
The Anti-Cholinesterase Potential of Fifteen Different Species of Narcissus L. (Amaryllidaceae) Collected in Spain

 ,
, 
 ,
,  , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Alkaloid Extracts Preparation
2.3. Cholinesterase Inhibitory Activity
2.4. Statistical Evaluation
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bremer, B.; Bremer, K.; Chase, M.W.; Fay, M.F.; Reveal, J.L.; Bailey, L.H.; Soltis, D.E.; Soltis, P.S.; Stevens, P.F.; Anderberg, A.A.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Hanks, G.R. (Ed.) Narcissus and Daffodil: The Genus Narcissus; vol. 21 in the series: ‘‘Medicinal and Aromatic Plants—Industrial Profiles; Taylor & Francis: London, UK, 2002; pp. 1–428. [Google Scholar]
- Bastida, J.; Lavilla, R.; Viladomat, F. Chemical and Biological Aspects of Narcissus Alkaloids. In The Alkaloids: Chemistry and Biology; Cordell, G.A., Ed.; Elsevier: San Diego, CA, USA, 2006; Volume 63, pp. 87–179. [Google Scholar]
- Berkov, S.; Martínez-Francés, V.; Bastida, J.; Codina, C.; Ríos, S. Evolution of alkaloid biosynthesis in the genus Narcissus. Phytochemistry 2014, 99, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.W.; Barrett, S.C.H. Phylogenetic reconstruction of the evolution of stylar polymorphisms in Narcissus (Amaryllidaceae). Am. J. Bot. 2004, 91, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Lisa-Molina, J.; Gómez-Murillo, P.; Arrelano-Martín, I.; Jiménez, C.; Rodríguez-Escobar, M.L.; Tallini, L.R.; Viladomat, F.; Torras-Claveria, L.; Bastida, J. Alkaloid profile in wild autumn-flowering daffodils and their acetylcholinesterase inhibitory activity. Molecules 2023, 28, 1239. [Google Scholar] [CrossRef] [PubMed]
- Rivera Núñez, D.; Obón de Castro, C.; Ríos Ruiz, S.; Alcaraz Ariza, F. The origin of cultivation and wild ancestors of daffodils (Narcissus subgenus Ajax) from an analysis of early illustrations. Sci. Hortic. 2003, 98, 307–330. [Google Scholar] [CrossRef]
- Marques, I.; Aguilar, J.F.; Martins-Louçao, M.A.; Moharrek, F.; Feliner, G.N. A three-genome five-gene comprehensive phylogeny of the bulbous genus Narcissus (Amaryllidaceae) challenges current classifications and reveals multiple hybridization events. Taxon 2017, 66, 832–854. [Google Scholar] [CrossRef]
- Berkov, S.; Georgieva, L.; Kondakova, V.; Atanassov, A.; Viladomat, F.; Bastida, J.; Codina, C. Plant sources of galanthamine: Phytochemical and biotechnological aspects. Biotechnol. Biotec. Eq. 2014, 23, 1170–1176. [Google Scholar] [CrossRef]
- Berkov, S.; Ivanov, I.; Georgiev, V.; Codina, C.; Pavlov, A. Galanthamine biosynthesis in plant in vitro systems. Eng. Life Sci. 2014, 14, 643–650. [Google Scholar] [CrossRef]
- van Goietsenoven, G.; Mathieu, V.; Lefranc, F.; Kornienko, A.; Evidente, A.; Kiss, R. Narciclasine as well as other Amaryllidaceae isocarbostyrils are Promising GTP-ase targeting agents against brain cancers. Med. Res. Rev. 2013, 33, 439–455. [Google Scholar] [CrossRef]
- Dioscorides, P. Acerca de la Materia Medicinal, y de los Venenos Mortiferos, Traducido de la Lengua Griega, en la Vulgar Castellana, & Illustrado con Claras y Substantiales Annotationes, y con las Figuras de Innumeras Plantas Exquisitas y Raras/Ror el Doctor Andres de Laguna, Medico de Iulio III. Pont. Maxi; Gast, M., Ed.; Real Jardín Botánico-CSIC: Salamanca, Spain, 1563; Available online: https://bibdigital.rjb.csic.es/records/item/13529-redirection (accessed on 7 February 2024).
