
Aroplectrus dimerus (Hymenoptera: Eulophidae), Ectoparasitoid of the Nettle Caterpillar, Oxyplax pallivitta (Lepidoptera: Limacodidae): Evaluation in the Hawaiian Islands

Abstract
:1. Introduction
2. Materials and Methods
2.1. Explorations and Origin of the Parasitoid Colony
2.2. Identity of the Primary and Secondary Parasitoids
2.3. Host Rearing
2.4. Parasitoid Rearing
2.5. Reproductive Parameters and Immature Measurements
2.6. Host Specificity Assessment
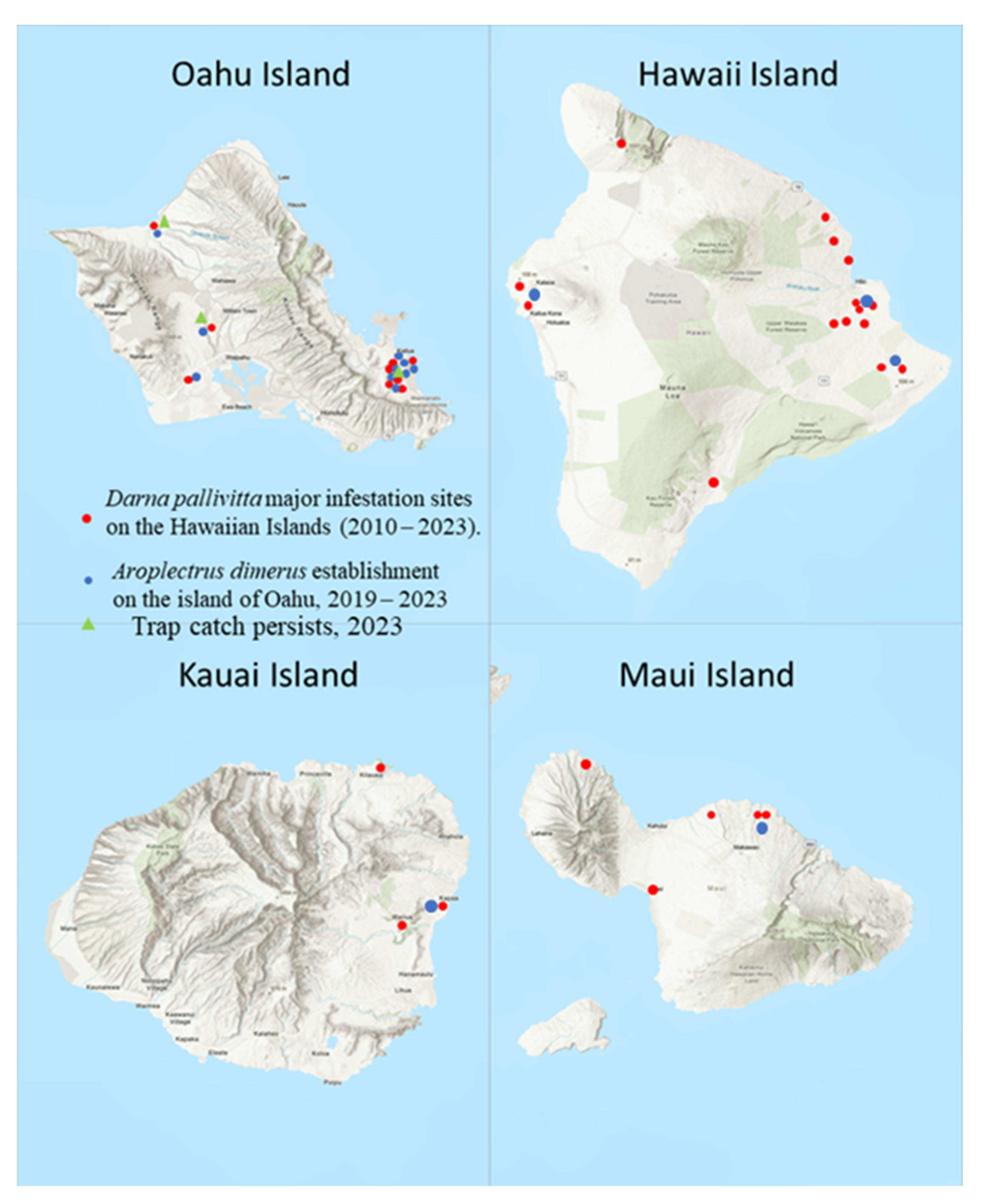
2.7. Colonization and Establishment Records on the Islands
2.8. Infestation Rates on Major Infested Sites on Oahu Island
2.9. Rates of Parasitism on Oahu Island
2.10. Trapping
2.11. Statistical Analysis and Vouchers
3. Results
3.1. Exploration
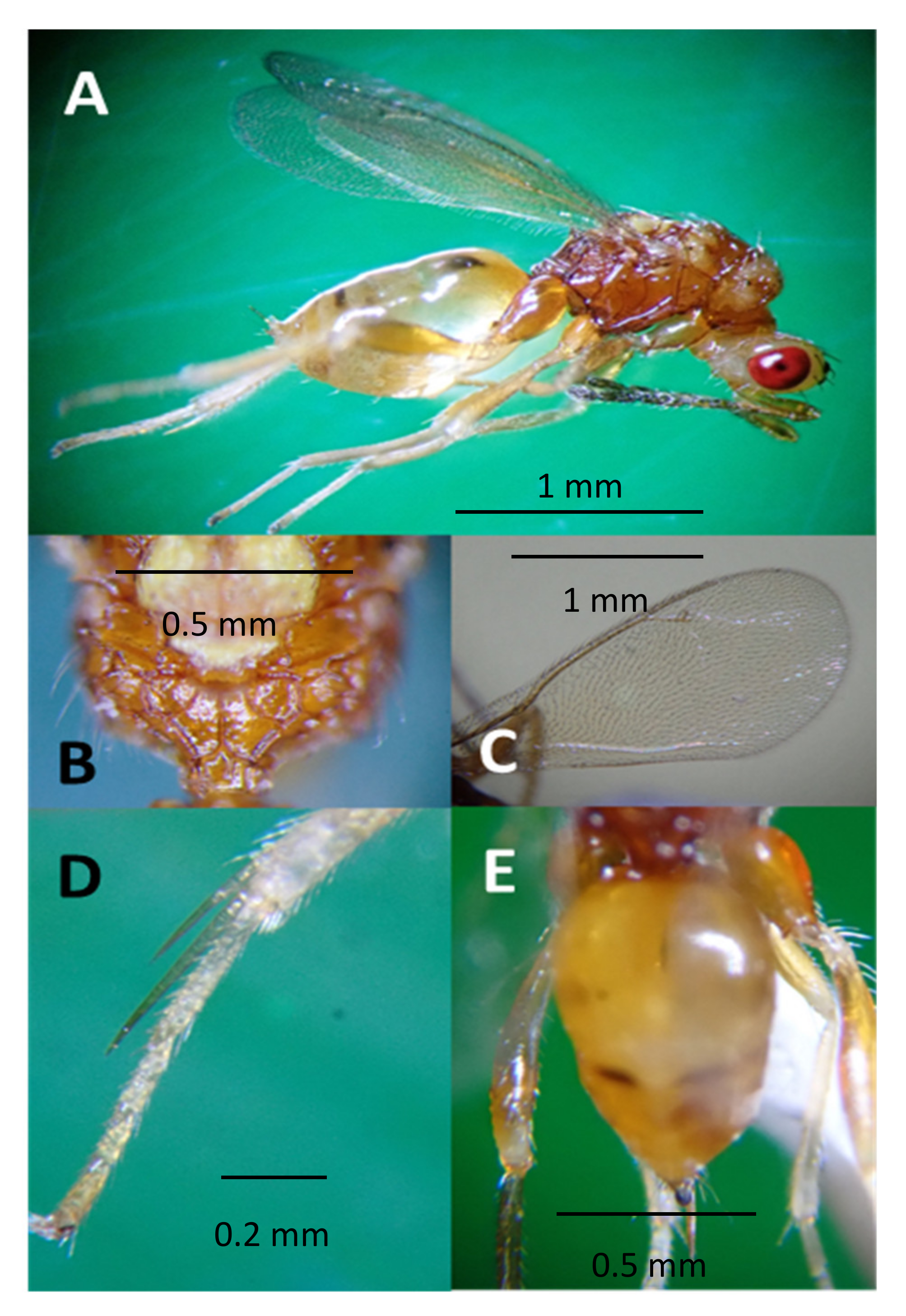
3.2. Identity of Parasitoids
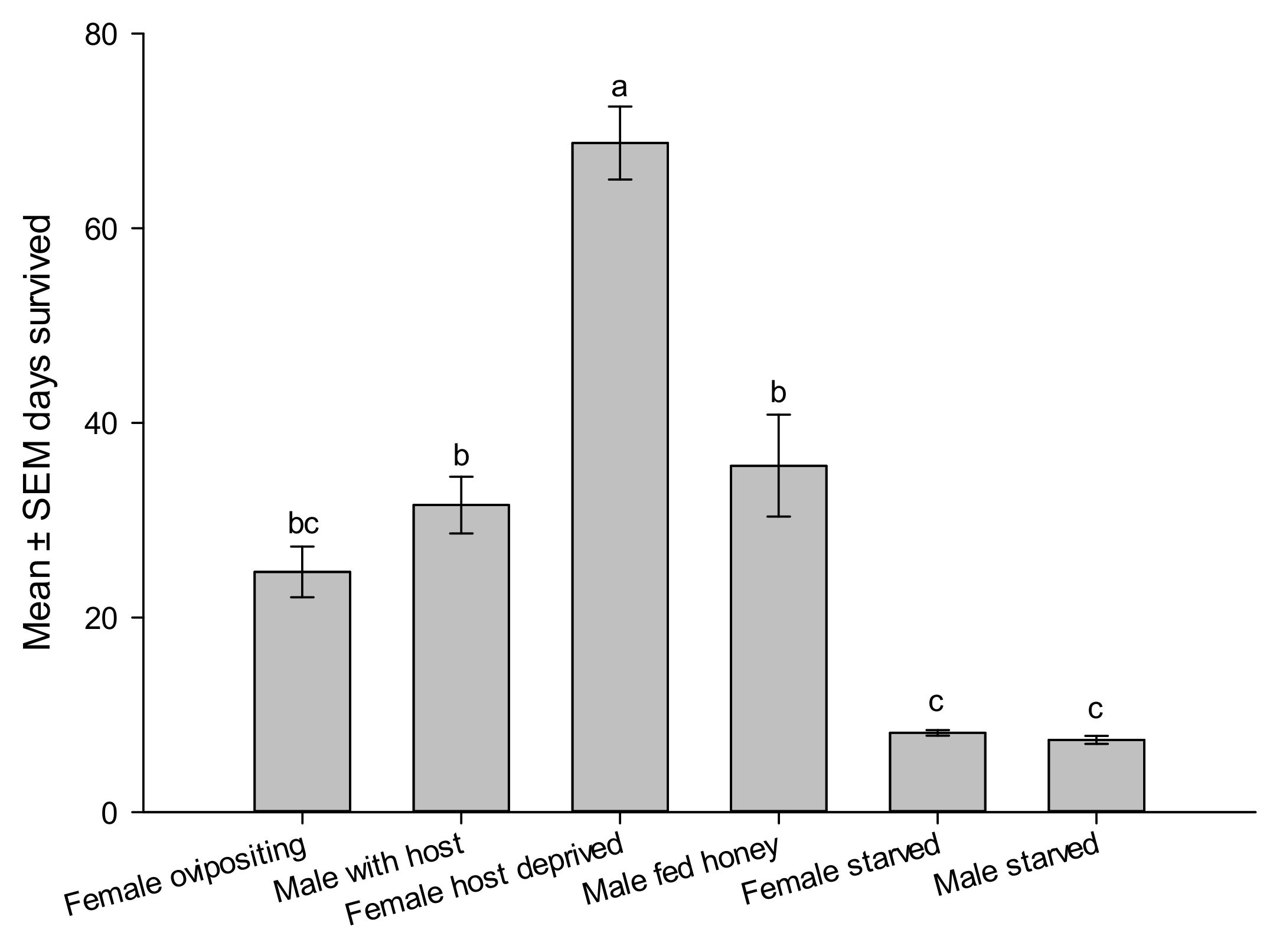
3.3. Life History and Reproductive Performance
3.4. Host Specificity Tests
3.5. Colonization Records on the Islands
3.6. Field Parasitism and Establishment
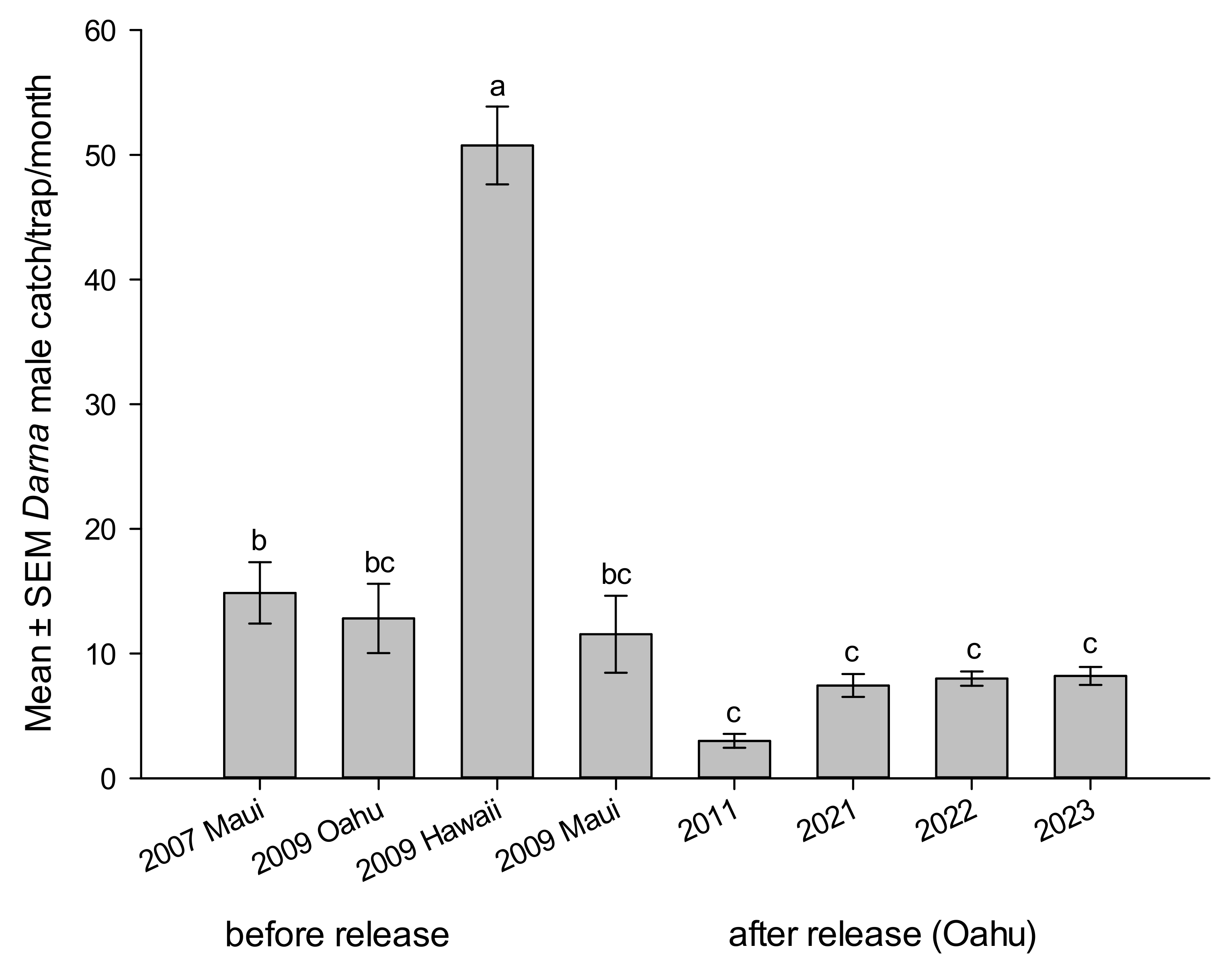
3.7. Male Trap Catches and Pupal Infestation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conant, P.; Hara, A.H.; Nakahara, L.M.; Heu, R.A. Nettle caterpillar Darna pallivitta Moore (Lepidoptera: Limacodidae). In New Pest Advisory; State of Hawaii Department of Agriculture: Honolulu, HI, USA, 2002. [Google Scholar]
