concentrations of 25-hydroxyvitamin D in meat eaters, fish eaters, vegetarians and vegans: Results from the EPIC-Oxford study. Public Health Nutr. 2011, 14, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Caillaud, D.; Cano, N.J. Vitamin D bioavailability: State of the art. Crit. Rev. Food Sci. Nutr. 2015, 55, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed]
- Chiu, N.T.; Tomlinson Guns, E.S.; Adomat, H.; Jia, W.; Deb, S. Identification of human cytochrome P450 enzymes involved in the hepatic and intestinal biotransformation of 20(S)-protopanaxadiol. Biopharm. Drug Dispos. 2014, 35, 104–118. [Google Scholar] [CrossRef]
- Surendradoss, J.; Varghese, A.; Deb, S. Drug Metabolism: Detoxification and Xenobiotic Biotransformation. In Biologically Active Small Molecules: Modern Applications and Therapeutic Perspectives; Mahapatra, D.K., Bharti, S.K., Eds.; Apple Academic Press: Cambridge, MA, USA; Taylor & Francis Group: Abingdon, UK, 2023; pp. 287–332. [Google Scholar] [CrossRef]
- Pascussi, J.M.; Robert, A.; Nguyen, M.; Walrant-Debray, O.; Garabedian, M.; Martin, P.; Pineau, T.; Saric, J.; Navarro, F.; Maurel, P.; et al. Possible involvement of pregnane X receptor-enhanced CYP24 expression in drug-induced osteomalacia. J. Clin. Investig. 2005, 115, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Schuster, I. Cytochromes P450 are essential players in the vitamin D signaling system. Biochim. Biophys. Acta 2011, 1814, 186–199. [Google Scholar] [CrossRef]
- Deb, S.; Pandey, M.; Adomat, H.; Guns, E.S. Cytochrome P450 3A-mediated microsomal biotransformation of 1alpha,25-dihydroxyvitamin D3 in mouse and human liver: Drug-related induction and inhibition of catabolism. Drug Metab. Dispos. 2012, 40, 907–918. [Google Scholar] [CrossRef]
- Zhou, C.; Assem, M.; Tay, J.C.; Watkins, P.B.; Blumberg, B.; Schuetz, E.G.; Thummel, K.E. Steroid and xenobiotic receptor and vitamin D receptor crosstalk mediates CYP24 expression and drug-induced osteomalacia. J. Clin. Investig. 2006, 116, 1703–1712. [Google Scholar] [CrossRef]
- Feldman, D.; Krishnan, A.V.; Swami, S.; Giovannucci, E.; Feldman, B.J. The role of vitamin D in reducing cancer risk and progression. Nat. Rev. Cancer 2014, 14, 342–357. [Google Scholar] [CrossRef]
- Garland, C.F.; Gorham, E.D.; Mohr, S.B.; Garland, F.C. Vitamin D for cancer prevention: Global perspective. Ann. Epidemiol. 2009, 19, 468–483. [Google Scholar] [CrossRef] [PubMed]
- Lappe, J.M.; Travers-Gustafson, D.; Davies, K.M.; Recker, R.R.; Heaney, R.P. Vitamin D and calcium supplementation reduces cancer risk: Results of a randomized trial. Am. J. Clin. Nutr. 2007, 85, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Osborne, J.E.; Hutchinson, P.E. Vitamin D and systemic cancer: Is this relevant to malignant melanoma? Br. J. Dermatol. 2002, 147, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.A.; Hoffman, R.M.; Slominski, A.T. Relevance of Vitamin D in Melanoma Development, Progression and Therapy. Anticancer. Res. 2020, 40, 473–489. [Google Scholar] [CrossRef] [PubMed]
- Galus, L.; Michalak, M.; Lorenz, M.; Stoinska-Swiniarek, R.; Tusien Malecka, D.; Galus, A.; Kolenda, T.; Leporowska, E.; Mackiewicz, J. Vitamin D supplementation increases objective response rate and prolongs progression-free time in patients with advanced melanoma undergoing anti-PD-1 therapy. Cancer 2023, 129, 2047–2055. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, A.; Zaucha, R.; Krol, O.; Zmijewski, M.A. Vitamin D Modulates the Response of Patient-Derived Metastatic Melanoma Cells to Anticancer Drugs. Int. J. Mol. Sci. 2023, 24, 8037. [Google Scholar] [CrossRef]
- Slominski, A.T.; Li, W.; Kim, T.K.; Semak, I.; Wang, J.; Zjawiony, J.K.; Tuckey, R.C. Novel activities of CYP11A1 and their potential physiological significance. J. Steroid Biochem. Mol. Biol. 2015, 151, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, C.; Velleuer, E. Vitamin D and the risk for cancer: A molecular analysis. Biochem. Pharmacol. 2022, 196, 114735. [Google Scholar] [CrossRef]
- Hutchinson, P.E.; Pringle, J.H. Consideration of possible effects of vitamin D on established cancer, with reference to malignant melanoma. Pigment. Cell Melanoma Res. 2022, 35, 408–424. [Google Scholar] [CrossRef]
- Kim, H.; Giovannucci, E. Vitamin D Status and Cancer Incidence, Survival, and Mortality. Adv. Exp. Med. Biol. 2020, 1268, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Kuznia, S.; Zhu, A.; Akutsu, T.; Buring, J.E.; Camargo, C.A., Jr.; Cook, N.R.; Chen, L.J.; Cheng, T.D.; Hantunen, S.; Lee, I.M.; et al. Efficacy of vitamin D(3) supplementation on cancer mortality: Systematic review and individual patient data meta-analysis of randomised controlled trials. Ageing Res. Rev. 2023, 87, 101923. [Google Scholar] [CrossRef] [PubMed]