- Maelicke, A.; Samochocki, M.; Jostock, R.; Fehrenbacher, A.; Ludwig, J.; Albuquerque, E.X.; Zerlin, M. Allosteric sensitization of nicotinic receptors by galantamine, a new treatment strategy for Alzheimer’s disease. Biol. Psychiatry 2001, 49, 279–288. [Google Scholar] [CrossRef]
- Heinrich, M.; Teoh, H.L. Galanthamine from snowdrop—The development of a modern drug against Alzheimer’s disease fromlocal Caucasian knowledge. J. Ethnopharmacol. 2004, 92, 147–162. [Google Scholar] [CrossRef]
- Berkov, S.; Georgieva, L.; Sidjimova, B.; Bastida, J. Evaluation of Hippeastrum papilio (Ravenna) Van Scheepen potencial as a new industrial source of galanthamine. Ind. Crop. Prod. 2022, 178, 114619. [Google Scholar] [CrossRef]
- Lubbe, A.; Pomahačová, B.; Choi, Y.H.; Verpoorte, R. Analysis of metabolic variation and galanthamine content in Narcissus bulbs by 1H NMR. Phytochem. Anal. 2010, 21, 66–72. [Google Scholar] [CrossRef]
- Lubbe, A.; Verpoorte, R.; Choi, Y.H. Effects of fungicides on galanthamine and metabolite profiles in Narcissus bulbs. Plant Physiol. Biochem. 2012, 58, 116–123. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Daffodils as potential crops of galanthamine. Assessment of more than 100 ornamental varieties for their alkaloid content and acetylcholinesterase inhibitory activity. Ind. Crop. Prod. 2013, 43, 237–244. [Google Scholar] [CrossRef]
- Berkov, S.; Osorio, E.; Viladomat, F.; Bastida, J. Chemodiversity, chemotaxonomy and chemoecology of Amaryllidaceae alkaloids. In The Alkaloids: Chemistry and Biology; Knölker, H.-J., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 83, pp. 113–185. [Google Scholar] [CrossRef]
- Battersby, A.R.; Fales, H.M.; Wildman, W.C. Biosynthesis in the Amaryllidaceae. Tyrosine and norbelladine as precursors of haemanthamine. J. Am. Chem. Soc. 1961, 83, 4098–4099. [Google Scholar] [CrossRef]
- Kilgore, M.B.; Augustin, M.M.; Starks, C.M.; O’Neil-Johnson, M.; May, G.D.; Crow, J.A.; Kutchan, T.M. Cloning and characterization of a norbelladine 4′-O-methyltransferase involved in the biosynthesis of the Alzheimer’s drug galanthamine in Narcissus sp. aff. pseudonarcissus. PLoS ONE 2014, 9, e103223. [Google Scholar] [CrossRef] [PubMed]
- Barton, D.H.R.; Cohen, T. Some biogenic aspects of phenol oxidation. In Festschrift Arthur Stoll; Birkhauser, A.G., Ed.; Birkhauser: Basel, Switzerland, 1956; pp. 117–143. [Google Scholar]
- Rivera, D.; Ríos, S.; Obón, C.; Alcaraz, F. The image of daffodils in art and botanical illustration: Clues to the History of domestication and selection of Narcissus subgenus Ajax (Amaryllidaceae). In Floriculture, Ornamental and Plant Biotechnology; Teixeira, J., Ed.; Global Science Books: Ikenobe, Japan, 2006; Volume IV, Chapter 39; pp. 360–369. [Google Scholar]
- Rivera, D.; Ríos, S.; Alcaraz, F.; Obón, C. The biogeographical patterns of floral form in wild daffodils and their contribution to the cultivar groups of Narcissus L. Subgenus Ajax Spach (Amaryllidaceae). In Floriculture, Ornamental and Plant Biotechnology; Teixeira, J., Ed.; Global Science Books: Ikenobe, Japan, 2006; Volume IV, Chapter 32; pp. 300–313. [Google Scholar]
- Rivera, D.; Ríos, S.; Alcaraz, F.; Obón, C.; Verde, A.; Fajardo, J. The biogeography and habitat selection of Ajax daffodils (Narcissus L. subgenus Ajax Spach, Amaryllidaceae) in the Iberian Peninsula. In Frontiers of Vegetation Science—An Evolutionary Angle; Mucina, L., Kalwij, J., Smith, V.R., Chytrỳ, M., White, P.S., Ciliers, S.S., Pillar, V.D., Zobel, M., Sun, I.-F., Eds.; Keith Phillips Images: Somerset West, South Africa, 2008; pp. 152–153. [Google Scholar]
- Ríos, S.; Juan, J.; Martínez-Francés, V.; Laguna, E.; Rivera, D.; Alcaraz, F.; Verde, A.; Fajardo, J.; Carreño, E.; Casas, J.L.; et al. Endemic species of Narcissus in Central Spain: Biodiversity and conservation under grazing pressure by wild and domestic herbivorous. In Options Mediterranéenes Series A; Porqueddu, S., Ríos, S., Eds.; CIHEAM: Zaragoza, Spain, 2010; Volume 92, pp. 79–83. [Google Scholar]
- Blanca, G.; López, M.R.; Lorite, J.; Martínez, M.J.; Molero-Mesa, J.; Quintas, S.; Ruíz, M.; Varo, M.A.; Vidal, S. Flora amenazada y endémica de Sierra Nevada; Universidad de Granada: Granada, Spain, 2002; p. 407. [Google Scholar]
- Gutiérrez, L.; Lorite, J.; Ruiz, M.; López, M.; Donaire, F. Narcissus nevadensis Pugsley subsp. nevadensis. In Atlas y Libro Rojo de la Flora Vascular Amenazada de España; Bañares, A., Blanca, G., Güemes, J., Moreno, J.C., Ortiz, S., Eds.; Dirección General de Conservación de la Naturaleza: Madrid, Spain, 2003; pp. 796–797. [Google Scholar]
- Moreno Saiz, J.C.; Sainz ollero, H. Atlas corológico de las monocotiledóneas endémicas de la Península Ibérica e Islas Baleares. Bases para una política de conservación. Colección Técnica. ICONA; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1992; p. 354. [Google Scholar]
- Hernández-Bermejo, E.; Prados, J.; Herrera-Molina, F.; Narcissus bugei. The IUCN Red List of Threatened Species 2006: E.T61601A12519064. Available online: https://www.iucnredlist.org/species/61601/12519064 (accessed on 7 February 2024).
- García Murillo, P.G.; Ríos Ruiz, S.; Gutiérrez Carretero, L.; Lorite, J.; Ruiz Girela, M.; López, M.; Donaire Sánchez, F.J.; Sánchez Gómez, P.; Carrión Vilches, M.Á.; Navarro, F.B.; et al. Narcissus nevadensis. The IUCN Red List of Threatened Species 2018: E.T61682A47061845. Available online: https://www.iucnredlist.org/species/61682/47061845 (accessed on 7 February 2024).