- Kishimoto, C.M. The Stinging Nettle Caterpillar, Darna pallivitta (Moore) (Lepidoptera: Limacodidae) in Hawaii: Its Current Host Range, Biology, and Population Dynamics. Master’s Thesis, University of Hawaii at Manoa, Honolulu, HI, USA, 2006; p. 91. [Google Scholar]
- Hara, A.H.; Kishimoto, C.M.; Niino-DuPonte, R.Y. Host range of the nettle caterpillar Darna pallivitta (Moore) (Lepidoptera: Limacodidae) in Hawaii. In Potential Invasive Pests of Agricultural Crops; Peňa, J.E., Ed.; CAB International: Wallingford, UK, 2013; pp. 183–191. [Google Scholar]
- Chun, S.; Hara, A.; Niino-DuPonte, R.; Nagamine, W.; Conant, P.; Hirayama, C. Stinging Nettle Caterpillar, Darna pallivitta, Pest Alert; University of Hawaii: Honolulu, HI, USA, 2011; p. 2. Available online: https://scholarspace.manoa.hawaii.edu/server/api/core/bitstreams/9fa2bd14-6088-4d29-a9cc-e40a0b56a6e5/content (accessed on 23 January 2024).
- Jang, E.B.; Siderhurst, M.S.; Conant, P.; Siderhurst, L.A. Phenology and population radiation of the nettle caterpillar, Darna pallivitta (Moore) (Lepidoptera: Limacodidae) in Hawaii. Chemoecology 2009, 19, 7–12. [Google Scholar] [CrossRef]
- Anonymous. Nettle Caterpillar, Darna Pallivitta (Lepidoptera: Limacodidae). Maui Nature, Invasive Species. Nettle Caterpillar. Available online: https://www.mauiinformationguide.com/nettle-caterpillar.php (accessed on 8 January 2024).