- Vanhevel, J.; Verlinden, L.; Doms, S.; Wildiers, H.; Verstuyf, A. The role of vitamin D in breast cancer risk and progression. Endocr. Relat. Cancer 2022, 29, R33–R55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fang, F.; Tang, J.; Jia, L.; Feng, Y.; Xu, P.; Faramand, A. Association between vitamin D supplementation and mortality: Systematic review and meta-analysis. BMJ 2019, 366, l4673. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, F.; Saba, A.; Zucchi, R. An Update on Vitamin D Metabolism. Int. J. Mol. Sci. 2020, 21, 6573. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Prosser, D.E.; Kaufmann, M. Cytochrome P450-mediated metabolism of vitamin D. J. Lipid Res. 2014, 55, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Bolerazska, B.; Rabajdova, M.; Spakova, I.; Marekova, M. Current knowledge on the active form of Vitamin D synthesized in the skin and its effects on malignant melanoma. Neoplasma 2017, 64, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.L.; Carpenter, E.L.; Slominski, A.T.; Indra, A.K. The Role of the Vitamin D Receptor in the Pathogenesis, Prognosis, and Treatment of Cutaneous Melanoma. Front. Oncol. 2021, 11, 743667. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D. The vitamin D receptor: A tumor suppressor in skin. Adv. Exp. Med. Biol. 2014, 810, 282–302. [Google Scholar] [CrossRef]
- Thummel, K.E.; Brimer, C.; Yasuda, K.; Thottassery, J.; Senn, T.; Lin, Y.; Ishizuka, H.; Kharasch, E.; Schuetz, J.; Schuetz, E. Transcriptional control of intestinal cytochrome P-4503A by 1alpha,25-dihydroxy vitamin D3. Mol. Pharmacol. 2001, 60, 1399–1406. [Google Scholar] [CrossRef]
- Reddy, K.R.; Deepika, M.L.; Supriya, K.; Latha, K.P.; Rao, S.S.; Rani, V.U.; Jahan, P. CYP11A1 microsatellite (tttta)n polymorphism in PCOS women from South India. J. Assist. Reprod. Genet. 2014, 31, 857–863. [Google Scholar] [CrossRef]
- Slominski, A.; Kim, T.K.; Zmijewski, M.A.; Janjetovic, Z.; Li, W.; Chen, J.; Kusniatsova, E.I.; Semak, I.; Postlethwaite, A.; Miller, D.D.; et al. Novel vitamin D photoproducts and their precursors in the skin. Dermatoendocrinology 2013, 5, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Nemazannikova, N.; Blatch, G.L.; Dass, C.R.; Sinclair, R.; Apostolopoulos, V. Vitamin D enzymes (CYP27A1, CYP27B1, and CYP24A1) and receptor expression in non-melanoma skin cancer. Acta Biochim. Biophys. Sin. 2019, 51, 444–447. [Google Scholar] [CrossRef] [PubMed]
- Pavanello, S.; Angelici, L.; Hoxha, M.; Cantone, L.; Campisi, M.; Tirelli, A.S.; Vigna, L.; Pesatori, A.C.; Bollati, V. Sterol 27-Hydroxylase Polymorphism Significantly Associates with Shorter Telomere, Higher Cardiovascular and Type-2 Diabetes Risk in Obese Subjects. Front. Endocrinol. 2018, 9, 309. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; DeLuca, H.F. Vitamin D 25-hydroxylase-Four decades of searching, are we there yet? Arch. Biochem. Biophys. 2012, 523, 30–36. [Google Scholar] [CrossRef]
- Dong, A.N.; Tan, B.H.; Pan, Y.; Ong, C.E. The CYP2R1 Enzyme: Structure, Function, Enzymatic Properties and Genetic Polymorphism. J. Pharm. Pharm. Sci. 2021, 24, 94–112. [Google Scholar] [CrossRef]
- Ellfolk, M.; Norlin, M.; Gyllensten, K.; Wikvall, K. Regulation of human vitamin D(3) 25-hydroxylases in dermal fibroblasts and prostate cancer LNCaP cells. Mol. Pharmacol. 2009, 75, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Schuetz, E.G.; Xu, Y.; Thummel, K.E. Interplay between vitamin D and the drug metabolizing enzyme CYP3A4. J. Steroid Biochem. Mol. Biol. 2013, 136, 54–58. [Google Scholar] [CrossRef]
- Friedrich, M.; Diesing, D.; Cordes, T.; Fischer, D.; Becker, S.; Chen, T.C.; Flanagan, J.N.; Tangpricha, V.; Gherson, I.; Holick, M.F.; et al. Analysis of 25-hydroxyvitamin D3-1alpha-hydroxylase in normal and malignant breast tissue. Anticancer. Res. 2006, 26, 2615–2620. [Google Scholar]
- Hewison, M.; Zehnder, D.; Chakraverty, R.; Adams, J.S. Vitamin D and barrier function: A novel role for extra-renal 1 alpha-hydroxylase. Mol. Cell Endocrinol. 2004, 215, 31–38. [Google Scholar] [CrossRef]
- King, A.N.; Beer, D.G.; Christensen, P.J.; Simpson, R.U.; Ramnath, N. The vitamin D/CYP24A1 story in cancer. Anticancer. Agents Med. Chem. 2010, 10, 213–224. [Google Scholar] [CrossRef]