- Juan Vicedo, J.; Ríos Ruiz, S.; García Murillo, P.G. Narcissus alcaracensis. The IUCN Red List of Threatened Species 2018: E.T61600A57488304. Available online: https://www.iucnredlist.org/species/61600/57488304 (accessed on 7 February 2024).
- Juan Vicedo, J.; Narcissus yepesii. The IUCN Red List of Threatened Species 2018: E.T57456144A57456147. Available online: https://www.iucnredlist.org/species/57456144/57456147 (accessed on 7 February 2024).
- Caldas, F.B.; Moreno Saiz, J.C.; Narcissus asturiensis. The IUCN Red List of Threatened Species 2013: E.T162098A5536148. Available online: https://www.iucnredlist.org/species/162098/5536148 (accessed on 7 February 2024).
- Juan Vicedo, J.; Narcissus hedraeanthus. The IUCN Red List of Threatened Species 2018: E.T57455245A57455282. Available online: https://www.iucnredlist.org/species/57455245/57455282 (accessed on 7 February 2024).
- Caldas, F.B.; Moreno Saiz, J.C.; Buord, S.; Narcissus triandrus. The IUCN Red List of Threatened Species 2013: E.T162050A5542491. Available online: https://www.iucnredlist.org/species/162050/5542491 (accessed on 7 February 2024).
- Pigni, N.B.; Ríos-Ruiz, S.; Martínez-Francés, V.; Nair, J.J.; Viladomat, F.; Codina, C.; Bastida, J. Alkaloids from Narcissus serotinus. J. Nat. Prod. 2012, 75, 1643–1647. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Metabolomic analysis of bioactive Amaryllidaceae alkaloids of ornamental varieties of Narcissus by GC-MS combined with k-means cluster analysis. Ind. Crop. Prod. 2014, 56, 211–222. [Google Scholar] [CrossRef]
- Takos, A.M.; Rook, F. Towards a molecular understanding of the biosynthesis of Amaryllidaceae alkaloids in support of their expanding medical use. Int. J. Mol. Sci. 2013, 14, 11713. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–90. [Google Scholar] [CrossRef] [PubMed]
- López, S.; Bastida, J.; Viladomat, F.; Codina, C. Acetylcholinesterase inhibitory activity of some Amaryllidaceae alkaloids and Narcissus extracts. Life Sci. 2002, 71, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.; Sener, B. Bioactivity-directed fractionation of alkaloids from some Amaryllidaceae plants and their anticholinesterase activity. Chem. Nat. Compd. 2003, 39, 383–386. [Google Scholar] [CrossRef]
