
Marine Science Can Contribute to the Search for Extra-Terrestrial Life

 ,
, 
 , , ,
, , ,  , ,
, ,  , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bibliographical Query to Identify “Terms” of Interest
- TITLE-ABS-KEY (astrobiol* OR astro-biol* OR “astro biol*” OR “exo-life” OR “exo life” OR “extraterr* life*”) AND TITLE-ABS-KEY (ocean* OR water OR marine OR “sub ice” OR “sub-ice” OR world* OR moon* OR “water-rock” OR “water rock” OR “water-sediment” OR “water sediment” OR “water world*”OR “Ocean* World*” OR “ocean* moon*” OR “water moon*” OR “water sediment” OR subsurface OR sub-surface OR “sub surface” OR “deep biosphere” OR “deep life” OR “hydrothermal vent*” OR “extreme environment” OR “marine extreme environment*” OR “vent*” OR “seep*”).
2.2. Analysis of Term Co-Occurrence with VOSviewer
2.3. Co-Citation Analysis Based on Keywords to Generate Article Clusters, Using CiteSpace
3. Results
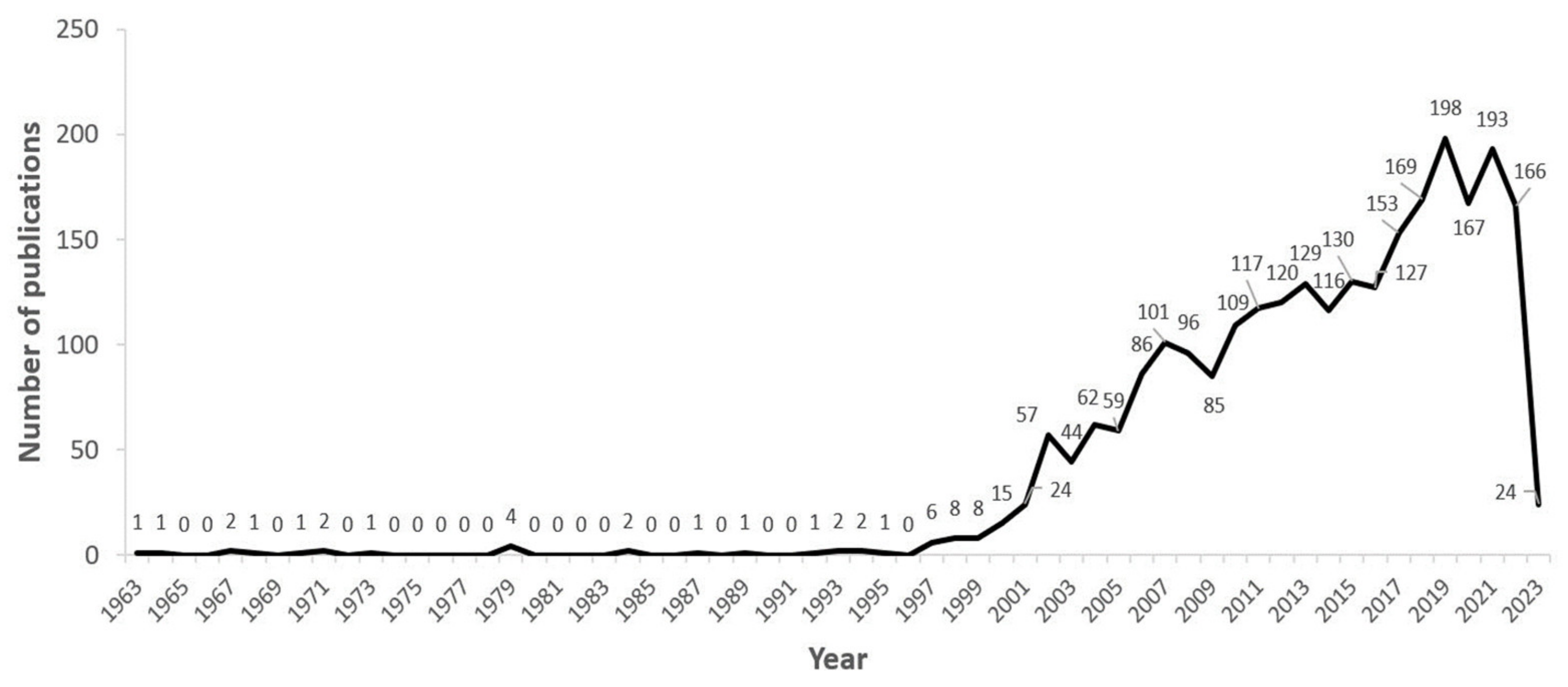
3.1. Emerging Research Trends from Term Co-Occurrence by VOSviewer Analysis
3.2. Emerging Research Trends form Article Co-Citation Analysis by CiteSpace
4. Discussion
4.1. The Search for Life on Mars
4.2. The Search for Life on Icy Moons and Their Exo-Oceans
4.3. Astronomical and Biological Parameters for Habitability of Planets, Including the Evolution of Life on Earth and Exo-Planets
4.4. Synthesis of the Three Emerging Research Areas: Life on Earth as Paradigm Constraining Astrobiology and Ocean Research
5. Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- German, C.R.; Reeves, E.P.; Türke, A.; Diehl, A.; Albers, E.; Bach, W.; Purser, A.; Ramalho, S.P.; Suman, S.; Mertens, C.; et al. Volcanically hosted venting with indications of ultramafic influence at Aurora hydrothermal field on Gakkel Ridge. Nat. Commun. 2022, 13, 6517. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Rogez, J.C.; Kalousová, K. Ocean Worlds in our Solar System. Elements 2022, 18, 161–166. [Google Scholar] [CrossRef]
- Carr, M.H.; Belton, M.J.S.; Chapman, C.R.; Davies, M.E.; Geissler, P.; Greenberg, R.; McEwen, A.S.; Tufts, B.R.; Greeley, R.; Sullivan, R.; et al. Evidence for a subsurface ocean on Europa. Nature 1998, 391, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Khurana, K.K.; Kivelson, M.G.; Stevenson, D.J.; Schubert, G.; Russell, C.T.; Walker, R.J.; Polanskey, C. Induced magnetic fields as evidence for subsurface oceans in Europa and Callisto. Nature 1998, 395, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Kivelson, M.G.; Khurana, K.K.; Russell, C.T.; Volwerk, M.; Walker, R.J.; Zimmer, C. Galileo magnetometer measurements: A stronger case for a subsurface ocean at Europa. Science 2000, 289, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Zolotov, M.Y.; Kargel, J.S. On the Chemical Composition of Europa’s Icy Shell, Ocean, and Underlying Rocks; University of Arizona Press: Tucson, AZ, USA, 2009; pp. 431–458. [Google Scholar]
- Zolotov, M.Y. An oceanic composition on early and today’s Enceladus. Geophys. Res. Lett. 2007, 34, L23203. [Google Scholar] [CrossRef]
- Postberg, F.; Tobie, G.; Dambeck, T. Under the sea of Enceladus. Sci. Am. 2016, 315, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Glein, C.R.; Baross, J.A.; Waite, J.H. The pH of Enceladus’ ocean. Geochim. Cosmochim. Acta 2015, 162, 202–219. [Google Scholar] [CrossRef]
- Glein, C.R.; Shock, E.L. Sodium chloride as a geophysical probe of a subsurface ocean on Enceladus. Geophys. Res. Lett. 2010, 37, L09204. [Google Scholar] [CrossRef]
- Cadek, O.; Tobie, G.; Van Hoolst, T.; Massé, M.; Choblet, G.; Lefèvre, A.; Mitri, G.; Baland, R.M.; Běhounková, M.; Bourgeois, O.; et al. Enceladus’s internal ocean and ice shell constrained from Cassini gravity, shape, and libration data. Geophys. Res. Lett. 2016, 43, 5653–5660. [Google Scholar] [CrossRef]
- Hendrix, A.R.; Hurford, T.A.; Barge, L.M.; Bland, M.T.; Bowman, J.S.; Brinckerhoff, W.; Buratti, B.J.; Cable, M.L.; Castillo-Rogez, J.; Collins, G.C.; et al. The NASA roadmap to ocean worlds. Astrobiology 2019, 19, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Arnold, N.S.; Butcher, F.E.G.; Conway, S.J.; Gallagher, C.; Balme, M.R. Surface topographic impact of subglacial water beneath the south polar ice cap of Mars. Nat. Astron. 2022, 6, 1256–1262. [Google Scholar] [CrossRef]
- Lauro Sebastian, E.; Pettinelli, E.; Caprarelli, G.; Baniamerian, J.; Mattei, E.; Cosciotti, B.; Stillman, D.E.; Primm, K.M.; Soldovieri, F.; Orosei, R. Using MARSIS signal attenuation to assess the presence of South Polar Layered Deposit subglacial brines. Nat. Comm. 2022, 13, 5686. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.; Riu, L.; Poulet, F.; Bibring, J.P.; Langevin, Y.; Gondet, B. A Mars Orbital Catalog of Aqueous Alteration Signatures (MOCAAS). Icarus 2023, 389, 115164. [Google Scholar] [CrossRef]