- Nagamine, W.T.; Epstein, M.E. Chronicles of Darna pallivitta (Moore 1877) (Lepidoptera: Limacodidae): Biology and larval morphology of a new pest in Hawaii. Pan-Pac. Entomol. 2007, 83, 120–135. [Google Scholar] [CrossRef]
- Ramadan, M.M.; Kaufman, L.V.; Wright, M.G. Insect and weed biological control in Hawaii: Recent case studies and trends. Biol. Control 2023, 179, 105170. [Google Scholar] [CrossRef]
- Areces-Berazain, F. Darna pallivitta (Nettle Caterpillar); CAB International: Wallingford, UK, 2022. [Google Scholar] [CrossRef]
- Cock, M.J.W.; Godfray, H.C.J.; Holloway, J.D. (Eds.) Slug and Nettle Caterpillars: The Biology, Taxonomy and Control of the Limacodidae of Economic Importance on Palms in South-East Asia; CAB International: Wallingford, UK, 1987; 270p. [Google Scholar]
- Holloway, J.D. The moths of Borneo. Part I. Key to families; Families Cossidae, Metarbelidae, Ratardidae, Dudgeoneidae, Epipyropidae and Limacodidae. Malay. Nat. J. 1986, 40, 1–166. [Google Scholar]
- Holloway, J.D. The Moths of Borneo. 2006. Available online: http://www.mothsofborneo.com/part-1/limacodidae/limacodidae-42-7.php (accessed on 8 January 2024).
- Shao, K.; Chung, K. Oxyplax pallivitta (Moore, 1877). The National Checklist of Taiwan (Catalogue of Life in Taiwan, TaiCoL); Version 1.11; Taiwan Biodiversity Information Facility (TaiBIF): Taipei, Taiwan, 2022; Available online: https://www.gbif.org/species/11014495 (accessed on 5 April 2024).