- Havlasová, J.; Šafratová, M.; Siatka, T.; Štěpánková, S.; Novák, Z.; Ločárek, M.; Opletal, L.; Hrabinová, M.; Jun, D.; Benešová, N.; et al. Chemical Composition of Bioactive Alkaloid Extracts from Some Narcissus Species and Varieties and their Biological Activity. Nat. Prod. Commun. 2014, 9, 1151–1155. [Google Scholar] [CrossRef]
- Gotti, R.; Fiori, J.; Bartolini, M.; Cabrini, V. Analysis of Amaryllidaceae alkaloids from Narcissus by GC-MS and capillary electrophoresis. J. Pharmaceut. Biomed. 2006, 42, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Frolova, L.; Yu, X.; Mathieu, V.; Cimmino, A.; De Carvalho, A.; Kiss, R.; Rogelj, S.; Pertsemlidis, A.; Kornienko, A.; et al. Jonquailine, a new pretazettine-type alkaloid isolated from Narcissus jonquilla quail, with activity against drug-resistant cancer. Fitoterapia 2015, 102, 41–48. [Google Scholar] [CrossRef]
- Bozkurt, B.; Coban, G.; Kaya, G.I.; Onur, M.A.; Unver-Somer, N. Alkaloid profiling, anticholinesterase activity and molecular docking of Galanthus elwesii. S. Afr. J. Bot. 2017, 113, 119–127. [Google Scholar] [CrossRef]
- Cortes, N.; Sierra, K.; Alzate, F.; Osorio, E.H.; Osorio, E. Alkaloids of Amaryllidaceae as Inhibittors of Cholinesterases (AChE and BuChEs): An Integrated Bioguide Study. Phytochem. Anal. 2017, 29, 217–227. [Google Scholar] [CrossRef] [PubMed]
- León, K.A.; Inca, A.; Tallini, L.R.; Osorio, E.H.; Robles, J.; Bastida, J.; Oleas, N.H. Alkaloids of Phaedranassa dubia (Kunth) J.F. Macbr. and Phaedranassa brevifolia Meerow (Amaryllidaceae) from Ecuador and its cholinesterase-inhibitory activitty. S. Afr. J. Bot. 2021, 136, 91–99. [Google Scholar] [CrossRef]
- Rojas-Vera, J.d.C.; Buitrago-Díaz, A.A.; Possamai, L.M.; Timmers, L.F.S.M.; Tallini, L.R.; Bastida, J. Alkaloid profile and cholinesterase inhibition activity of five species of Amaryllidaceae family collected from Mérida state-Venezuela. S. Afr. J. Bot. 2021, 136, 126–136. [Google Scholar] [CrossRef]
- Tallini, L.R.; Osorio, E.H.; Berkov, S.; Torras-Claveria, L.; Rodríguez-Escobar, M.L.; Viladomat, F.; Meerow, A.; Bastida, J. Chemical Survey of Three Species of the Genus Rauhia Traub (Amaryllidaceae). Plants 2022, 11, 3549. [Google Scholar] [CrossRef] [PubMed]