- Donahue, T.M.; Hoffman, J.H.; Hodges, R.R.; Watson, A.J. Venus was wet—A measurement of the ratio of deuterium to hydrogen. Science 1982, 216, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Michalski, J.R.; Dobrea, E.Z.N.; Niles, P.B.; Cuadros, J. Ancient hydrothermal seafloor deposits in Eridania basin on Mars. Nat. Commun. 2017, 8, 15978. [Google Scholar] [CrossRef] [PubMed]
- Garvin, J.B.; Getty, S.A.; Arney, G.N.; Johnson, N.M.; Kohler, E.; Schwer, K.O.; Sekerak, M.; Bartels, A.; Saylor, R.S.; Elliott, V.E.; et al. Revealing the Mysteries of Venus: The DAVINCI Mission. Planet. Sci. J. 2022, 3, 117. [Google Scholar] [CrossRef]
- Bean, J.L.; Abbot, D.S.; Kempton, E.M.R. A statistical comparative planetology approach to the hunt for habitable exoplanets and life beyond the Solar System. Astrophys. J. Lett. 2017, 841, L24. [Google Scholar] [CrossRef]
- Dressing, C.D.; Charbonneau, D.; Dumusque, X.; Gettel, S.; Pepe, F.; Cameron, A.C.; Latham, D.W.; Molinari, E.; Udry, S.; Affer, L.; et al. The mass of Kepler-93b and the composition of terrestrial planets. Astrophys. J. 2015, 800, 135. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Heller, R.; Guinan, E. In search for a planet better than Earth: Top contenders for a superhabitable world. Astrobiology 2020, 20, 1394–1404. [Google Scholar] [CrossRef]
- Kane, S.R.; Li, Z.; Wolf, E.T.; Ostberg, C.; Hill, M.L. Eccentricity Driven Climate Effects in the Kepler-1649 System. Astron. J. 2020, 161, 31. [Google Scholar] [CrossRef]
- Madhusudhan, N.; Piette, A.A.; Constantinou, S. Habitability and biosignatures of Hycean worlds. Astrophys. J. 2021, 918, 1. [Google Scholar] [CrossRef]
- Ida, S. Predicting ‘Earth-like’ planets around red dwarfs. Nat. Astron. 2022, 6, 1231–1232. [Google Scholar] [CrossRef]
- Mol Lous, M.; Helled, R.; Mordasini, C. Potential long-term habitable conditions on planets with primordial H–He atmospheres. Nat. Astron. 2022, 6, 819–827. [Google Scholar] [CrossRef]
- Ranjan, S.; Wordsworth, R.; Sasselov, D.D. The surface UV environment on planets orbiting M dwarfs: Implications for prebiotic chemistry and the need for experimental follow-up. Astrophys. J. 2017, 843, 110. [Google Scholar] [CrossRef]
- Rodríguez-Mozos, J.M.; Moya, A. Internal structures and magnetic moments of rocky planets-Application to the first exoplanets discovered by TESS. Astron. Astrophys. 2022, 661, A101. [Google Scholar] [CrossRef]
- Chen, H.; Wolf, E.T.; Zhan, Z.; Horton, D.E. Habitability and spectroscopic observability of warm M-dwarf exoplanets evaluated with a 3D chemistry-climate model. Astrophys. J. 2019, 886, 16. [Google Scholar] [CrossRef]
- Cleaves, H.J.; Miller, S.L. Oceanic protection of prebiotic organic compounds from UV radiation. Proc. Natl. Acad. Sci. USA 1998, 95, 7260–7263. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Fanelli, E.; Aguzzi, J.; Billett, D.; Carugati, L.; Corinaldesi, C.; Dell’Anno, A.; Gjerde, K.; Jamieson, A.J.; Kark, S.; et al. Ecological variables for developing a global deep-ocean monitoring and conservation strategy. Nat. Ecol. Evol. 2020, 4, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Snelgrove, P.V.; Tyler, P. Challenging the paradigms of deep-sea ecology. Trends Ecol. Evol. 2014, 29, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.M.; Goordial, J.M.; Orcutt, B.N. Low energy subsurface environments as extraterrestrial analogs. Front. Microbiol. 2018, 9, 1605. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Hall, A.J.; Cairns-Smith, A.G.; Braterman, P.S. Submarine hot springs and the origin of life. Nature 1988, 336, 117. [Google Scholar] [CrossRef]
- Westall, F.; Hickman-Lewis, K.; Hinman, N.; Gautret, P.; Campbell, K.A.; Bréhéret, J.G.; Foucher Hubert, F.A.; Sorieul, S.; Dass, A.V.; Kee, T.P.; et al. A hydrothermal-sedimentary context for the origin of life. Astrobiology 2018, 18, 259–293. [Google Scholar] [CrossRef] [PubMed]
- Preiner, M.; Asche, S.; Becker, S.; Betts, H.C.; Boniface, A.; Camprubi, E.; Chandru, K.; Erastova, V.; Garg, S.G.; Khawaja, N.; et al. The future of origin of life research: Bridging decades-old divisions. Life 2020, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Cody, G.D. Geochemical connections to primitive metabolism. Elements 2005, 1, 139–143. [Google Scholar] [CrossRef]
- Marion, G.M.; Fritsen, C.H.; Eicken, H.; Payne, M.C. The search for life on Europa: Limiting environmental factors, potential habitats, and earth analogues. Astrobiology 2003, 3, 785–811. [Google Scholar] [CrossRef] [PubMed]