- Lin, K.S. Revision of the tribe Euplectrini from Taiwan. Part 1. (Hymenoptera: Eulophidae.). Q. J. Taiwan Museum. 1963, 161, 101–124. [Google Scholar] [CrossRef]
- Khan, M.Y.; Shafee, S.A. Species of the genus Pediobius Eulophidae Entedontinae from India. J. Bombay Nat. Hist. Society. 1982, 79, 370–374. Available online: https://www.biodiversitylibrary.org/part/156894 (accessed on 23 January 2024).
- Nishida, G. Hawaiian Terrestrial Arthropod Checklist, 4th ed.; Nishida, G.M., Ed.; Bishop Museum: Honolulu, HI, USA, 2002; 262p, Available online: http://www2.bishopmuseum.org/HBS/checklist/query.asp?grp=Arthropod (accessed on 20 October 2023).
- Siderhurst, M.S.; Jang, E.B.; Hara, A.H.; Conant, P. n-Butyl (E)-7,9-decadienoate: Sex pheromone component of the nettle caterpillar, Darna pallivitta. Entomol. Exp. Et Applicata 2007, 125, 63–69. [Google Scholar] [CrossRef]
- JMP®, Version 11, SAS Institute Inc.:: Cary, NC, USA, 2023.
- Narendran, T.C.; Fousi, K.; Rajmohana, K.; Mohan, C. A new species of Eulophidae (Hymenoptera: Chalcidoidea) parasitoid on the slug caterpillar pest, Contheyla rotunda Hampson (Lepidoptera: Cochliidae) of coconut in Kerala. J. Appl. Zool. Res. 2002, 13, 31–34. [Google Scholar]
- Noyes, J. UCD: Universal Chalcidoidea Database (version Sep 2007). In Species 2000 & ITIS Catalogue of Life; Roskov, Y., Ower, G., Orrell, T., Nicolson, D., Bailly, N., Kirk, P.M., Bourgoin, T., DeWalt, R.E., Decock, W., van Nieukerken, E., et al., Eds.; Annual Checklist; Species 2000; Naturalis: Leiden, The Netherlands, 2019; Available online: https://www.catalogueoflife.org/annual-checklist/2019 (accessed on 20 January 2024).
- Narendran, T.C. Fauna of India, Eulophinae (Hymenoptera: Eulophidae). Zool. Surv. India Eulophidae 2011, 1–342. [Google Scholar]
- Hansson, C. Eulophidae of Costa Rica (Hymenoptera: Chalcidoidea), 1. Mem. Am. Entomol. Inst. 2002, 67, 1–290. [Google Scholar]
- Hansson, C.; Smith, M.A.; Janzen, D.H.; Hallwachs, W. Integrative taxonomy of New World Euplectrus Westwood (Hymenoptera, Eulophidae), with focus on 55 new species from Area de Conservación Guanacaste, northwestern Costa Rica. ZooKeys 2015, 485, 1–236. [Google Scholar] [CrossRef] [PubMed]
- Walker, F. Characters of some undescribed species of Chalcidites. (Continued). Ann. Mag. Nat. Hist. 1846, 17, 177–185. [Google Scholar] [CrossRef]
- Kerrich, G.J. A revision of the tropical and subtropical species of the eulophid genus Pediobius Walker (Hymenoptera: Chalcidoidea). Bull. Br. Mus. (Nat. Hist.) Entomol. 1973, 29, 115–200. [Google Scholar]
- Schauff, M.E. The Holarctic genera of Entedoninae (Hymenoptera: Eulophidae). Contrib. Am. Entomol. Institute. 1991, 26, 1–109. [Google Scholar]
- Ghosh, S.M.; Abdurahiman, U.C. On the biology of Pediobius imbreus (Hymenoptera: Eulophidae), a hyperparasite of Opisina arenosella, the black-headed caterpillar of coconut. COCOS 1988, 6, 27–30. [Google Scholar] [CrossRef]
- Ghosh, S.M.; Chandrasekharan, K.; Abdurahiman, U.C. The bionomics of Pediobius imbreus (Hymenoptera) and its impact on the biological control of the coconut caterpillar. J. Ecobiol. 1993, 5, 161–166. [Google Scholar]
- Siderhurst, M.S.; Jang, E.B.; Carvalho, L.A.F.N.; Nagata, J.T.; Derstine, N.T. Disruption of Darna pallivitta (Lepidoptera: Limacodidae) by conventional and mobile pheromone deployment. J. Insect. Sci. 2015, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Conant, P.; Hirayama, C.K.; Kishimoto, C.M.; Hara, A.H. Trichogramma papilionis (Nagarkatti), the first recorded Trichogramma species to parasitize eggs in the family Limacodidae. Proc. Hawaii. Entomol. Soc. 2006, 38, 133–135. [Google Scholar]
- Singh, B.P.; Singh, R.P.; Verma, V.D. New record of Aroplectrus dimerus Linn. [sic] and Platyplectrus sp. as a larval parasite of slug caterpillar (Parasa bicolor Walk) [sic] from U.P. Farm Sci. J. 1988, 3, 199–200. Available online: http://trophort.com/002/169/002169592.html (accessed on 18 January 2024).
- Philippine Coconut Authority. The Small Limacodid Pethocrates sp. a Pest on Coconut. CPD Technoguide 1999, 5. Available online: http://www.pca.da.gov.ph/pdf/techno/limacodid.pdf (accessed on 18 January 2024).
- Zhu, C.D.; Huang, D.W. A taxonomic study on Eulophidae from Guangxi, China (Hymenoptera: Chalcidoidea). Acta Zootaxonomica Sin. 2002, 27, 583–607. [Google Scholar]
- Fry, J.M. Natural Enemy Databank, 1987. A Catalogue of Natural Enemies of Arthropods Derived from Records in the CIBC Natural Enemy Databank. CAB International: Wallingford, UK, 1989; 185p. [Google Scholar]
- Yoshimoto, H. Darna pallivitta (Moore) from Okinawa I., a limacodid moth new to Japan. Jpn. Heterocerists’ J. 1997, 192, 273–274. Available online: http://publ.moth.jp/tsushin/151-200/jhj192.pdf (accessed on 23 January 2024).