- Llabrés, J.M.; Viladomat, F.; Bastida, J.; Codina, C.; Rubiralta, M. Phenanthridine alkaloids from Narcissus assoanus. Phytochemistry 1986, 25, 2637–2638. [Google Scholar] [CrossRef]
- Viladomat, F.; Llabrés, J.M.; Bastida, J.; Cusidó, R.M.; Codina, C. Ontogenic variations in the alkaloids of Narcissus assoanus. Physiol. Plant. 1986, 68, 657–661. [Google Scholar] [CrossRef]
- Bastida, J.; Viladomat, F.; Llabrés, J.M.; Codina, C.; Rubiralta, M. Alkaloids from Narcissus jacetanus. Planta Med. 1988, 54, 362. [Google Scholar] [CrossRef]
- Tallini, L.R.; Bastida, J.; Cortes, N.; Osorio, E.H.; Theoduloz, C.; Schmeda-Hirschmann, G. Cholinesterase inhibition activity, alkaloid profiling and molecular docking of Chilean Rhodophiala (Amaryllidaceae). Molecules 2018, 23, 1532. [Google Scholar] [CrossRef] [PubMed]
- Brito, M.F.B.; Ferreira, J.V.; de Souza, L.R.; Gemaque, L.R.P.; Sousa, K.P.A.; dos Santos, C.F.; Braga, F.S.; Pernomian, L.; da Silva, C.H.T.P.; Santos, C.B.R.; et al. Computational Molecular Modeling of Compounds from Amaryllidaceae Family as Potential Acetylcholinesterase Inhibitors. Curr. Bioact. Compd. 2017, 13, 121–129. [Google Scholar] [CrossRef]
- Labraña, J.; Choy, G.; Solans, X.; Font-Bardia, M.; de la Fuente, G.; Viladomat, F.; Codina, C.; Bastida, J. Alkaloids from Narcissus bujei (Amaryllidaceae). Phytochemistry 1999, 50, 183–188. [Google Scholar] [CrossRef]
- Bastida, J.; Codina, C.; Viladomat, F. Narcissus alkaloids, XIV. (+)-8-O-Acetylhomolycorine and vasconine, two novel alkaloids from Narcissus vasconicus. J. Nat. Prod. 1992, 55, 122–125. [Google Scholar] [CrossRef]
- Pigni, N.B.; Ríos-Ruiz, S.; Luque, L.; Viladomat, F.; Codina, C.; Bastida, J. Wild daffodils of the section Ganymedes from the Iberian Peninsula as a source of mesembrane alkaloids. Phytochemistry 2013, 95, 384–393. [Google Scholar] [CrossRef]
- Evidente, A. Advances on the Amaryllidaceae alkaloids collected in South Africa, Andean South America and the Mediterranean basin. Molecules 2023, 28, 4055. [Google Scholar] [CrossRef]
- Seijas, J.A.; Vázquez-Tato, M.P.; Linares, M.T.; Ramil-Rego, P.; Buján, M.I. Mesembrine alkaloids from Narcissus triandrus L. In Proceedings of ECSOC-8, The Eight International Electronic Conference on Synthetic Organic Chemistry; Seijas, J.A., Vázquez-Tato, M.P., Eds.; MDPI: Basel, Switzerland,, 2004; Available online: https://www.mdpi.org/ecsoc/ecsoc-8/BOCNP/008/index.htm (accessed on 7 February 2024).