- Antunes, A.; Olsson-Francis, K.; McGenity, T.J. Exploring deep-sea brines as potential Terrestrial analogues of oceans in the icy moons of the outer solar system. Curr. Issues Mol. Biol. 2020, 38, 123–162. [Google Scholar] [CrossRef] [PubMed]
- Hand, K.P.; Chyba, C.F.; Priscu, J.C.; Carlson, R.W.; Nealson, K.H. Astrobiology and the potential for life on Europa. Europa 2009, 589–630. [Google Scholar]
- Hsu, H.W.; Postberg, F.; Sekine, Y.; Shibuya, T.; Kempf, S.; Horányi, M.; Juhász, A.; Altobelli, N.; Suzuki, K.; Masaki, Y.; et al. Ongoing hydrothermal activities within Enceladus. Nature 2015, 519, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Sekine, Y.; Shibuya, T.; Postberg, F.; Hsu, H.W.; Suzuki, K.; Masaki, Y.; Kuwatani, T.; Mori, M.; Hong, P.K.; Yoshizaki, M.; et al. High-temperature water-rock interactions and hydrothermal environments in the chondrite-like core of Enceladus. Nat. Commun. 2015, 6, 8604. [Google Scholar] [CrossRef] [PubMed]
- Waite, J.H.; Glein, C.R.; Perryman, R.S.; Teolis, B.D.; Magee, B.A.; Miller, G.; Grimes, J.; Perry, M.E.; Miller, K.E.; Bouquet, A.; et al. Cassini finds molecular hydrogen in the Enceladus plume: Evidence for hydrothermal processes. Science 2017, 356, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Robidel, R.; Le Mouélic, S.; Tobie, G.; Massé, M.; Seignovert, B.; Sotin, C.; Rodriguez, S. Photometrically-corrected global infrared mosaics of Enceladus: New implications for its spectral diversity and geological activity. Icarus 2020, 349, 113848. [Google Scholar] [CrossRef]
- Hao, J.; Glein, C.R.; Huang, F.; Yee, N.; Catling, D.C.; Postberg, F.; Hillier, J.K.; Hazen, R.M. Abundant phosphorus expected for possible life in Enceladus’s ocean. Proc. Natl. Acad. Sci. USA 2022, 119, e2201388119. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, M.J.; Eaton, D.A.R.; Maya-Lastra, C.A.; Landis, M.J.; Sweeney, P.W.; Olson, M.E.; Cacho, N.I.; Moeglein, M.K.; Gardner, J.R.; Heaphy, N.M.; et al. Replicated radiation off a plant clade along a cloud forest archipelago. Nat. Ecol. Evol. 2022, 6, 1318–1329. [Google Scholar] [CrossRef] [PubMed]
- Michalski, J.R.; Goudge, T.A.; Crowe, S.A.; Cuadros, J.; Mustard, J.F.; Johnson, S.S. Geological diversity and microbiological potential of lakes on Mars. Nat. Astron. 2022, 6, 1133–1141. [Google Scholar] [CrossRef]
- Stone, J.; Edgar, J.O.; Gould, J.A.; Telling, J. Tectonically-driven oxidant production in the hot biosphere. Nat. Commun. 2022, 13, 4529. [Google Scholar] [CrossRef]
- Wallner, M.; Jarraya, M.; Olsson, E.; Idebohn, V.; Squibb, R.J.; Yaghlane, S.B.; Nyman, G.; Heland, J.H.D.; Feifel, R.; Hochlaf, M. Abiotic molecular oxygen production—Ionic pathway from sulfur dioxide. Sci. Adv. 2022, 8, eabq5411. [Google Scholar] [CrossRef]
- Aguzzi, J.; Costa, C.; Calisti, M.; Funari, V.; Stefanni, S.; Danovaro, R.; Gomes, H.I.; Vecchi, F.; Dartnell, L.R.; Weiss, P.; et al. Research trends and future perspectives in marine biomimicking robotics. Sensors 2021, 21, 3778. [Google Scholar] [CrossRef] [PubMed]
- Angelis, G.; Kordopati, G.G.; Zingkou, E.; Karioti, A.; Sotiropoulou, G.; Pampalakis, G. Plausible emergence of biochemistry in Enceladus based on chemobrionics. Chem. Eur. J. 2021, 27, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Belousov, A.; Miller, M.; Continetti, R.; Madzunkov, S.; Simcic, J.; Nikolic, D.; Maiwald, F.; Waller, S.; Malaska, M.; Cable, M. Sampling accelerated micron scale ice particles with a quadrupole ion trap mass spectrometer. J. Am. Soc. Mass Spectrom. 2021, 32, 1162–1168. [Google Scholar] [CrossRef] [PubMed]
- Cable, M.L.; Porco, C.; Glein, C.R.; German, C.R.; MacKenzie, S.M.; Neveu, M.; Hoehler, T.M.; Hofmann, A.E.; Hendrix, A.R.; Eigenbrode, J.; et al. The science case for a return to Enceladus. Planet. Sci. J. 2021, 2, 132. [Google Scholar] [CrossRef]
- Choukroun, M.; Backes, P.; Cable, M.L.; Fayolle, E.C.; Hodyss, R.; Murdza, A.; Schulson, E.M.; Badescu, M.; Malaska, M.J.; Marteau, E.; et al. Sampling plume deposits on Enceladus’ surface to explore ocean materials and search for traces of life or biosignatures. Planet. Sci. J. 2021, 2, 100. [Google Scholar] [CrossRef]
- Chow, L.; Mahaffy, P.; Trainer, M.; Eigenbrode, J.; Arevalo, R.; Brinckerhoff, W.; Getty, S.; Grefenstette, N.; Da Poian, V.; Fricke, G.M.; et al. Planetary mass spectrometry for agnostic life detection in the Solar System. Front. Astron. Space Sci. 2021, 8, 755100. [Google Scholar] [CrossRef]
- Duca, Z.A.; Craft, K.L.; Cable, M.L.; Stockton, A.M. Quantitative and compositional analysis of trace amino acids in icy moon analogues using a microcapillary electrophoresis laser-induced fluorescence detection system. ACS Earth Space Chem. 2022, 6, 333–345. [Google Scholar] [CrossRef]