- Wijesekara, G.A.W.; Gunawardena, T. A Synopsis of the Tribe Euplectrini of Hymenoptera Eulophidac of Sri Lanka. Trop. Agric. Res. 1989, 1, 114–1120. [Google Scholar]
- Bouček, Z. Australian Chalcidoidea (Hymenoptera). A Biosystematic Revision of Genera of Fourteen Families, with Reunification of Species; Commonwealth Institution of Entomology, CAB International: Wallingford, UK, 1988; 832p. [Google Scholar]
- Peck, O. The taxonomy of the Nearctic species of Pediobius (Hymenoptera: Eulophidae), especially Canadian and Alaskan forms. Can. Entomol. 1985, 117, 647–704. [Google Scholar] [CrossRef]
- Gates, M.W.; Lill, J.T.; Kula, R.R.; O’Hara, J.E.; Wahl, D.B.; Smith, D.R.; Whitfield, J.B.; Murphy, S.M.; Stoepler, T.M. Review of Parasitoid Wasps and Flies (Hymenoptera, Diptera) Associated with Limacodidae (Lepidoptera) in North America, with a Key to Genera. Proc. Entomol. Soc. Wash. 2012, 114, 24–110. [Google Scholar] [CrossRef]
- Budhachandra, T.; Khan, M.A.; Pandey, S.; Manhanvi, K. A new species of genus Pediobius Walker (Hymenoptera: Eulophidae) with new records of eulophid parasitoids from Uttarakhand, India. Pantnagar J. Res. 2013, 11, 29. [Google Scholar]
- Chalapathi Rao, N.B.V.; Padma, E.; Ramanandam, G. Pediobius imbreus (Hymenoptera: Eulophidae), a promising parasitoid of coconut slug caterpillar, Macroplectra nararia Moore (Lepidoptera: Limacodidae) in Andhra Pradesh. J. Pest Manag. Hortic. Ecosyst. 2017, 23, 176–178. [Google Scholar]
- Yoshimoto, C.M. Title: Synopsis of Hawaiian Eulophidae including Aphelininae (Hym.: Chalcidoidea). Pac. Insects 1965, 7, 665–699. [Google Scholar]
- HDOA. Natural Enemy of Stinging Caterpillar to be Released on O’ahu. News Release 2010, 10–17. Available online: http://hawaii.gov/hdoa/news/2010-news-releases/natural-enemyof-stinging-caterpillar-to-be-released-on-o-ahu (accessed on 15 May 2011).
- HDOA. Kaua’i Residents Asked to Report Sightings of Stinging Caterpillar. News Releases 2011, 11–11. Available online: http://hawaii.gov/hdoa/news/news-releases-2011/kaua-i-residentsasked-to-report-sightings-of-stinging-caterpillar (accessed on 8 June 2011).
- California Department of Food and Agriculture (CDFA). Darna pallivitta, p. 21; California Plant Pest and Disease Report; California Department of Food and Agriculture: Sacramento, CA, USA, 2005; Volume 22, pp. 1–76.
- California Department of Food and Agriculture (CDFA). Darna pallivitta, p. 12; California Plant Pest and Disease Report; California Department of Food and Agriculture: Sacramento, CA, USA, 2006; Volume 23, pp. 1–117.
- Bautista, R.C.; Yalemar, J.A.; Conant, P.; Arakaki, D.K.; Reimer, N. Taming a stinging caterpillar in Hawaii with a parasitic wasp. Biocontrol News Inf. 2014, 35, 9N–17N. Available online: https://www.cabi.org/BNI (accessed on 10 January 2024).
- Matsunaga, J.N.; Howarth, F.G.; Kumashiro, B.R. New State Records and Additions to the Alien Terrestrial Arthropod Fauna in the Hawaiian Islands. Proc. Hawaii. Entomol. Soc. 2019, 51, 1–71. [Google Scholar]
- Koop, A.L. New Pest Advisory Group Report. Darna pallivitta Moore: Nettle Caterpillar. 2006. Available online: https://download.ceris.purdue.edu/file/3058 (accessed on 8 February 2024).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family of Lepidoptera | Scientific and Common Name | Status and Source | Host Plant Common and Scientific Name |
|---|---|---|---|
| Erebidae | Podomachla apicalis (Walker) a leaf-feeder | Beneficial, Lab-reared | fireweed leaves, Senecio madagascariensis |
| Erebidae | Secusio extensa (Butler) a leaf-feeder | Beneficial Lab-reared | fireweed leaves, Senecio madagascariensis |
| Choreutidae | Choreutis sp. a leaf-tier | Pest Field-collected | weeping fig leaves, Ficus benjamina |
| Crambidae | Diaphania nitidalis Cramer pickleworm | Pest Lab-reared | cucumber flowers/fruit, Cucumis sativa, Pipturis albidus |
| Crambidae | Omiodes blackburni (Butler) coconut leaf roller | Endemic Lab-reared | coconut leaves, Cocos nucifera |
| Crambidae | Udea stellata (Butler) a leaf-feeder | Endemic Lab-reared, | mamaki leaves, |
| Ethmiidae | Ethmia nigroapicella (Sallmuller) kou leafworm | Pest Field-collected | kou leaves, Cordia subcordata |
| Geometridae | Anacamptodes fragilaria (Grossbeck), koa haole looper | Pest Field-collected | koa-haole leaves, Leucaena leucocephala |
| Geometridae | Macaria abydata Guenee koa haole moth | Pest Field-collected, | koa-haole leaves, Leucaena leucocephala |
| Lycaenidae | Lampides boeticus (Linnaeus) bean butterfly | Pest Field-collected | rattlepod beans, Crotalaria sp. |
| Noctuidae | Achaea janata (Linnaeus) croton caterpillar | Pest Field-collected | castor bean leaves, Ricinus communis |