- Schafer, A.; Emmert, S.; Kruppa, J.; Schubert, S.; Tzvetkov, M.; Mossner, R.; Reich, K.; Berking, C.; Volkenandt, M.; Pfohler, C.; et al. No association of vitamin D metabolism-related polymorphisms and melanoma risk as well as melanoma prognosis: A case-control study. Arch. Dermatol. Res. 2012, 304, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Prosser, D.E.; Jones, G. Enzymes involved in the activation and inactivation of vitamin D. Trends Biochem. Sci. 2004, 29, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.A.; Jochymski, C.; Janjetovic, Z.; Jozwicki, W.; Tuckey, R.C.; Slominski, A.T. CYP24A1 expression inversely correlates with melanoma progression: Clinic-pathological studies. Int. J. Mol. Sci. 2014, 15, 19000–19017. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, T.; Xu, Y.; Mohutsky, M.A.; Alberts, J.; Hadden, C.; Kalhorn, T.F.; Isoherranen, N.; Shuhart, M.C.; Thummel, K.E. Identification of human UDP-glucuronosyltransferases catalyzing hepatic 1alpha,25-dihydroxyvitamin D3 conjugation. Biochem. Pharmacol. 2008, 75, 1240–1250. [Google Scholar] [CrossRef] [PubMed]
- Kurogi, K.; Sakakibara, Y.; Suiko, M.; Liu, M.C. Sulfation of vitamin D(3) -related compounds-identification and characterization of the responsible human cytosolic sulfotransferases. FEBS Lett. 2017, 591, 2417–2425. [Google Scholar] [CrossRef] [PubMed]
- Wasiewicz, T.; Piotrowska, A.; Wierzbicka, J.; Slominski, A.T.; Zmijewski, M.A. Antiproliferative Activity of Non-Calcemic Vitamin D Analogs on Human Melanoma Lines in Relation to VDR and PDIA3 Receptors. Int. J. Mol. Sci. 2018, 19, 2583. [Google Scholar] [CrossRef] [PubMed]
- Seifert, M.; Rech, M.; Meineke, V.; Tilgen, W.; Reichrath, J. Differential biological effects of 1,25-dihydroxyVitamin D3 on melanoma cell lines in vitro. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Reichrath, J.; Rech, M.; Moeini, M.; Meese, E.; Tilgen, W.; Seifert, M. In vitro comparison of the vitamin D endocrine system in 1,25(OH)2D3-responsive and -resistant melanoma cells. Cancer Biol. Ther. 2007, 6, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Radermacher, J.; Diesel, B.; Seifert, M.; Tilgen, W.; Reichrath, J.; Fischer, U.; Meese, E. Expression analysis of CYP27B1 in tumor biopsies and cell cultures. Anticancer Res. 2006, 26, 2683–2686. [Google Scholar]
- Slominski, A.T.; Brozyna, A.A.; Skobowiat, C.; Zmijewski, M.A.; Kim, T.K.; Janjetovic, Z.; Oak, A.S.; Jozwicki, W.; Jetten, A.M.; Mason, R.S.; et al. On the role of classical and novel forms of vitamin D in melanoma progression and management. J. Steroid Biochem. Mol. Biol. 2018, 177, 159–170. [Google Scholar] [CrossRef]
- Reichrath, J.; Biersack, F.; Wagenpfeil, S.; Schope, J.; Pfohler, C.; Saternus, R.; Vogt, T. Low Vitamin D Status Predicts Poor Clinical Outcome in Advanced Melanoma Treated with Immune Checkpoint or BRAF/MEK Inhibitors: A Prospective Non-Interventional Side-by-Side Analysis. Front. Oncol. 2022, 12, 839816. [Google Scholar] [CrossRef]
- Evans, S.R.; Houghton, A.M.; Schumaker, L.; Brenner, R.V.; Buras, R.R.; Davoodi, F.; Nauta, R.J.; Shabahang, M. Vitamin D receptor and growth inhibition by 1,25-dihydroxyvitamin D3 in human malignant melanoma cell lines. J. Surg. Res. 1996, 61, 127–133. [Google Scholar] [CrossRef]
- Sutedja, E.K.; Amarassaphira, D.; Goenawan, H.; Susanti Pratiwi, Y.; Sylviana, N.; Setiabudiawan, B.; Suwarsa, O.; Tina Dewi Judistiani, R.; Supratman, U.; Lesmana, R. Calcitriol Inhibits Proliferation and Potentially Induces Apoptosis in B16-F10 Cells. Med. Sci. Monit. Basic. Res. 2022, 28, e935139. [Google Scholar] [CrossRef]
- Danielsson, C.; Fehsel, K.; Polly, P.; Carlberg, C. Differential apoptotic response of human melanoma cells to 1 alpha,25-dihydroxyvitamin D3 and its analogues. Cell Death Differ. 1998, 5, 946–952. [Google Scholar] [CrossRef]
- Harant, H.; Andrew, P.J.; Reddy, G.S.; Foglar, E.; Lindley, I.J. 1alpha,25-dihydroxyvitamin D3 and a variety of its natural metabolites transcriptionally repress nuclear-factor-kappaB-mediated interleukin-8 gene expression. Eur. J. Biochem. 1997, 250, 63–71. [Google Scholar] [CrossRef]
- Sauer, B.; Ruwisch, L.; Kleuser, B. Antiapoptotic action of 1alpha,25-dihydroxyvitamin D3 in primary human melanocytes. Melanoma Res. 2003, 13, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Philips, N.; Samuel, P.; Keller, T.; Alharbi, A.; Alshalan, S.; Shamlan, S.A. Beneficial Regulation of Cellular Oxidative Stress Effects, and Expression of Inflammatory, Angiogenic, and the Extracellular Matrix Remodeling Proteins by 1alpha,25-Dihydroxyvitamin D3 in a Melanoma Cell Line. Molecules 2020, 25, 1164. [Google Scholar] [CrossRef] [PubMed]
- Markiewicz, A.; Brozyna, A.A.; Podgorska, E.; Elas, M.; Urbanska, K.; Jetten, A.M.; Slominski, A.T.; Jozwicki, W.; Orlowska-Heitzman, J.; Dyduch, G.; et al. Vitamin D receptors (VDR), hydroxylases CYP27B1 and CYP24A1 and retinoid-related orphan receptors (ROR) level in human uveal tract and ocular melanoma with different melanization levels. Sci. Rep. 2019, 9, 9142. [Google Scholar] [CrossRef]