- Edwards, D.; Nordmann, A.J.; Garcia, O.; Ruppert, N.; Gallagher, J.; Bayandor, J. Subglacial Ocean Probe Exploration, Access, and Research (SPEAR). In Proceedings of the AIAA SCITECH 2022 Forum, San Diego, CA, USA, 3–7 January 2022. [Google Scholar] [CrossRef]
- Hand, E. Planetary science. Dawn probe to look for a habitable ocean on Ceres. Science 2015, 347, 813–814. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.M.; Neveu, M.; Davila, A.F.; Lunine, J.I.; Cable, M.L.; Phillips-Lander, C.M.; Eigenbrode, J.L.; Waite, J.H.; Craft, K.L.; Hofgartner, J.D.; et al. Science objectives for flagship-class mission concepts for the search for evidence of life at Enceladus. Astrobiology 2022, 22, 685–712. [Google Scholar] [CrossRef] [PubMed]
- New, J.S.; Kazemi, B.; Spathis, V.; Price, M.C.; Mathies, R.A.; Butterworth, A.L. Quantitative evaluation of the feasibility of sampling the ice plumes at Enceladus for biomarkers of extraterrestrial life. Proc. Natl. Acad. Sci. USA 2021, 118, e2106197118. [Google Scholar] [CrossRef]
- Aguzzi, J.; Flogel, S.; Marini, S.; Thomsen, L.; Albiez, J.; Weiss, P.; Picardi, G.; Calisti, M.; Stefanni, S.; Mirimin, L.; et al. Developing technological synergies between deep-sea and space research. Elem. Sci. Anthr. 2022, 10, 00064. [Google Scholar] [CrossRef]
- Aguzzi, J.; Flexas, M.M.; Flogel, S.; Lo Iacono, C.; Tangherlini, M.; Costa, C.; Marini, S.; Danovaro, R.; Stefanni, S.; Thomsen, L.; et al. Exo-oceans exploration with deep-sea sensor and platform technologies. Astrobiology 2020, 20, 897–915. [Google Scholar] [CrossRef]
- Robison, B.H.; Reisenbichler, K.R.; Sherlock, R.E. The coevolution of midwater research and ROV technology at MBARI. Oceanography 2017, 30, 26–37. [Google Scholar] [CrossRef]
- Liang, J.; Feng, J.C.; Zhang, S.; Cai, Y.; Yang, Z.; Ni, T.; Yang, H.Y. Role of deep-sea equipment in promoting the forefront of studies on life in extreme environments. Iscience 2021, 24, 103299. [Google Scholar] [CrossRef] [PubMed]
- Aguzzi, J.; Chatzievangelou, D.; Marini, S.; Fanelli, E.; Danovaro, R.; Flogel, S.; Lebris, N.; Juanes, F.; De Leo, F.C.; Del Rio, J.; et al. New high-tech interactive and flexible networks for the future monitoring of deep-sea ecosystems. Environ. Sci. Technol. 2019, 53, 6616–6631. [Google Scholar] [CrossRef] [PubMed]
- Stefanni, S.; Mirimin, L.; Stankovic, D.; Chatzievangelou, D.; Bongiorni, L.; Marini, S.; Modica, M.V.; Manea, E.; Bonofiglio, F.; Del Rio Fernandez, J.; et al. Framing cutting-edge integrative taxonomy in deep-sea biodiversity monitoring via eDNA and optoacoustic augmented observatories. Front. Mar. Sci. 2022, 8, 1914. [Google Scholar] [CrossRef]
- Waldmann, C.; de Vera, J.P.; Dachwald, B.; Strasdeit, H.; Sohl, F.; Hanff, H.; Kowalski, J.; Heinen, D.; Macht, S.; Bestmann, U.; et al. Search for life in ice-covered oceans and lakes beyond Earth. In Proceedings of the 2018 IEEE/OES Autonomous Underwater Vehicle Workshop (AUV), Porto, Portugal, 6–9 November 2018; pp. 1–7. [Google Scholar]
- Wirtz, M.; Hildebrandt, M. IceShuttle Teredo: An ice-penetrating robotic system to transport an exploration AUV into the ocean of Jupiter’s moon Europa. In Proceedings of the 67th International Astronautical Congress (IAC), Guadalajara, Mexico, 26–30 September 2016; pp. 26–30. [Google Scholar]
- Van Eck, N.; Waltman, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2010, 84, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Donthu, N.; Kumar, S.; Mukherjee, D.; Pandey, N.; Lim, W.M. How to conduct a bibliometric analysis: An overview and guidelines. J. Bus. Res. 2021, 133, 285–296. [Google Scholar] [CrossRef]
- Aydinoglu, A.U.; Taşkın, Z. Origins of life research: A bibliometric approach. Orig. Life Evol. Biosph. 2018, 48, 55–71. [Google Scholar] [CrossRef]
- Malaterre, C.; Lareau, F. The Emergence of Astrobiology: A Topic-Modeling Perspective. Astrobiology 2023, 23, 496–512. [Google Scholar] [CrossRef] [PubMed]
- Tonietti, L.; Barosa, B.; Pioltelli, E.; Giovannelli, D.; Covone, G.; Di Donato, P.; Cordone, A.; Inno, L.; Magliano, C.; Fiscale, S.; et al. Exploring the Development of Astrobiology Scientific Research through Bibliometric Network Analysis: A Focus on Biomining and Bioleaching. Minerals 2023, 13, 797. [Google Scholar] [CrossRef]
- Costa, C.; Fanelli, E.; Marini, S.; Danovaro, R.; Aguzzi, J. Global deep-sea biodiversity research trends highlighted by science mapping approach. Front. Mar. Sci. 2020, 7, 384. [Google Scholar] [CrossRef]
- Ioannoni, V.; Vitale, T.; Costa, C.; Elliott, I. Depicting communities of Romani studies: On the who, when and where of Roma related scientific publications. Scientometrics 2020, 122, 1473–1490. [Google Scholar] [CrossRef]