| Noctuidae | Agrotis sp. a cutworm | Pest Lab-reared | cotton leaves, Gossypium hirsutum |
| Noctuidae | Anomis flava (Fabricius) hibiscus caterpillar | Pest Lab-reared | cotton leaves, Gossypium hirsutum |
| Noctuidae | Heliothis virescens (Fabricius) tobacco budworm | Pest Field-collected | love-in-a-mist flowers, Passiflora foetida |
| Noctuidae | Pandesma anysa Guenee a leaf-feeder | Pest Field-collected | opiuma leaves, Pithecellobium dulce |
| Noctuidae | Spodoptera mauritia (Boisduval), lawn armyworm | Pest Lab-reared | undetermined grass species |
| Nymphalidae | Agraulis vanillae (Linnaeus) passion vine butterfly | Pest Field-collected | passion vine leaves, Passiflora edulis |
| Nymphalidae | Vanessa cardui (Linnaeus) painted lady | Pest Field-collected | cheeseweed leaves, Malva parviflora |
| Pieridae | Pieris rapae (Linnaeus) imported cabbageworm | Pest Field-collected | broccoli leaves, Brassica oleracea |
| Plutellidae | Plutella xylostella (Linnaeus), diamondback moth | Pest Field-collected | broccoli leaves, Brassica oleracea |
| Pyralidae | Hellula undalis (Fabricius) imported cabbage webworm | Pest Field-collected | mustard cabbage leaves, Brassica juncea |
| Sphingidae | Daphnis nerii (Linnaeus) oleander hawk moth | Pest Field-collected | oleander leaves, Nerium oleander |
| Tortricidae | Croesia zimmermani Clarke a biocontrol agent | Beneficial Field-collected | blackberry leaves, Rubus argutus |
| Tortricidae | Cryptophlebia ombrodelta (Lower), litchi fruit moth | Pest Field-collected | undetermined legume species |
| Tortricidae | Episimus utilis Zimmerman a biocontrol agent | Beneficial Field-collected | x-mas berry leaves, Schinus terebinthifolius |
| Reproductive Parameter | n | Mean ± SEM | Range | Unit |
|---|---|---|---|---|
| Preoviposition period | 40 | 1.60 ± 0.15 | 0–5 | days |
| Oviposition period | 40 | 7.77 ± 0.65 | 2–21 | days |
| Post-oviposition period | 36 | 10.86 ± 2.42 | 0–60 | days |
| Age at peak oviposition | 40 | 7.50 ± 0.79 | 2–24 | days |
| Highest oviposition eggs/day | 40 | 9.45 ± 0.55 | 4–19 | number eggs |
| Egg deposition /day/larva | 280 | 4.39 ± 0.26 | 0–19 | number eggs |
| Fecundity total eggs deposited/female | 40 | 41.67 ± 4.09 | 10–130 | total eggs |
| Adult offspring emergence rate | 15 | 58.20 ± 3.00 | 25–79 | percentage |
| Female longevity | 40 | 24.67 ± 2.61 | 8–76 | days |
| Male longevity * | 40 | 31.55 ± 2.93 | 8–90 | days |
| Sex ratio (% females’ offspring) ** | 24 | 63.23 ± 3.00 | 8.9–85.7 | % female |
| Sex ratio (% males’ offspring) | 24 | 36.76 ± 3.03 | 14.2–91.04 | % male |
| Life span | 54 | 10.57 ± 0.19 | 9–13 | days |
| Egg incubation period | 14 | 2.0 ± 0.0 | 2 | days |
| Larval developmental period | 14 | 4.5 ± 0.14 | 4–5 | days |
| Pupal developmental period | 14 | 5.64 ± 0.20 | 5–7 | days |
| Measurements of immatures | ||||
| Egg length | 15 | 0.348 ± 0.005 | 0.32–0.40 | mm |
| Egg width | 15 | 0.139 ± 0.003 | 0.12–0.16 | mm |
| Mature larval length | 15 | 2.28 ± 0.137 | 1.25–3.0 | mm |
| Mature larval width | 15 | 0.88 ± 0.054 | 0.57–1.25 | mm |
| Pupal length | 15 | 2.78 ± 0.063 | 2.44–3.12 | mm |
| Pupal width | 15 | 0.95 ± 0.035 | 0.76–1.16 | mm |
| Species Name | Parasitoid Eggs Deposited on Larvae (Mean ± SEM) | No. Larvae Parasitized | No. Parasitoids Emerging | No. and % Moths of Test Species Emerging |
|---|---|---|---|---|
| Podomachla apicalis | 0 b | 0 | 0 | 20 (100%) |
| O. pallivitta (control) | 3.0 ± 0.9 a | 10 (50%) | 49 (42 ♀, 7 ♂) | - |
| Secusio extensa | 0 b | 0 | 0 | 15 (75%) |
| O. pallivitta (control) | 5.1 ± 1.1 a | 13 (65%) | 94 (61 ♀, 33 ♂) | - |
| Choreutis sp. | 0 b | 0 | 0 | 15 (75%) |
| O. pallivitta (control) | 4.1 ± 0.9 a | 12 (60%) | 73 (52 ♀, 21 ♂) | - |
| Diaphania nitidalis | 0 b | 0 | 0 | 20 (100%) |
| O. pallivitta (control) | 3.4 ± 0.7 a | 14 (70%) | 67 (52 ♀, 23 ♂) | - |
| Omiodes blackburni | 0 b | 0 | 0 | 14 (70%) |
| O. pallivitta (control) | 4.5 ± 1.2 a | 11 (55%) | 87 (62 ♀, 25 ♂) | - |
| Udea stellata | 0 b | 0 | 0 | 20 (100%) |
| O. pallivitta (control) | 5.1 ± 1.1 a | 12 (60%) | 38 (26 ♀, 12 ♂) | - |
| Ethmia nigroapicella | 0 b | 0 | 0 | 15 (75%) |
| O. pallivitta (control) | 3.5 ± 0.8 a | 13 (65%) | 16 (10 ♀, 6 ♂) | - |
| Anacamptodes fragilaria | 0 b | 0 | 0 | 0 (0%) 1 |
| O. pallivitta (control) | 4.9 ± 0.9 a | 17 (85%) | 36 (18 ♀, 18 ♂) | - |
| Macaria abydata | 0 b | 0 | 0 | 4 (20%) |
| O. pallivitta (control) | 3.6 ± 0.9 a | 12 (60%) | 27 (20 ♀, 7 ♂) | - |
| Lampides boeticus | 0 b | 0 | 0 | 20 (100%) |
| O. pallivitta (control) | 5.5 ± 1.0 a | 16 (80%) | 45 (28 ♀, 17 ♂) | - |