- Brozyna, A.A.; Jozwicki, W.; Janjetovic, Z.; Slominski, A.T. Expression of the vitamin D-activating enzyme 1alpha-hydroxylase (CYP27B1) decreases during melanoma progression. Hum. Pathol. 2013, 44, 374–387. [Google Scholar] [CrossRef]
- Reichrath, J.; Zouboulis, C.C.; Vogt, T.; Holick, M.F. Targeting the vitamin D endocrine system (VDES) for the management of inflammatory and malignant skin diseases: An historical view and outlook. Rev. Endocr. Metab. Disord. 2016, 17, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Reichrath, J.; Saternus, R.; Vogt, T. Endocrine actions of vitamin D in skin: Relevance for photocarcinogenesis of non-melanoma skin cancer, and beyond. Mol. Cell Endocrinol. 2017, 453, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Fearfield, L.; Nobbs, J.; Petruckevitch, A.; Harland, C. Severe vitamin D deficiency associated with BRAF-mutated melanoma. Br. J. Dermatol. 2019, 181, 1343. [Google Scholar] [CrossRef] [PubMed]
- Timerman, D.; McEnery-Stonelake, M.; Joyce, C.J.; Nambudiri, V.E.; Hodi, F.S.; Claus, E.B.; Ibrahim, N.; Lin, J.Y. Vitamin D deficiency is associated with a worse prognosis in metastatic melanoma. Oncotarget 2017, 8, 6873–6882. [Google Scholar] [CrossRef] [PubMed]
- Bade, B.; Zdebik, A.; Wagenpfeil, S.; Graber, S.; Geisel, J.; Vogt, T.; Reichrath, J. Low serum 25-hydroxyvitamin d concentrations are associated with increased risk for melanoma and unfavourable prognosis. PLoS ONE 2014, 9, e112863. [Google Scholar] [CrossRef] [PubMed]
- Gambichler, T.; Bindsteiner, M.; Hoxtermann, S.; Kreuter, A. Serum 25-hydroxyvitamin D serum levels in a large German cohort of patients with melanoma. Br. J. Dermatol. 2013, 168, 625–628. [Google Scholar] [CrossRef]
- Moreno-Arrones, O.M.; Zegeer, J.; Gerbo, M.; Manrique-Silva, E.; Requena, C.; Traves, V.; Nagore, E. Decreased vitamin D serum levels at melanoma diagnosis are associated with tumor ulceration and high tumor mitotic rate. Melanoma Res. 2019, 29, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Nurnberg, B.; Graber, S.; Gartner, B.; Geisel, J.; Pfohler, C.; Schadendorf, D.; Tilgen, W.; Reichrath, J. Reduced serum 25-hydroxyvitamin D levels in stage IV melanoma patients. Anticancer. Res. 2009, 29, 3669–3674. [Google Scholar]
- Moro, R.; Sanchez-Silva, A.; Aguerralde-Martin, M.; Gonzalez-Cuevas, R.; Peruilh-Bagolini, L.; Traves, V.; Manrique-Silva, E.; Requena, C.; Nagore, E. Prognostic Value of Vitamin D Serum Levels in Cutaneous Melanoma. Actas Dermosifiliogr. 2022, 113, 347–353. [Google Scholar] [CrossRef]
- Song, Y.; Lu, H.; Cheng, Y. To identify the association between dietary vitamin D intake and serum levels and risk or prognostic factors for melanoma-systematic review and meta-analysis. BMJ Open 2022, 12, e052442. [Google Scholar] [CrossRef]
- De Smedt, J.; Van Kelst, S.; Janssen, L.; Marasigan, V.; Boecxstaens, V.; Stas, M.; Vanderschueren, D.; Guler, I.; Bogaerts, K.; Vandenberghe, K.; et al. Determinants of 25-hydroxyvitamin D Status in a Cutaneous Melanoma Population. Acta Derm. Venereol. 2022, 102, adv00692. [Google Scholar] [CrossRef] [PubMed]
- American Association of Endocrinology. Vitamin D Deficiency; American Association of Endocrinology: Jacksonville, FL, USA, 2019.
- Institute of Medicine (US) Committee to Review Dietary Reference Intakes for Vitamin D and Calcium. Dietary Reference Intakes for Calcium and Vitamin D; Ross, A.C., Taylor, C.L., Yaktine, A.L., Del Valle, H.B., Eds.; Institute of Medicine (US) Committee to Review Dietary Reference Intakes for Vitamin D and Calcium: Washington, DC, USA, 2011. [CrossRef]
- Office of Dietary Supplements, National Institute of Health. Vitamin D Fact Sheet for Health Professionals. Available online: https://ods.od.nih.gov/factsheets/VitaminD-HealthProfessional/ (accessed on 24 February 2024).
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M. Guidelines for preventing and treating vitamin D deficiency and insufficiency revisited. J. Clin. Endocrinol. Metab. 2012, 97, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M.; Endocrine, S. Evaluation, treatment, and prevention of vitamin D deficiency: An Endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [PubMed]
- Amrein, K.; Scherkl, M.; Hoffmann, M.; Neuwersch-Sommeregger, S.; Kostenberger, M.; Tmava Berisha, A.; Martucci, G.; Pilz, S.; Malle, O. Vitamin D deficiency 2.0: An update on the current status worldwide. Eur. J. Clin. Nutr. 2020, 74, 1498–1513. [Google Scholar] [CrossRef] [PubMed]
- National Institute for Health and Care Excellence. Vitamin D: Supplement Use in Specific Population Groups; National Institute for Health and Care Excellence: London, UK, 2014.