- Manfredini, A.; Malusà, E.; Costa, C.; Pallottino, F.; Mocali, S.; Pinzari, F.; Canfora, L. Current methods, common practices, and perspectives in tracking and monitoring bioinoculants in soil. Front. Microbiol. 2021, 12, 698491. [Google Scholar] [CrossRef] [PubMed]
- Waltman, L.; Van Eck, N.J. A smart local moving algorithm for large-scale modularity-based community detection. Eur. Phys. J. B 2013, 86, 471. [Google Scholar] [CrossRef]
- Waltman, L.; Van Eck, N.J.; Noyons, E.C. A unified approach to mapping and clustering of bibliometric networks. J. Informetr. 2010, 4, 629–635. [Google Scholar] [CrossRef]
- Chen, C. CiteSpace: A Practical Guide for Mapping Scientific Literature; Nova Science Publishers: Hauppauge, NY, USA, 2016; pp. 41–44. [Google Scholar]
- Chen, C. The CiteSpace Manual. 2014. Available online: http://cluster.ischool.drexel.edu/~cchen/citespace/CiteSpaceManual.pdf (accessed on 17 November 2023).
- Su, X.; Li, X.; Kang, Y. A bibliometric analysis of research on intangible cultural heritage using CiteSpace. Sage Open 2019, 9, 2158244019840119. [Google Scholar] [CrossRef]
- Dastani, M.; Chelak, A.M.; Ziaei, S.; Delghandi, F. Identifying emerging trends in scientific texts using TF-IDF algorithm: A case study of medical librarianship and information articles. Health Technol. Assess. Action 2020, 4, e6231. [Google Scholar] [CrossRef]
- Miner, G. Practical Text Mining and Statistical Analysis for Non-Structured Text Data Applications; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Porter, M.F. An algorithm for suffix stripping. Program 1980, 14, 130–137. [Google Scholar] [CrossRef]
- Kleinberg, J. Bursty and hierarchical structure in streams. In Proceedings of the Eighth ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, Edmonton, AB, Canada, 23–26 July 2002; pp. 91–101. [Google Scholar] [CrossRef]
- Hecht, M.H.; Catling, D.C.; Clark, B.C.; De Flores, L.; Gospodinova, K.; Kapit, J.; Kounaves, S.P.; Ming, D.W.; Quinn, R.C.; West, S.J.; et al. Perchlorate in Martian soil: Evidence and implications. In Proceedings of the 40th Annual Lunar and Planetary Science Conference, The Woodlands, TX, USA, 23–27 March 2009; p. 2420. [Google Scholar]
- Mumma, M.J.; Villanueva, G.L.; Novak, R.E.; Hewagama, T.; Bonev, B.P.; Disanti, M.A.; Mandell, A.M.; Smith, M.D. Strong release of methane on Mars in northern summer 2003. Science 2009, 323, 1041–1045. [Google Scholar] [CrossRef]
- Grotzinger, J.P.; Sumner, D.Y.; Kah, L.C.; Stack, K.; Gupta, S.; Edgar, L.; Rubin, D.; Lewis, K.; Schieber, J.; Mangold, N.; et al. A habitable fluvio-Lacustrine environment at Yellowknife Bay, Gale Crater, Mars. Science 2014, 343, 1242777. [Google Scholar] [CrossRef] [PubMed]
- Eigenbrode, J.L.; Summons, R.E.; Steele, A.; Freissinet, C.; Millan, M.; Navarro-Gonzàlez, R.; Sutter, B.; McAdam, A.C.; Franz, H.B.; Glavin, D.P.; et al. Organic matter preserved in 3-billion-year-old mudstones at Gale crater, Mars. Science 2018, 360, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Gillon, M.; Triaud, A.H.M.J.; Demory, B.O.; Jehin, E.; Agol, E.; Deck, K.M.; Lederer, S.M.; de Wit, J.; Burdanov, A.; Ingalls, J.G.; et al. Seven temperate terrestrial planets around the nearby ultracool dwarf star TRAPPIST-1. Nature 2017, 542, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Kopparapu, R.K.; Ramirez, R.; Kasting, J.F.; Eymet, V.; Robinson, T.D.; Mahadevan, S.; Terrien, R.C.; Domagal-Goldman, S.; Meadows, V.; Deshpande, R. Habitable zones around main-sequence stars: New estimates. Astrophys. J. 2013, 765, 131. [Google Scholar] [CrossRef]
- Allwood, A.C.; Walter, M.R.; Kamber, B.S.; Marshall, C.P.; Burch, I.W. Stromatolite reef from the Early Archaean era of Australia. Nature 2006, 441, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Poulet, F.; Bibring, J.P.; Mustard, J.F.; Gendrin, A.; Mangold, N.; Langevin, Y.; Arvidson, R.E.; Gondet, B.; Gomez, C. Phyllosilicates on Mars and implications for early Martian climate. Nature 2005, 438, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Barnes, R.; Heller, R. Habitable planets around white and brown dwarfs: The perils of a cooling primary. Astrobiology 2013, 13, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.; Loizeau, D.; Foucher, F.; Bost, N.; Betrand, M.; Vago, J.; Kminek, G. Habitability on Mars from a microbial point of view. Astrobiology 2013, 13, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Ojha, L.; Wilhelm, M.B.; Murchie, S.L.; McEwen, A.S.; Wray, J.J.; Hanley, J.; Massé, M.; Chojnacki, M. Spectral evidence for hydrated salts in recurring slope lineae on Mars. Nat. Geosci. 2015, 8, 829–832. [Google Scholar] [CrossRef]