| Achaea janata | 0 b | 0 | 0 | 19 (95%) |
| O. pallivitta (control) | 4.7 ± 1.0 a | 14 (70%) | 88 (54 ♀, 34 ♂) | - |
| Agrotis sp. | 0 b | 0 | 0 | 1 (5%) 1 |
| O. pallivitta (control) | 3.7 ± 1.2 a | 11 (55%) | 49 (22 ♀, 29 ♂) | - |
| Anomis flava | 0 b | 0 | 0 | 20 (100%) |
| O. pallivitta (control) | 4.8 ± 0.9 a | 13 (65%) | 91 (61 ♀, 30 ♂) | - |
| Heliothis virescens | 0 b | 0 | 0 | 13 (65%) |
| O. pallivitta (control) | 5.1 ± 1.4 a | 9 (45%) | 60 (38 ♀, 22 ♂) | - |
| Pandesma anysa | 0 b | 0 | 0 | 18 (90%) |
| O. pallivitta (control) | 5.3 ± 1.1 a | 14 (70%) | 101 (70 ♀, 31 ♂) | - |
| Spodoptera mauritia | 0 b | 0 | 0 | 19 (95%) |
| O. pallivitta (control) | 4.2 ± 0.9 a | 14 (70%) | 80 (67 ♀, 13 ♂) | - |
| Agraulis vanillae | 0 b | 0 | 0 | 1 (5%) 1 |
| O. pallivitta (control) | 4.5 ± 0.9 | 14 (70%) | 76 (46 ♀, 30 ♂) | - |
| Vanessa cardui | 0 b | 0 | 0 | 19 (95%) |
| O. pallivitta (control) | 4.4 ± 1.0 a | 12 (60%) | 68 (45 ♀, 23 ♂) | - |
| Pieris rapae | 0 b | 0 | 0 | 19 (95%) |
| O. pallivitta (control) | 4.4 ± 1.0 a | 13 (65%) | 68 (39 ♀, 29 ♂) | - |
| Plutella xylostella | 0 b | 0 | 0 | 20 (100%) |
| O. pallivitta (control) | 3.5 ± 1.3 a | 8 (40%) | 44 (28 ♀, 16 ♂) | - |
| Hellula undalis | 0 b | 0 | 0 | 10 (50%) |
| O. pallivitta (control) | 4.8 ± 1.0 a | 12 (60%) | 56 (33 ♀, 23 ♂) | - |
| Daphnis nerii | 0 b | 0 | 0 | 18 (90%) |
| O. pallivitta (control) | 4.6 ± 0.9 a | 15 (75%) | 67 (6 ♀, 61 ♂) | - |
| Croesia zimmermani | 0 b | 0 | 0 | 19 (95%) |
| O. pallivitta (control) | 2.6 ± 0.8 a | 8 (40%) | 13 (9 ♀, 4 ♂) | - |
| Cryptophlebia ombrodelta | 0 b | 0 | 0 | 20 (100%) |
| O. pallivitta (control) | 3.0 ± 0.9 a | 11 (55%) | 33 (24 ♀, 9 ♂) | - |
| Episimus utilis | 0 b | 0 | 0 | 20 (100%) |
| O. pallivitta (control) | 3.9 ± 0.5 a | 17 (85%) | 42 (23 ♀, 19 ♂) | - |
| Island | Locality | Release Period | Major Infestation Release Sites Geographic Coordinates and Elevation | Release Sets and Range Wasps/Lot | Total Wasp Released |
|---|---|---|---|---|---|
| Oahu Island | Waimanalo, Winward Oahu 2010 | 17 May 2010– 18 November 2010 | Ahiki, (21°20′08.42″ N, 157°42′58.88″ W, 27 m AMSL) | 12 (50–441) | 1494 |
| “ | Waimanalo 2010 | 9 October 2010– 3 December 2010 | Leilani Nursery, (21°20′32.52″ N, 157°43′26.16″ W, 21 m AMSL) C and L Nursery, (21°19′38.78″ N, 157°42′57.10″ W, 89 m AMSL) | 5 (50–110) | 310 |
| “ | Waimanalo 2011 | 13 January 2011– 8 June 2011 | Ahiki, (21°20′08.42″ N, 157°42′58.88″ W, 27 m AMSL) | 5 (50–100) | 400 |
| “ | Central Oahu 2010 | 8 June 2010– 1 November 2010 | Kipapa Gulch, 21°27′32.80″ N, 158°00′57.32″ W, 215 m AMSL) Uka Elem. Sch, Mililani (21°26′14.33″ N, 158°00′55.49″ W, 179 m AMSL) Poloahilani St, Mililani (21°26′51.10″ N, 158°00′12.34″ W, 212 m AMSL) | 12 (20–386) | 849 |
| “ | Central Oahu 2011 | 2 February 2011– 14 June 2011 | Noholoa Park, Mililani (21°26′29.16″ N, 158V 00′29.55″ W, 184 m AMSL) Takenaka’s Nursery, Wahiawa (21°25′44.31″ N, 158°00′55.56″ W, 151 m AMSL) | 5 (50–200) | 610 |
| “ | Windward Oahu 2021 | 3 October 2021– 16 December 2021 | Olomana (21°21′55.00″ N, 157°44′22.61″ W, 56 m AMSL) Lanikai (21°23′30.78″ N, 157°42′56.97″ W, 5 m AMSL) Enchanted lake, (21°23′05.17″ N, 157°44′04.84″ W, 5 m AMSL) | 9 (40–70) | 526 |
| “ | Winward and North Oahu 2022 | 15 February 2022– 20 December 2022 | Haleiwa (21°35′33.15″ N, 158°06′12.88″ W, 1m AMSL) Pauahilani st. kaillua (21°23′29.96″ N, 157°43′27.53″ W, 21 m AMSL) | 22 (50–100) | 1465 |
| “ | Winward and Central Oahu 2023 | 6 February 2023– 23 August 2023 | Maunawili (21°22′50.57″ N, 157°45′22.49″ W, 37 m AMSL) Kaululena St. Mililani (21°27′17.52″ N, 158°00′13.31″ W, 222 m AMSL) | 8 (40–220) | 880 |
| Hawaii Island | North Kona 2010 | 5 August 2010– 30 October 2010 | 3-Ring ranch, Kailua, Kona (19°38′37.21′N, 155°57′55.67″ W, 252 m AMSL) Loloa Way (19°43′29.75″ N, 155°59′27.00″ W, 325 m AMSL) Hawaiian Sunshine (21°20′26.44″ N, 157°43′0324″ W, 13 m AMSL) | 7(40–100) | 420 |
| “ | North Hilo and Puna 2010 | 16 June 2010– 12 November 2010 | Umauma, (19°54′18.50″ N, 155°08′28.8″ W, 115 m AMSL) Kurtistown, (19°35′34.87″ N, 155°03′27.95″ W, 200 m AMSL) Stainback (UH Exptl. Sta.) (19°39′11.48″ N, 155°02′58.20′W, 77 m AMSL) | 7 (50–250) | 800 |
| “ | Kona, North Kohala, North, South Hilo 2011 | 16 January 2011– 21 March 2011 | Puna Orchids, Kapoho (19°29′50.97″ N, 154 °57′03.03″ W, 190 m AMSL) Kohala (20°14′15.25″ N, 155°49′07.74″ W, 158 m AMSL) Onomea (19°48′31.67″ N, 155°05′45.24″ W, 87 m AMSL) Akaka Falls (19°51′14.25″ N, 155°09′07.47″ W, 366 m AMSL) Panaewa, Umauma (19°39′34.28″ N, 155°02′50.93″ W, 57 m AMSL) | 31 (50–100) | 2725 |