- Berger, M.M.; Shenkin, A.; Schweinlin, A.; Amrein, K.; Augsburger, M.; Biesalski, H.K.; Bischoff, S.C.; Casaer, M.P.; Gundogan, K.; Lepp, H.L.; et al. ESPEN micronutrient guideline. Clin. Nutr. 2022, 41, 1357–1424. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, A.; Wierzbicka, J.; Kwiatkowska, K.; Chodynski, M.; Kutner, A.; Zmijewski, M.A. Antiproliferative activity of side-chain truncated vitamin D analogs (PRI-1203 and PRI-1204) against human malignant melanoma cell lines. Eur. J. Pharmacol. 2020, 881, 173170. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.Y.; Kuo, C.Y.; Huang, Y.C. The association between serum vitamin D level and risk and prognosis of melanoma: A systematic review and meta-analysis. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1722–1729. [Google Scholar] [CrossRef] [PubMed]
- Befon, A.; Katoulis, A.C.; Georgala, S.; Katsampas, A.; Chardalia, V.; Melpidou, A.; Tzanetakou, V.; Chasapi, V.; Polydorou, D.; Desinioti, C.; et al. Serum Total 25-Hydroxyvitamin D Levels in Patients with Cutaneous Malignant Melanoma: A Case-Control Study in a Low-Risk Southern European Population. Dermatol. Pr. Pract. Concept. 2020, 10, e2020010. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, M.; Vigezzi, A.; Ietto, G.; Franchi, C.; Iori, V.; Masci, F.; Scorza, A.; Macchi, S.; Iovino, D.; Parise, C.; et al. Role of vitamin D serum levels in prevention of primary and recurrent melanoma. Sci. Rep. 2021, 11, 5815. [Google Scholar] [CrossRef]
- Raymond-Lezman, J.R.; Riskin, S.I. Benefits and Risks of Sun Exposure to Maintain Adequate Vitamin D Levels. Cureus 2023, 15, e38578. [Google Scholar] [CrossRef]
- Binkley, N.; Novotny, R.; Krueger, D.; Kawahara, T.; Daida, Y.G.; Lensmeyer, G.; Hollis, B.W.; Drezner, M.K. Low vitamin D status despite abundant sun exposure. J. Clin. Endocrinol. Metab. 2007, 92, 2130–2135. [Google Scholar] [CrossRef]
- Boaventura, B.C.B.; Cembranel, F. Protective effect of vitamin D on oxidative stress in elderly people. In Aging, 2nd ed.; Victor, R., Preedy, V.B.P., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 337–343. [Google Scholar] [CrossRef]
- Omar, H.S.; Taha, F.M.; Fouad, S.; Ibrahim, F.A.; El Gendy, A.; Bassyouni, I.H.; El-Shazly, R. The association between vitamin D levels and oxidative stress markers in Egyptian Behcet’s disease patients. Orphanet J. Rare Dis. 2022, 17, 264. [Google Scholar] [CrossRef]
- Gu, J.C.; Wu, Y.G.; Huang, W.G.; Fan, X.J.; Chen, X.H.; Zhou, B.; Lin, Z.J.; Feng, X.L. Effect of vitamin D on oxidative stress and serum inflammatory factors in the patients with type 2 diabetes. J. Clin. Lab. Anal. 2022, 36, e24430. [Google Scholar] [CrossRef]
- Renke, G.; Starling-Soares, B.; Baesso, T.; Petronio, R.; Aguiar, D.; Paes, R. Effects of Vitamin D on Cardiovascular Risk and Oxidative Stress. Nutrients 2023, 15, 769. [Google Scholar] [CrossRef]
- Bisevac, J.P.; Djukic, M.; Stanojevic, I.; Stevanovic, I.; Mijuskovic, Z.; Djuric, A.; Gobeljic, B.; Banovic, T.; Vojvodic, D. Association Between Oxidative Stress and Melanoma Progression. J. Med. Biochem. 2018, 37, 12–20. [Google Scholar] [CrossRef]
- Ghoneum, A.; Abdulfattah, A.Y.; Warren, B.O.; Shu, J.; Said, N. Redox Homeostasis and Metabolism in Cancer: A Complex Mechanism and Potential Targeted Therapeutics. Int. J. Mol. Sci. 2020, 21, 3100. [Google Scholar] [CrossRef]
- Karampinis, E.; Nechalioti, P.-M.; Georgopoulou, K.E.; Goniotakis, G.; Roussaki Schulze, A.V.; Zafiriou, E.; Kouretas, D. Systemic Oxidative Stress Parameters in Skin Cancer Patients and Patients with Benign Lesions. Stresses 2023, 3, 785–812. [Google Scholar] [CrossRef]
- Karampinis, E.; Aloizou, A.M.; Zafiriou, E.; Bargiota, A.; Skaperda, Z.; Kouretas, D.; Roussaki-Schulze, A.V. Non-Melanoma Skin Cancer and Vitamin D: The “Lost Sunlight” Paradox and the Oxidative Stress Explanation. Antioxidants 2023, 12, 1107. [Google Scholar] [CrossRef]
- Muindi, J.R.; Yu, W.D.; Ma, Y.; Engler, K.L.; Kong, R.X.; Trump, D.L.; Johnson, C.S. CYP24A1 inhibition enhances the antitumor activity of calcitriol. Endocrinology 2010, 151, 4301–4312. [Google Scholar] [CrossRef]