- Bastida, J.; Viladomat, F.; LLabres, J.M.; Codina, C.; Feliz, M.; Rubiralta, M. Alkaloids from Narcissus confusus. Phytochemistry 1987, 26, 1519–1524. [Google Scholar] [CrossRef]
- Bastida, J.; Llabrés, J.M.; Viladomat, F.; Codina, C. Narcissus alkaloids, III. 9-O-demethylhomolycorine from Narcissus confusus. J. Nat. Prod. 1987, 50, 199–202. [Google Scholar] [CrossRef]
- Berkov, S.; Bastida, J.; Viladomat, F.; Codina, C. Development and validation of a GC-MS method for rapid determination of galanthamine in Leucojum aestivum and Narcissus ssp.: A metabolic approach. Talanta 2011, 83, 1455–1465. [Google Scholar] [CrossRef]
- Karakoyun, Ç.; Ünver-Somer, N. Simultaneous quantitative analysis of biologically important Amaryllidaceae alkaloids in Narcissus tazetta L. subsp. tazetta by HPLC/PDA. J. Res. Pharm. 2019, 23, 498–505. [Google Scholar] [CrossRef]
- Karakoyun, Ç.; Bozkurt, B.; Çoban, G.; Masi, M.; Cimmino, A.; Evidente, A.; Somer, N.U. A comprehensive study on Narcissus tazetta subsp. tazetta L.: Chemo-profiling, isolation, anticholinesterase activity and molecular docking of Amaryllidaceae alkaloids. S. Afr. J. Bot. 2020; 130, 148–154. [Google Scholar] [CrossRef]
- Tarakemeh, A.; Azizi, M.; Rowshan, V.; Salehi, H.; Spina, R.; Dupire, F.; Arouie, H.; Laurain-Mattar, D. Screening of Amaryllidaceae alkaloids in bulbs and tissue cultures of Narcissus papyraceus and four varieties of N. tazetta. J. Pharmaceut. Biomed. 2019, 172, 230–237. [Google Scholar] [CrossRef]
- Viladomat, F.; Sellés, M.; Codina, C.; Bastida, J. Alkaloids from Narcissus asturiensis. Planta Med. 1997, 63, 583. [Google Scholar] [CrossRef]
- Mamun, A.A.; Maríková, J.; Hulcová, D.; Janoušek, J.; Šafratová, M.; Nováková, L.; Kucera, T.; Hrabinová, M.; Kuneš, J.; Korábecny, J.; et al. Amaryllidaceae Alkaloids of belladine-type from Narcissus pseudonarcissus cv. Carlton as new selective inhibitors of butyrylcholinesterase. Biomolecules 2020, 10, 800. [Google Scholar] [CrossRef]