| “ | Hilo, Puna, districts 2011 | 27 January 2011– 8 February 2011 | Pahua (19″ 27′41.80″ N, 154″ 56′25.71 W, 294 m) Panaewa, (19°39′34.28″ N, 155°02′50.93″ W, 57 m AMSL) Kurtistown, (19°35′34.87″ N, 155°03′27.95″ W, 200 m AMSL) | 4 (50) | 200 |
| Maui Island | North shore and East Maui 2010 | 3 August 2010– 23 November 2010 | Haiku (20°55′02.87″ N, 156°19′32.93″ W, 144 m AMSL) Hana, Maliko Gulch (20°55′54.48″ N, 156°20′19.66″ W, 11 m AMSL) | 9 (50–200) | 910 |
| “ | North Shore 2011 | 25 February 2011– 26 August 2011 | Twin Falls off, Hana. (20°54′43.80″ N, 156°14′34.31″ W, 148 m AMSL) | 6 (50–150) | 640 |
| Kauai Island | East and North districts 2010 | 19 October 2010 | Kuamoo Rd, Kapaa (22°03′27.94″ N, 159°22′52.16″ W, 98 m AMSL) Kiluea, Kauai Orchids (22°11′51.05″ N, 159°22′34.09″ W, 109 m AMSL) | 5 (15–100) | 200 |
| “ | East district 2011 | 5 May 2011– 30 August 2011 | Kapaa, Transfer Station (22°05′00.14″ N, 159°19′26.43″ W, 21 m AMSL) | 15 (25–100) | 950 |
| Total releases and mean/locality | 162 (release sites) | 13,379 836.2 ± 158.8 | |||
| Date | Locality | No. of Samples | Total Oxyplax Larvae Collected | Oxyplax Larvae/Sample Mean ± SEM | Total Larvae Parasitized by Aroplectrus | % Parasitism by Aroplectrus | Total Larvae with Hyper Parasitoid | % Parasitism by Pediobius |
|---|---|---|---|---|---|---|---|---|
| 10 September 2019 | Enchanted lake, Kailua | 1 | 52 | 52 | 0 | 0 | - | - |
| 24 September 2009– 23 December 2019 | Lanikai | 12 | 812 | 67.7 ± 8.9 | 52 | 6.4 | 5 | 9.6 |
| 23 October 2019– 10 December 2019 | Waimanalo | 3 | 125 | 41.7 ± 7.9 | 10 | 8.0 | 1 | 10.0 |
| 27 November 2019 | Kipapa Gulch | 1 | 1 | 1 | 0 | - | - | - |
| 27 January 2020– 18 October 2020 | Lanikai | 4 | 77 | 19.3 ± 4.1 | 16 | 20.8 | 4 | 25.0 |
| 28 July 2020 | Kailua | 1 | 20 | 20 | 0 | - | - | - |
| 16 October 2020– 16 December 2020 | Maunawili Loop, Kailua | 3 | 261 | 87.0 ± 27.2 | 2 | 0.8 | 2 | 100 |
| 23 January 2020 | Waimanalo | 3 | 54 | 18.0 ± 7.2 | 5 | 9.2 | 5 | 100 |
| 4 January 2021– 26 July 2021 | Haleiwa | 4 | 47 | 11.7 ± 6.7 | 3 | 6.4 | 1 | 33.3 |
| 10 March 2021 | Waimanalo | 7 | 58 | 8.3 ± 3.8 | 38 | 65.5 | 5 | 13.1 |
| 7 April 2021– 11 August 2021 | Maunawili, Kailua | 6 | 51 | 8.5 ± 3.0 | 0 | 0 | 0 | - |
| 7 April 2021– 20 October 2021 | Lanikai | 11 | 314 | 28.5 ± 5.9 | 23 | 7.3 | 0 | 0 |
| 5 August 2021– 11 November 2021 | Olomana | 6 | 466 | 77.7 ± 17.9 | 36 | 7.7 | 0 | 0 |
| 14 June 2021– 16 December 2021 | Enchanted Lake, Kailua | 6 | 417 | 69.5 ± 25.3 | 26 | 6.2 | 0 | 0 |
| 19 January 2022– 15 October, 2022 | Enchanted lake, Kailua | 6 | 41 | 6.8 ± 1.8 | 18 | 43.9 | 2 | 11.1 |
| 23 February 2022– 6 December 2022 | Maunawili | 2 | 24 | 12.0 ± 8.0 | 0 | 0 | 0 | - |
| 11 January 2022 – 20 December 2022 | Lanikai | 8 | 239 | 29.9 ± 7.0 | 26 | 10.9 | 1 | 3.8 |
| 19 January 2022– 26 September 2022 | Olomana | 8 | 262 | 32.7 ± 7.7 | 121 | 46.2 | 65 | 53.7 |
| 9 March 2022– 12 July 2022 | Waimanalo | 2 | 2 | 2 | 2 | 100 | 0 | 0 |
| 10 November 2022 | kaillua | 1 | 13 | 13 | 0 | |||
| 10 January 2023– 23 October 2023 | Lanikai | 8 | 248 | 31.0 ± 9.1 | 12 | 4.8 | 1 | 8.3 |
| 18 January 2023– 31 October 2023 | Olomana | 5 | 125 | 25.0 ± 7.0 | 70 | 56.0 | 27 | 38.6 |
| 18 January 2023– 28 July 2023 | Enchanted lake, Kailua | 2 | 5 | 2.5 ± 0.5 | 0 | 0 | 0 | |
| 28 April 2023– 23 August 2022 | Mililani | 4 | 59 | 14.7 ± 6.4 | 5 | 8.5 | 3 | 60.0 |
| 6 February 2023– 21 August 2023 | Waimanalo | 2 | 50 | 25.0 ± 13.0 | 4 | 8.0 | 1 | 25.0 |
| Mean ± SEM /site | 28.22 ± 4.9 | 18.76 ± 5.6 | 18.93 ± 5.6 | 5.86 ± 3.2 | 27.30 ± 7.6 | |||
| All samples | 117 samples (25 sites) | 3923 larvae | 469 larvae | 123 larvae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yalemar, J.A.; Nagamine, W.T.; Bautista, R.C.; Cho, D.Y.; Nakahara, L.M.; Ramadan, M.M. Aroplectrus dimerus (Hymenoptera: Eulophidae), Ectoparasitoid of the Nettle Caterpillar, Oxyplax pallivitta (Lepidoptera: Limacodidae): Evaluation in the Hawaiian Islands. Life 2024, 14, 509. https://doi.org/10.3390/life14040509
Yalemar JA, Nagamine WT, Bautista RC, Cho DY, Nakahara LM, Ramadan MM. Aroplectrus dimerus (Hymenoptera: Eulophidae), Ectoparasitoid of the Nettle Caterpillar, Oxyplax pallivitta (Lepidoptera: Limacodidae): Evaluation in the Hawaiian Islands. Life. 2024; 14(4):509. https://doi.org/10.3390/life14040509
Chicago/Turabian StyleYalemar, Juliana A., Walter T. Nagamine, Renato C. Bautista, Dexter Y. Cho, Larry M. Nakahara, and Mohsen M. Ramadan. 2024. "Aroplectrus dimerus (Hymenoptera: Eulophidae), Ectoparasitoid of the Nettle Caterpillar, Oxyplax pallivitta (Lepidoptera: Limacodidae): Evaluation in the Hawaiian Islands" Life 14, no. 4: 509. https://doi.org/10.3390/life14040509