- Drocourt, L.; Ourlin, J.C.; Pascussi, J.M.; Maurel, P.; Vilarem, M.J. Expression of CYP3A4, CYP2B6, and CYP2C9 is regulated by the vitamin D receptor pathway in primary human hepatocytes. J. Biol. Chem. 2002, 277, 25125–25132. [Google Scholar] [CrossRef]
- Schmiedlin-Ren, P.; Thummel, K.E.; Fisher, J.M.; Paine, M.F.; Watkins, P.B. Induction of CYP3A4 by 1 alpha,25-dihydroxyvitamin D3 is human cell line-specific and is unlikely to involve pregnane X receptor. Drug Metab. Dispos. 2001, 29, 1446–1453. [Google Scholar]
- Xie, W.; Barwick, J.L.; Simon, C.M.; Pierce, A.M.; Safe, S.; Blumberg, B.; Guzelian, P.S.; Evans, R.M. Reciprocal activation of xenobiotic response genes by nuclear receptors SXR/PXR and CAR. Genes. Dev. 2000, 14, 3014–3023. [Google Scholar] [CrossRef]
- Norlin, M.; Lundqvist, J.; Ellfolk, M.; Hellstrom Pigg, M.; Gustafsson, J.; Wikvall, K. Drug-Mediated Gene Regulation of Vitamin D(3) Metabolism in Primary Human Dermal Fibroblasts. Basic. Clin. Pharmacol. Toxicol. 2017, 120, 59–63. [Google Scholar] [CrossRef]
- Grober, U.; Kisters, K. Influence of drugs on vitamin D and calcium metabolism. Dermatoendocrinology 2012, 4, 158–166. [Google Scholar] [CrossRef]
- Sheng, L.; Xue, Y.; He, X.; Zhu, Y.; Li, H.; Wu, Y.; Dang, R.; Tang, M.; Jiang, P. Effects of repeated administration of rifampicin and isoniazid on vitamin D metabolism in mice. Steroids 2015, 104, 203–207. [Google Scholar] [CrossRef]
- Kasarla, S.S.; Garikapati, V.; Kumar, Y.; Dodoala, S. Interplay of Vitamin D and CYP3A4 Polymorphisms in Endocrine Disorders and Cancer. Endocrinol. Metab. 2022, 37, 392–407. [Google Scholar] [CrossRef]
- Robien, K.; Oppeneer, S.J.; Kelly, J.A.; Hamilton-Reeves, J.M. Drug-vitamin D interactions: A systematic review of the literature. Nutr. Clin. Pr. Pract. 2013, 28, 194–208. [Google Scholar] [CrossRef]
- Wang, Z.; Lin, Y.S.; Dickmann, L.J.; Poulton, E.J.; Eaton, D.L.; Lampe, J.W.; Shen, D.D.; Davis, C.L.; Shuhart, M.C.; Thummel, K.E. Enhancement of hepatic 4-hydroxylation of 25-hydroxyvitamin D3 through CYP3A4 induction in vitro and in vivo: Implications for drug-induced osteomalacia. J. Bone Min. Min. Res. 2013, 28, 1101–1116. [Google Scholar] [CrossRef]
- Wang, Z.; Lin, Y.S.; Zheng, X.E.; Senn, T.; Hashizume, T.; Scian, M.; Dickmann, L.J.; Nelson, S.D.; Baillie, T.A.; Hebert, M.F.; et al. An inducible cytochrome P450 3A4-dependent vitamin D catabolic pathway. Mol. Pharmacol. 2012, 81, 498–509. [Google Scholar] [CrossRef]
- Xu, Y.; Hashizume, T.; Shuhart, M.C.; Davis, C.L.; Nelson, W.L.; Sakaki, T.; Kalhorn, T.F.; Watkins, P.B.; Schuetz, E.G.; Thummel, K.E. Intestinal and hepatic CYP3A4 catalyze hydroxylation of 1alpha,25-dihydroxyvitamin D(3): Implications for drug-induced osteomalacia. Mol. Pharmacol. 2006, 69, 56–65. [Google Scholar] [CrossRef]
- Wakeman, M. A Literature Review of the Potential Impact of Medication on Vitamin D Status. Risk Manag. Heal. Healthc. Policy 2021, 14, 3357–3381. [Google Scholar] [CrossRef] [PubMed]
- Ben-Eltriki, M.; Hopefl, R.; Wright, J.M.; Deb, S. Association between Vitamin D Status and Risk of Developing Severe COVID-19 Infection: A Meta-Analysis of Observational Studies. J. Am. Nutr. Assoc. 2022, 41, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Hopefl, R.; Ben-Eltriki, M.; Deb, S. Association Between Vitamin D Levels and Inflammatory Markers in COVID-19 Patients: A Meta-Analysis of Observational Studies. J. Pharm. Pharm. Sci. 2022, 25, 124–136. [Google Scholar] [CrossRef]
- Grenet, O.; Bobadilla, M.; Chibout, S.D.; Steiner, S. Evidence for the impairment of the vitamin D activation pathway by cyclosporine A. Biochem. Pharmacol. 2000, 59, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, M.; Vidal, M.; Arcidiacono, M.V.; Tebas, P.; Yarasheski, K.E.; Dusso, A.S. HIV-protease inhibitors impair vitamin D bioactivation to 1,25-dihydroxyvitamin D. AIDS 2003, 17, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Ly, L.H.; Zhao, X.Y.; Holloway, L.; Feldman, D. Liarozole acts synergistically with 1alpha,25-dihydroxyvitamin D3 to inhibit growth of DU 145 human prostate cancer cells by blocking 24-hydroxylase activity. Endocrinology 1999, 140, 2071–2076. [Google Scholar] [CrossRef] [PubMed]
- Yee, S.W.; Campbell, M.J.; Simons, C. Inhibition of Vitamin D3 metabolism enhances VDR signalling in androgen-independent prostate cancer cells. J. Steroid Biochem. Mol. Biol. 2006, 98, 228–235. [Google Scholar] [CrossRef]