- Westall, F.; Foucher, F.; Bost, N.; Bertrand, M.; Loizeau, D.; Vago, J.L.; Kminek, G.; Gaboyer, F.; Campbell, K.A.; Bréhéret, J.G.; et al. Biosignatures on Mars: What, where, and how? Implications for the search for martian life. Astrobiology 2015, 15, 998–1029. [Google Scholar] [CrossRef] [PubMed]
- Lammer, H.; Bredehöft, J.H.; Coustenis, A.; Khodachenko, M.L.; Kaltenegger, L.; Grasset, O.; Prieur, D.; Raulin, F.; Ehrenfreund, P.; Yamauchi, M.; et al. What makes a planet habitable? Astron. Astrophys. Rev. 2009, 17, 181–249. [Google Scholar] [CrossRef]
- McEwen, A.S.; Ojha, L.; Dundas, C.M.; Mattson, S.S.; Byrne, S.; Wray, J.J.; Cull, S.C.; Murchie, S.L.; Thomas, N.; Gulick, V. Seasonal flows on warm Martian slopes. Science 2011, 333, 740–743. [Google Scholar] [CrossRef] [PubMed]
- Vago, J.L.; Westall, F.; Coates, A.J.; Jaumann, R.; Korablev, O.; Ciarletti, V.; Mitrofanov, I.; Josset, J.L.; De Sanctis, M.C.; Bibring, J.P.; et al. Habitability on early Mars and the search for biosignatures with the ExoMars Rover. Astrobiology 2017, 17, 471–510. [Google Scholar] [CrossRef] [PubMed]
- Hays, L.E.; Graham, H.V.; Des Marais, D.J.; Hausrath, E.M.; Horgan, B.; McCollom, T.M.; Parentau, M.N.; Potter-McIntyre, S.L.; Williams, A.J.; Lynch, K.L. Biosignature preservation and detection in Mars analog environments. Astrobiology 2017, 17, 363–400. [Google Scholar] [CrossRef] [PubMed]
- Goesmann, F.; Brinckerhoff, W.B.; Raulin, F.; Goetz, W.; Danell, R.M.; Getty, S.A.; Siljeström, S.; Mißbach, H.; Steininger, H.; Arevalo, R.D.; et al. The Mars Organic Molecule Analyzer (MOMA) instrument: Characterization of organic material in Martian sediments. Astrobiology 2017, 17, 655–685. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, R.M.; Kaltenegger, L. A methane extension to the classical habitable zone. Astrophys. J. 2018, 858, 72. [Google Scholar] [CrossRef]
- Luger, R.; Barnes, R. Extreme water loss and abiotic O2 buildup on planets throughout the habitable zones of M dwarfs. Astrobiology 2015, 15, 119–143. [Google Scholar] [CrossRef] [PubMed]
- Greaves, J.S.; Richards, A.M.S.; Bains, W.; Rimmer, P.B.; Sagawa, H.; Clements, D.L.; Seager, S.; Petkowski, J.J.; Sousa-Silva, C.; Ranjan, S.; et al. Phosphine gas in the cloud decks of Venus. Nat. Astron. 2020, 5, 655–664. [Google Scholar] [CrossRef]
- Bashir, A.K.; Wink, L.; Duller, S.; Schwendner, P.; Cockell, C.; Rettberg, P.; Mahnert, A.; Beblo-Vranesevic, K.; Bohmeier, M.; Rabbow, E.; et al. Taxonomic and functional analyses of intact microbial communities thriving in extreme, astrobiology-relevant, anoxic sites. Microbiome 2021, 9, 50. [Google Scholar] [CrossRef] [PubMed]
- Salter, T.L.; Watson, J.S.; Waite, J.H.; Sephton, M.A. Hydrothermal processing of microorganisms: Mass spectral signals of degraded biosignatures for life detection on icy moons. ACS Earth Space Chem. 2022, 6, 2508–2518. [Google Scholar] [CrossRef] [PubMed]
- Jebbar, M.; Hickman-Lewis, K.; Cavalazzi, B.; Taubner, R.S.; Rittmann, S.K.M.; Antunes, A. Microbial Diversity and Biosignatures: An Icy Moons Perspective. Space Sci. Rev. 2020, 216, 10. [Google Scholar] [CrossRef]
- Kotsyurbenko, O.R.; Cordova, J.A.; Belov, A.A.; Cheptsov, V.S.; Kolbl, D.; Khrunyk, Y.K.; Kryuchkova, M.O.; Milojevic, T.; Mogul, R.; Sasaki, S.; et al. Exobiology of the Venusian clouds: New insights into habitability through terrestrial models and methods of detection. Astrobiology 2021, 21, 1186–1205. [Google Scholar] [CrossRef]
- Limaye, S.S.; Mogul, R.; Baines, K.H.; Bullock, M.A.; Cockell, C.; Cutts, J.A.; Gentry, D.M.; Grinspoon, D.H.; Head, J.W.; Jessup, K.L.; et al. Venus, an Astrobiology Target. Astrobiology 2021, 21, 1163–1185. [Google Scholar] [CrossRef] [PubMed]
- Pontefract, A.; Hachey, J.; Mojarro, A.; Walker, V.K.; Rowedder, H.; Zhu, T.F.; Lui, C.; Zuber, M.T.; Ruvkun, G.; Carr, C.E. Understanding habitability and biosignature preservation in a hypersaline Mars analog environment: Lessons from Spotted Lake. In Proceedings of the 48th Annual Lunar and Planetary Science Conference, The Woodlands, TX, USA, 20–24 March 2017; p. 1124. [Google Scholar]
- Wang, J.L.; Dragone, N.B.; Avard, G.; Hynek, B.M. Microbial Survival in an Extreme Martian Analog Ecosystem: Poás Volcano, Costa Rica. Front. Astron. Space Sci. 2022, 9, 817900. [Google Scholar] [CrossRef]
- Finkel, P.L.; Carrizo, D.; Parro, V.; Sánchez-García, L. An overview of lipid biomarkers in terrestrial extreme environments with relevance for Mars Exploration. Astrobiology 2023, 23, 563–604. [Google Scholar] [CrossRef] [PubMed]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the extremes: Extremophiles and the limits of life in a planetary context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef]