- Hulcová, D.; Mariková, J.; Korábecny, J.; Hostalková, A.; Jun, D.; Kuneš, J.; Chlebek, J.; Opletal, L.; De Simone, A.; Nováková, L.; et al. Amaryllidaceae alkaloids from Narcissus pseudonarcissus L. cv. Dutch Master as potential drugs in treatment of Alzheimer’s disease. Phytochemistry 2019, 165, 112055. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
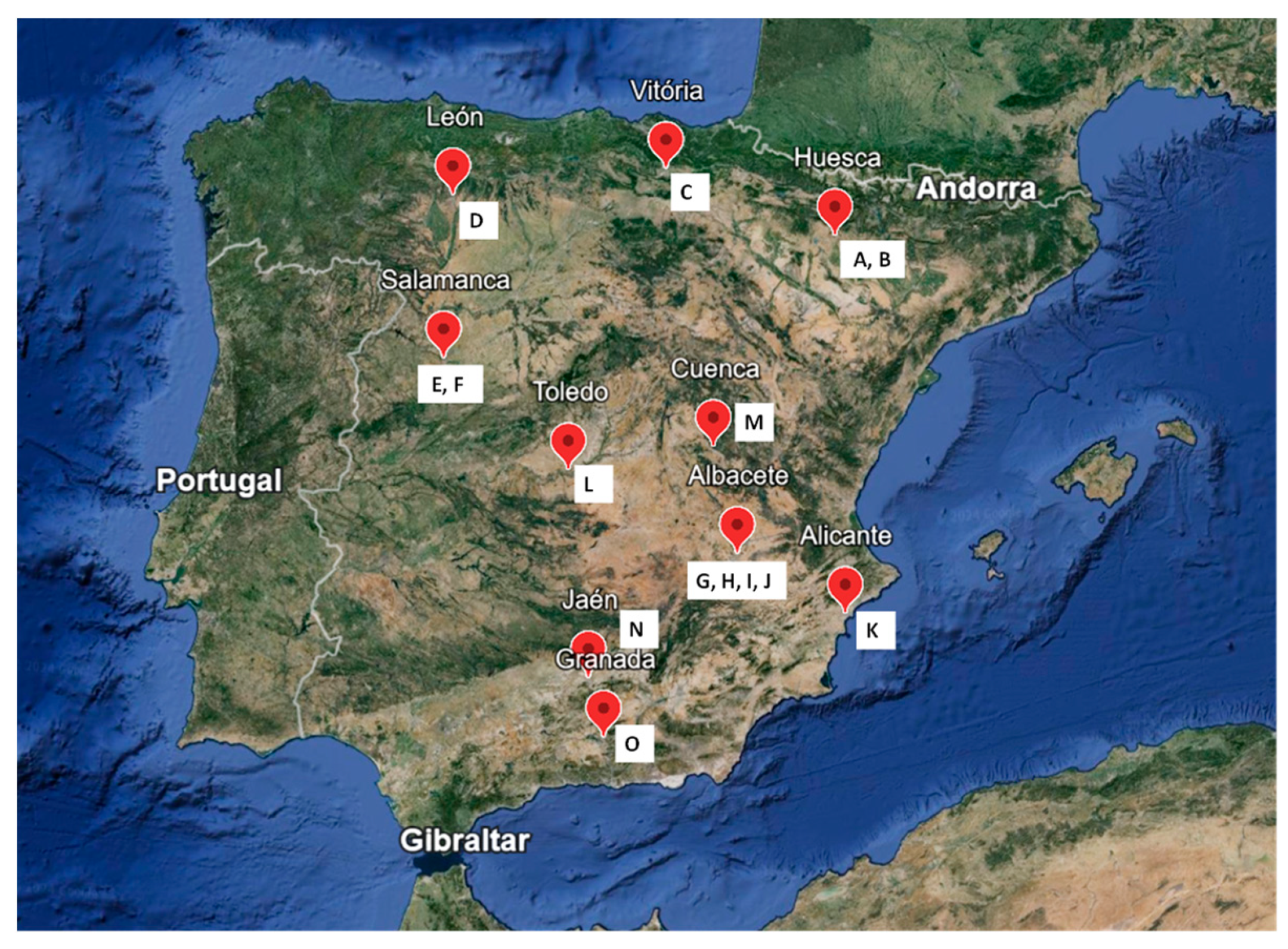
| Code | Species | Section | Biogeographical Distribution | Origin |
|---|---|---|---|---|
| A | N. assoanus Dufour ex Schult. and Schult.f. | Jonquillae DC. | Med Occ | Huesca (AR) |
| B | N. jacetanus Fern.Casas | Pseudonarcissus DC. | Iber/LAlp | Huesca (AR) |
| C | N. vasconicus (Fern.Casas) Fern.Casas | Pseudonarcissus DC. | Iber/LAlp | Vitoria (BAS) |
| D | N. minor L. | Pseudonarcissus DC. | Iber/LAlp | León (CL) |
| E | N. confusus Pugsley | Pseudonarcissus DC. | Iber/LAlp | Salamanca (CL) |
| F | N. asturiensis (Jord.) Pugsley | Pseudonarcissus DC. | Iber/LAlp | Salamanca (CL) |
| G | N. hedraeanthus (Webb and Heldr.) Colmeiro | Bulbocodium (Salisb.) DC. | Iber/NAfric | Albacete (CLM) |
| H | N. alcaracensis S.Ríos, D.Rivera, Alcaraz and Obón | Nevadensis Zonn. | Baet | Albacete (CLM) |
| I | N. bujei (Fern.Casas) Fern.Casas | Pseudonarcissus DC. | Baet | Albacete (CLM) |
| J | N. pallidulus Graells | Ganymedes (Salisb.) Schultes f. | Iber | Albacete (CLM) |
| K | N. tazetta L. | Tazettae DC. | CircMed | Alicante (VAL) |
| L | N. jonquilla L. | Jonquillae DC. | Iber | Toledo (CLM) |
| M | N. genesii-lopezii Fern.Casas | Pseudonarcissus DC. | Iber | Cuenca (CLM) |
| N | N. yepesii S.Ríos, D.Rivera, Alcaraz and Obón | Nevadensis Zonn. | Baet | Jaén (AND) |
| O | N. nevadensis Pugsley | Nevadensis Zonn. | Baet | Granada (AND) |
| Samples | Dry Weight (g) | Alkaloid Extract (mg) | Yield (%) |
|---|---|---|---|
| A | 1.00080 | 8.93 | 0.89 |
| B | 1.00114 | 4.69 | 0.47 |
| C | 1.00046 | 6.05 | 0.60 |
| D | 1.00050 | 4.95 | 0.49 |
| E | 1.00032 | 14.54 | 1.45 |
| F | 1.00035 | 9.79 | 0.98 |
| G | 0.88253 | 6.83 | 0.77 |
| H | 1.00069 | 10.66 | 1.07 |
| I | 1.00125 | 18.83 | 1.88 |
| J | 1.00137 | 10.17 | 1.02 |
| K | 1.00103 | 8.38 | 0.84 |
| L | 1.00094 | 15.32 | 1.53 |
| M | 1.00099 | 6.85 | 0.68 |
| N | 1.00037 | 61.43 | 6.14 |
| O | 1.00066 | 9.27 | 0.93 |
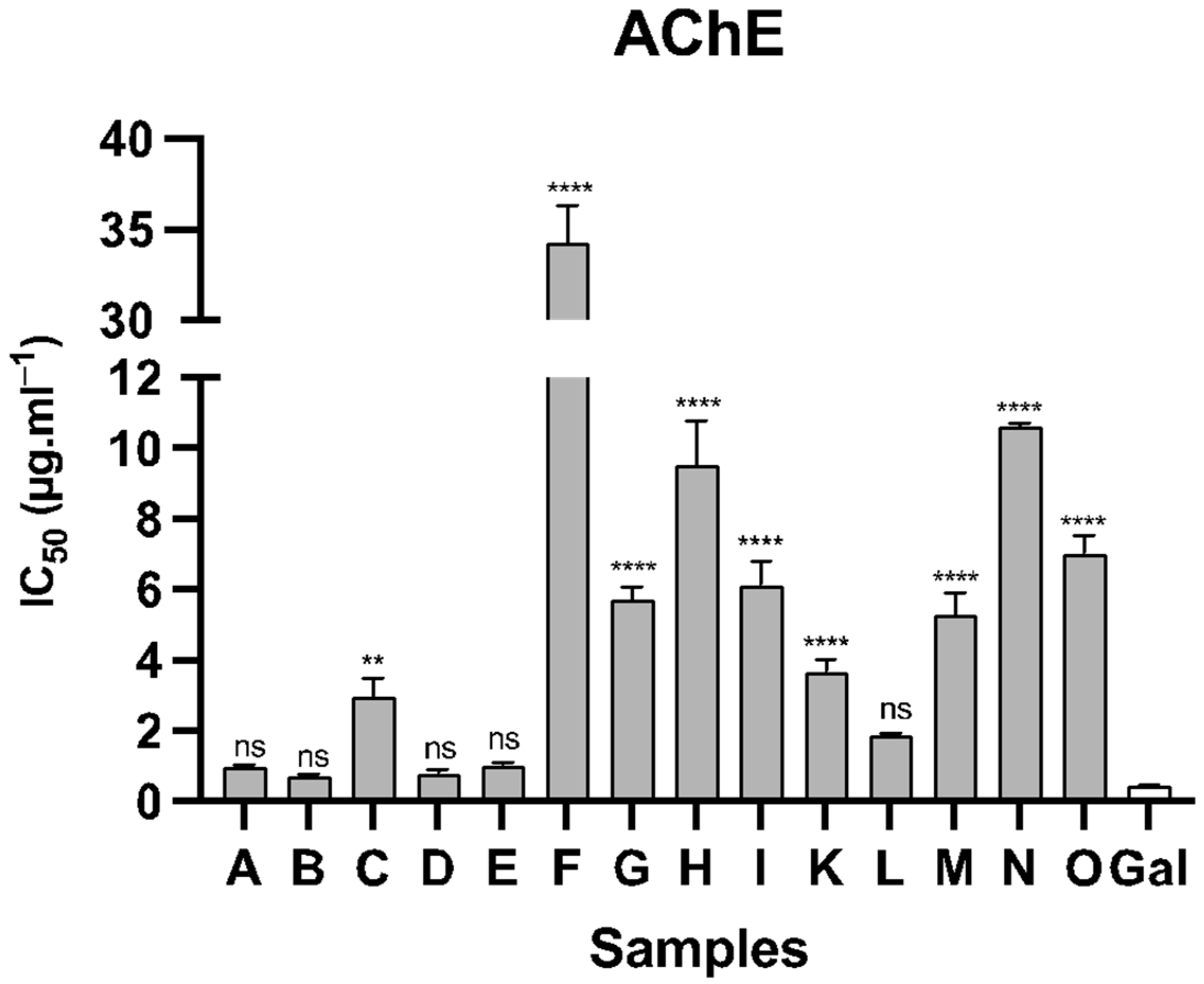
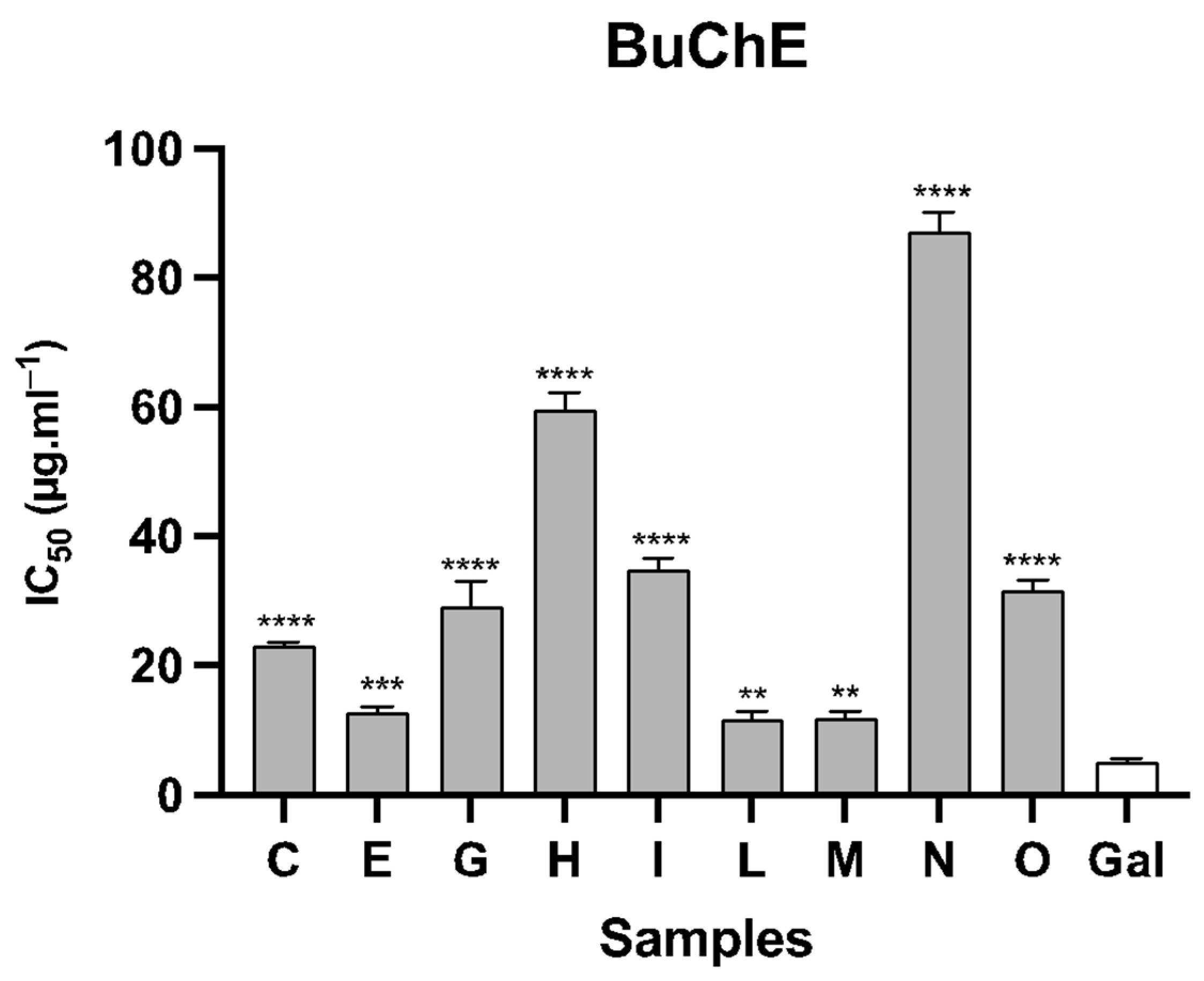
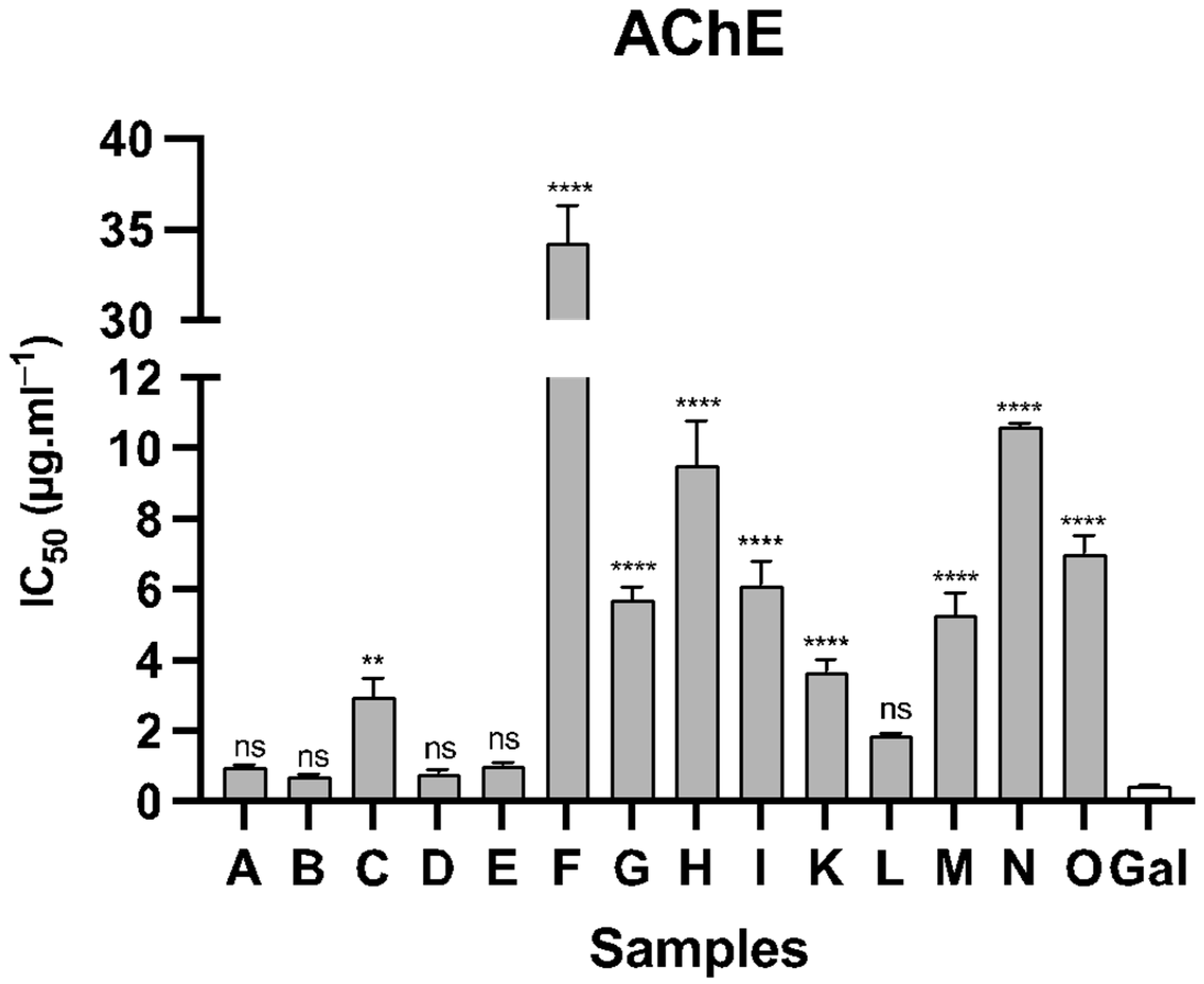