- Ben-Eltriki, M.; Hassona, M.; Meckling, G.; Adomat, H.; Deb, S.; Tomlinson Guns, E.S. Pharmacokinetic interaction of calcitriol with 20(S)-protopanaxadiol in mice: Determined by LC/MS analysis. Eur. J. Pharm. Sci. 2019, 130, 173–180. [Google Scholar] [CrossRef]
- Deb, S.; Chin, M.Y.; Adomat, H.; Guns, E.S. Ginsenoside-mediated blockade of 1alpha,25-dihydroxyvitamin D3 inactivation in human liver and intestine in vitro. J. Steroid Biochem. Mol. Biol. 2014, 141, 94–103. [Google Scholar] [CrossRef]
- Deb, S.; Chin, M.Y.; Adomat, H.; Guns, E.S. Abiraterone inhibits 1alpha,25-dihydroxyvitamin D3 metabolism by CYP3A4 in human liver and intestine in vitro. J. Steroid Biochem. Mol. Biol. 2014, 144, 50–58. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Slominski, A.T.; Tuckey, R.C. Hydroxylation of 20-hydroxyvitamin D3 by human CYP3A4. J. Steroid Biochem. Mol. Biol. 2016, 159, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Stucci, L.S.; D’Oronzo, S.; Tucci, M.; Macerollo, A.; Ribero, S.; Spagnolo, F.; Marra, E.; Picasso, V.; Orgiano, L.; Marconcini, R.; et al. Vitamin D in melanoma: Controversies and potential role in combination with immune check-point inhibitors. Cancer Treat. Rev. 2018, 69, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Lin, Q.; Ke, R.; Shan, X.; Yu, J.; Ni, X.; Lin, X.; Wang, B. Causal association between serum 25-Hydroxyvitamin D levels and cutaneous melanoma: A two-sample Mendelian randomization study. Front. Oncol. 2023, 13, 1154107. [Google Scholar] [CrossRef] [PubMed]
- Shellenberger, R.A.; Gowda, S.; Kurn, H.; Albright, J.; Mayo, M.H. Vitamin D insufficiency and serum levels related to the incidence and stage of cutaneous melanoma: A systematic review and meta-analysis. Melanoma Res. 2023, 33, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, C.; Lucas, R.M.; Hurst, C.; Kimlin, M.G. Vitamin D deficiency at melanoma diagnosis is associated with higher Breslow thickness. PLoS ONE 2015, 10, e0126394. [Google Scholar] [CrossRef] [PubMed]
- Chandler, P.D.; Chen, W.Y.; Ajala, O.N.; Hazra, A.; Cook, N.; Bubes, V.; Lee, I.M.; Giovannucci, E.L.; Willett, W.; Buring, J.E.; et al. Effect of Vitamin D3 Supplements on Development of Advanced Cancer: A Secondary Analysis of the VITAL Randomized Clinical Trial. JAMA Netw. Open 2020, 3, e2025850. [Google Scholar] [CrossRef] [PubMed]
- Pilz, S.; Trummer, C.; Theiler-Schwetz, V.; Grubler, M.R.; Verheyen, N.D.; Odler, B.; Karras, S.N.; Zittermann, A.; Marz, W. Critical Appraisal of Large Vitamin D Randomized Controlled Trials. Nutrients 2022, 14, 303. [Google Scholar] [CrossRef] [PubMed]
- Ben-Eltriki, M.; Deb, S.; Adomat, H.; Tomlinson Guns, E.S. Calcitriol and 20(S)-protopanaxadiol synergistically inhibit growth and induce apoptosis in human prostate cancer cells. J. Steroid Biochem. Mol. Biol. 2016, 158, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Ben-Eltriki, M.; Deb, S.; Guns, E.S.T. 1alpha,25-Dihydroxyvitamin D(3) synergistically enhances anticancer effects of ginsenoside Rh2 in human prostate cancer cells. J. Steroid Biochem. Mol. Biol. 2021, 209, 105828. [Google Scholar] [CrossRef]
- Ben-Eltriki, M.; Deb, S.; Shankar, G.; Meckling, G.; Hassona, M.; Yamazaki, T.; Fazli, L.; Chin, M.Y.; Tomlinson Guns, E.S. Anti-Tumor Effects of Ginsenoside 20(S)-Protopanaxadiol and 1,25-Dihydroxyvitamin D3 Combination in Castration Resistant Prostate Cancer. Medicines 2021, 8, 28. [Google Scholar] [CrossRef]
- Sheng, L.; Anderson, P.H.; Turner, A.G.; Pishas, K.I.; Dhatrak, D.J.; Gill, P.G.; Morris, H.A.; Callen, D.F. Identification of vitamin D(3) target genes in human breast cancer tissue. J. Steroid Biochem. Mol. Biol. 2016, 164, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Brozyna, A.; Jozwicki, W.; Tuckey, R.C. Vitamin D as an adjuvant in melanoma therapy. Melanoma Manag. 2015, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.Y.; Fu, T.; Leblanc, E.; Manson, J.E.; Feldman, D.; Linos, E.; Vitolins, M.Z.; Zeitouni, N.C.; Larson, J.; Stefanick, M.L. Calcium plus vitamin D supplementation and the risk of nonmelanoma and melanoma skin cancer: Post hoc analyses of the women’s health initiative randomized controlled trial. J. Clin. Oncol. 2011, 29, 3078–3084. [Google Scholar] [CrossRef] [PubMed]