- Cheptsov, V.S.; Belov, A.A.; Vorobyova, E.A.; Pavlov, A.K.; Lomasov, V.N. Effects of radiation intensity, mineral matrix, and pre-irradiation on the bacterial resistance to gamma irradiation under low temperature conditions. Microorganisms 2021, 9, 198. [Google Scholar] [CrossRef] [PubMed]
- Gevi, F.; Leo, P.; Leo, P.; Cassaro, A.; Pacelli, C.; de Vera, J.P.P.; Rabbow, E.; Timperio, A.M.; Onofri, S. Metabolomic profile of the fungus Cryomyces antarcticus under simulated Martian and space conditions as support for life-detection missions on Mars. Front. Microbiol. 2022, 13, 749396. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, A.; Cheptsov, V.; Tsurkov, D.; Lomasov, V.; Frolov, D.; Vasiliev, G. Survival of radioresistant bacteria on Europa’s surface after pulse ejection of subsurface ocean water. Geosciences 2019, 9, 9. [Google Scholar] [CrossRef]
- Sadlok, G. On a hypothetical mechanism of interstellar life transfer through nomadic objects. Orig. Life Evol. Biosph. 2020, 50, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Rowe, L.; Peller, J.; Mammoser, C.; Davidson, K.; Gunter, A.; Brown, B.; Dhar, S. Stability of non-proteinogenic amino acids to UV and gamma irradiation. Int. J. Astrobiol. 2019, 18, 426–435. [Google Scholar] [CrossRef]
- Chela-Flores, J. Testing S isotopes as biomarkers for Mars. Int. J. Astrobiol. 2019, 18, 436–439. [Google Scholar] [CrossRef]
- Cieslarova, Z.; Noell, A.C.; Willis, P.A.; Mora, M.F. From Microorganisms to biosignatures: Subcritical water extraction as a sample preparation technique for future life detection missions. Geophys. Res. Lett. 2022, 49, e2022GL098082. [Google Scholar] [CrossRef]
- Enya, K.; Yoshimura, Y.; Kobayashi, K.; Yamagishi, A. Extraterrestrial life signature detection microscopy: Search and analysis of cells and organics on mars and other Solar System bodies. Space Sci. Rev. 2022, 218, 49. [Google Scholar] [CrossRef]
- Lingam, M. Theoretical constraints imposed by gradient detection and dispersal on microbial size in astrobiological environments. Astrobiology 2021, 21, 813–830. [Google Scholar] [CrossRef] [PubMed]
- Materese, C.K.; Gerakines, P.A.; Hudson, R.L. The Radiation Stability of Thymine in Solid H2O. Astrobiology 2020, 20, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Moelling, K.; Broecker, F. Viroids-first: A model for life on earth, Mars and exoplanets. Geosciences 2019, 9, 241. [Google Scholar] [CrossRef]
- Des Marais, D.J. Exploring Mars for evidence of habitable environments and life. Proc. Am. Philos. Soc. 2010, 154, 402–421. [Google Scholar]
- Michalski, J.R.; Cuadros, J.; Niles, P.B.; Parnell, J.; Deanne Rogers, A.; Wright, S.P. Groundwater activity on Mars and implications for a deep biosphere. Nat. Geosci. 2013, 6, 133–138. [Google Scholar] [CrossRef]
- Rucker, H.R.; Ely, T.D.; LaRowe, D.E.; Giovannelli, D.; Price, R.E. Quantifying the Bioavailable Energy in an Ancient Hydrothermal Vent on Mars and a Modern Earth-Based Analog. Astrobiology 2023, 23, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Azua-Bustos, A.; Gonzalez-Silva, C.; Fairen, A.G. The Atacama Desert in Northern Chile as an Analog Model of Mars. Front. Astron. Space Sci. 2022, 8, 810426. [Google Scholar] [CrossRef]
- Carré, L.; Zaccai, G.; Delfosse, X.; Girard, E.; Franzetti, B. Relevance of Earth-Bound Extremophiles in the Search for Extraterrestrial Life. Astrobiology 2022, 22, 322–367. [Google Scholar] [CrossRef] [PubMed]
- Coleine, C.; Delgado-Baquerizo, M. Unearthing terrestrial extreme microbiomes for searching terrestrial-like life in the Solar System. Trends Microbiol. 2022, 30, 1101–1115. [Google Scholar] [CrossRef] [PubMed]
- Canfora, L.; Bacci, G.; Pinzari, F.; Lo Papa, G.; Dazzi, C.; Benedetti, A. Salinity and bacterial diversity: To what extent does the concentration of salt affect the bacterial community in a saline soil? PLoS ONE 2014, 9, e106662. [Google Scholar] [CrossRef] [PubMed]
- Beblo-Vranesevic, K.; Piepjohn, J.; Antunes, A.; Rettberg, P. Surviving Mars: New insights into the persistence of facultative anaerobic microbes from analogue sites. Int. J. Astrobiol. 2022, 21, 110–127. [Google Scholar] [CrossRef]
- Naz, N.; Liu, D.; Harandi, B.F.; Kounaves, S.P. Microbial growth in Martian soil simulants under Terrestrial conditions: Guiding the search for life on Mars. Astrobiology 2022, 22, 1210–1221. [Google Scholar] [CrossRef] [PubMed]
- Maus, D.; Heinz, J.; Schirmack, J.; Airo, A.; Kounaves, S.P.; Wagner, D.; Schulze-Makuch, D. Methanogenic Archaea can produce methane in deliquescence-driven Mars analog environments. Sci. Rep. 2020, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, E.; Altshuler, I.; Fernández-Martínez, M.Á.; Chen, Y.J.; Maggiori, C.; Goordial, J.; Whyte, L.G. Active lithoautotrophic and methane-oxidizing microbial community in an anoxic, sub-zero, and hypersaline High Arctic spring. ISME J. 2022, 16, 1798–1808. [Google Scholar] [CrossRef] [PubMed]