| Samples | Species | AChE | BuChE |
|---|---|---|---|
| A | N. assoanus | 0.99 ± 0.06 | >100 |
| B | N. jacetanus | 0.75 ± 0.03 | >100 |
| C | N. vasconicus | 2.98 ± 0.51 | 23.14 ± 0.58 |
| D | N. minor | 0.81 ± 0.10 | >100 |
| E | N. confusus | 1.04 ± 0.07 | 12.83 ± 0.87 |
| F | N. asturiensis | 34.28 ± 2.06 | >100 |
| G | N. hedraeanthus | 5.73 ± 0.36 | 29.23 ± 3.84 |
| H | N. alcaracencis | 9.54 ± 1.26 | 59.60 ± 2.61 |
| I | N. bujei | 6.14 ± 0.68 | 34.86 ± 1.75 |
| J | N. pallidulus | >100 | >100 |
| K | N. tazetta | 3.68 ± 0.36 | >100 |
| L | N. jonquilla | 1.88 ± 0.05 | 11.72 ± 1.15 |
| M | N. genesii-lopezii | 5.28 ± 0.64 | 11.98 ± 0.93 |
| N | N. yepesii | 10.63 ± 0.08 | 87.20 ± 3.01 |
| O | N. nevadensis | 7.03 ± 1.49 | 31.68 ± 0.49 |
| Gal | 0.46 ± 0.03 | 5.13 ± 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tallini, L.R.; Manfredini, G.; Rodríguez-Escobar, M.L.; Ríos, S.; Martínez-Francés, V.; Feresin, G.E.; Borges, W.d.S.; Bastida, J.; Viladomat, F.; Torras-Claveria, L. The Anti-Cholinesterase Potential of Fifteen Different Species of Narcissus L. (Amaryllidaceae) Collected in Spain. Life 2024, 14, 536. https://doi.org/10.3390/life14040536
Tallini LR, Manfredini G, Rodríguez-Escobar ML, Ríos S, Martínez-Francés V, Feresin GE, Borges WdS, Bastida J, Viladomat F, Torras-Claveria L. The Anti-Cholinesterase Potential of Fifteen Different Species of Narcissus L. (Amaryllidaceae) Collected in Spain. Life. 2024; 14(4):536. https://doi.org/10.3390/life14040536
Chicago/Turabian StyleTallini, Luciana R., Giulia Manfredini, María Lenny Rodríguez-Escobar, Segundo Ríos, Vanessa Martínez-Francés, Gabriela E. Feresin, Warley de Souza Borges, Jaume Bastida, Francesc Viladomat, and Laura Torras-Claveria. 2024. "The Anti-Cholinesterase Potential of Fifteen Different Species of Narcissus L. (Amaryllidaceae) Collected in Spain" Life 14, no. 4: 536. https://doi.org/10.3390/life14040536
APA StyleTallini, L. R., Manfredini, G., Rodríguez-Escobar, M. L., Ríos, S., Martínez-Francés, V., Feresin, G. E., Borges, W. d. S., Bastida, J., Viladomat, F., & Torras-Claveria, L. (2024). The Anti-Cholinesterase Potential of Fifteen Different Species of Narcissus L. (Amaryllidaceae) Collected in Spain. Life, 14(4), 536. https://doi.org/10.3390/life14040536