| CYP | Tissue Localization | Substrate | Role (Anabolism/Catabolism) | Tissue Expression | Polymorphic (Yes/No) | Refs. |
|---|---|---|---|---|---|---|
| CYP11A1 | Mitochondria | D3 | Catabolism | Brain, GI tract, Skin | Yes | [2,42,43] |
| CYP24A1 | Mitochondria | Calcitriol 25(OH)D3 1α,24,25(OH)3D3 1α,23,25(OH)3D3 | Catabolism Hydroxylation | Kidney, Intestine, Skin | Yes | [15,37,44] |
| CYP27A1 | Mitochondria | D3 1α(OH)D3 | Anabolism and Catabolism Pharmacological Hydroxylation | Liver, Skin, Macrophage | Yes | [37,44,45,46] |
| CYP27B1 | Mitochondria | 25(OH)D3 | Anabolism and Catabolism Hydroxylation | Kidney, Skin | Yes | [15,37,44] |
| CYP2R1 | Microsomes | D3 | Physiological Hydroxylation | Liver, Skin | Yes | [37,46,47] |
| CYP3A4 | Microsomes | Calcitriol D3 | Hydroxylation | Liver, Intestine, Skin | Yes | [37,46] |
| CYP2D25 | Microsomes | D3 | Hydroxylation | Liver, Kidney | No | [46] |
| CYP2J2/3 | Microsomes | D3 | Hydroxylation Catabolism | Liver, Heart, Placenta, Brain | Yes | [36,46] |
| Vitamin D | Model | Signaling Proteins | Significance | Refs. |
|---|---|---|---|---|
| 1α,25(OH)2D3 | -Human melanoma cell lines -A375 -ME18 -MeWo -RPMI 7951 -SK Mel 28 -SKMEL-188 -WM35 -WM1341 | -increase in Bcl-2 and Bcl-X (antiapoptotic) -decrease in BAX, GOS2, DAP-3, FADD, and caspases (proapoptotic) -upregulation of beclin-1 | prodifferentiation | [2,58] |
| 1,25,26(OH)3D3 | -malignant melanoma MM96 cells -mouse B16 hamster Bomirski melanomas | -increasing the expression of TGFβ -blockade of epithelial–mesenchymal transition (EMT) -blocking of MMP-2 and MMP-9 secretion | suppressed proliferation | [2] |
| 21(OH)D | -SKMEL-188 -hamster Ab -AbC1 melanoma lines | -decreased expression of VDR, RXR, PDIA3, and CYP2R1 | antiproliferative activity | [2,58] |
| 20(OH)D3 | -in vitro assays -melanoma cells -melanocytes -hamster melanoma | -inhibition of NFκβ activity -CYP24A1 | enhanced anti-melanoma activity | [2] |
| 20(OH)D2 | -in vitro -malignant cells -melanocytes -hamster melanoma | VDR-mediated | antiproliferative activity | [2] |
| 20,23(OH)2D | -in vitro assays -malignant cells -melanocytes -hamster melanoma | -VDR-mediated -Albumin | antiproliferative | [2,58] |
| 20,23(OH)2D3 | malignant cells | -VDR-mediated | antiproliferative | [2] |
| 20,24(OH)2D3 | -in vitro -immunostaining of melanomas | -CYP24A1 | anti-melanoma activity | [2] |
| 20,25(OH)2D3 | -human melanoma cells | -CYP24A1 | inhibits proliferation | [2,55] |
| 1,25(OH)2D3 25(OH)D3 | -human melanoma cell lines -MeWo -MeWo (EB1089) -SkMeI28 -SkMeI28 -(EB1089)2.05 -SkMeI25 -IGR -MeIJuso | -VDR-mediated -CYP24A1 | antiproliferative | [60] |
| 1,25(OH)2D3 | -human melanoma cell lines -MeWo -SkMeI5 | VDR-mediated -CYP24A1 | inhibits proliferation | [59] |
| 1,25(OH)2D3 |
-RPMI 7951 (high VDR) -SK-MEL-28 (low VDR) | [64] | ||
| 1,25(OH)2D3 | -B16-F10 | -increase in cleaved caspase-3, caspase 8, caspase 9, Beclin, and PARP | antiproliferative | [65] |
| 1,25(OH)2D3 | -human melanoma cell lines -MeWo -WM1341 | -VDR-mediated | induce apoptosis | [66] |
| 1,25(OH)2D3 | -human melanoma cell lines-G-361/A3 | -VDR-mediated -nuclear factor KB (NF-KB) | anti-inflammatory action | [67] |
| 1,25(OH)2D3 | -human melanoma cell line | -blocks the formation of the sphingolipid degradation product sphingosine 1-phosphate (S1P) | induce apoptosis | [68] |
| 1,25(OH)2D3 | -CRL-1619 | -inhibition of oxidative DNA/RNA damage | anti-inflammatory action | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Eltriki, M.; Gayle, E.J.; Paras, J.M.; Nyame-Addo, L.; Chhabra, M.; Deb, S. Vitamin D in Melanoma: Potential Role of Cytochrome P450 Enzymes. Life 2024, 14, 510. https://doi.org/10.3390/life14040510
Ben-Eltriki M, Gayle EJ, Paras JM, Nyame-Addo L, Chhabra M, Deb S. Vitamin D in Melanoma: Potential Role of Cytochrome P450 Enzymes. Life. 2024; 14(4):510. https://doi.org/10.3390/life14040510
Chicago/Turabian StyleBen-Eltriki, Mohamed, Erysa J. Gayle, Jhoanne M. Paras, Louisa Nyame-Addo, Manik Chhabra, and Subrata Deb. 2024. "Vitamin D in Melanoma: Potential Role of Cytochrome P450 Enzymes" Life 14, no. 4: 510. https://doi.org/10.3390/life14040510