- Goordial, J. Cryomicrobial ecology: Still much to learn about life left out in the cold. mSystems 2021, 6, e00852-21. [Google Scholar] [CrossRef] [PubMed]
- Gómez, F.; Walter, N.; Amils, R.; Rull, F.; Klingelhöfer, A.K.; Kviderova, J.; Sarrazin, P.; Foing, B.; Behar, A.; Fleischer, I.; et al. Multidisciplinary integrated field campaign to an acidic Martian Earth analogue with astrobiological interest: Rio Tinto. Int. J. Astrobiol. 2011, 10, 291–305. [Google Scholar] [CrossRef]
- Ramos-Tapia, I.; Nuñez, R.; Salinas, C.; Salinas, P.; Soto, J.; Paneque, M. Study of wetland soils of the Salar de Atacama with different zonal vegetative formations reveals changes in the microbiota associated with hygrophile plant type on the soil surface. Microbiol. Spectr. 2022, 10, e00533-22. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, H.; Hur, S.D.; Sul, W.J.; Kim, O.S. Glaciers as microbial habitats: Current knowledge and implication. J. Microbiol. 2022, 60, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Vachier, J.; Wettlaufer, J.S. Biolocomotion and premelting in Ice. Front. Phys. 2022, 10, 904836. [Google Scholar] [CrossRef]
- Von Hegner, I. Extremophiles: A special or general case in the search for extraterrestrial life? Extremophiles 2020, 24, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Von Hegner, I. A limbus mundi elucidation of habitability: The Goldilocks edge. Int. J. Astrobiol. 2020, 19, 320–329. [Google Scholar] [CrossRef]
- Dickson, J.; Martinez, E.; Pagano, J.J.; Hudson, R.; Perl, S.M.; Barge, L.M. Incorporating Microbes into Laboratory-Grown Chimneys for Hydrothermal Microbiology Experiments. ACS Earth Space Chem. 2022, 6, 953–961. [Google Scholar] [CrossRef]
- Hallsworth, J.E.; Mancinelli, R.L.; Conley, C.A.; Dallas, T.D.; Rinaldi, T.; Davila, A.F.; Benison, K.C.; Rapoport, A.; Cavalazzi, B.; Selbmann, L.; et al. Astrobiology of life on Earth. Environ. Microbiol. 2021, 23, 3335–3344. [Google Scholar] [CrossRef] [PubMed]
- Longo, A.; Damer, B. Factoring origin of life hypotheses into the search for life in the Solar system and beyond. Life 2020, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- McClain, C.R.; Bryant, S.R.; Hanks, G.; Bowles, M.W. Extremophiles in Earth’s Deep seas: A view toward life in exo-oceans. Astrobiology 2022, 22, 1009–1028. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Barge, L.M.; Bhartia, R.; Bocanegra, D.; Bracher, P.J.; Branscomb, E.; Kidd, R.; McGlynn, S.; Meier, D.H.; Nitschke, W.; et al. The drive to life on wet and Icy Worlds. Astrobiology 2014, 14, 308–343. [Google Scholar] [CrossRef] [PubMed]
- Soderlund, K.M.; Kalousová, K.; Buffo, J.J.; Glein, C.R.; Goodman, J.C.; Mitri, G.; Patterson, G.W.; Postberg, F.; Rovira-Navarro, M.; Rückriemen, T.; et al. Ice-ocean exchange processes in the Jovian and Saturnian satellites. Space Sci. Rev. 2020, 216, 80. [Google Scholar] [CrossRef]
- Taubner, R.S.; Pappenreiter, P.; Zwicker, J.; Smrzka, D.; Pruckner, C.; Kolar, P.; Bernacchi, S.; Seifert, A.H.; Krajete, A.; Bach, W.; et al. Simulating putative Enceladus-like conditions: The possibility of biological methane production on Saturn’s icy moon. Proc. Int. Astron. Union. 2019, 14, 219–221. [Google Scholar] [CrossRef]
- Bywaters, K.; Stoker, C.R.; Batista Do Nascimento, N.; Lemke, L. Towards determining biosignature retention in Icy world plumes. Life 2020, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Salter, T.L.; Magee, B.A.; Waite, J.H.; Sephton, M.A. Spectrometric Fingerprints of Bacteria and Archaea for Life Detection on Icy Moons. Astrobiology 2022, 22, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.B.; Drew, A.; Malloy, J.F.; Walker, S.I. Seeding biochemistry on other worlds: Enceladus as a case study. Astrobiology 2021, 21, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Coelho, L.F.; Madden, J.; Kaltenegger, L.; Zinder, S.; Philpot, W.; Esquìvel, M.G.; Canàrio, J.; Costa, R.; Vincent, W.F.; Martins, Z. Color catalogue of life in ice: Surface biosignatures on icy worlds. Astrobiology 2022, 22, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Kanik, I.; Paul de Vera, J.P. Astrobiology of Mars, Europa, Titan and Enceladus-Most Likely Places for Alien Life. Front. Astron. Space Sci. 2021, 8, 643268. [Google Scholar] [CrossRef]
- McKay, C.P. An approach to searching for life on Mars, Europa, and Enceladus. Strateg. Life Detect. 2008, 135, 49–54. [Google Scholar] [CrossRef]
- Moreras-Marti, A.; Fox-Powell, M.; Cousins, C.R.; Macey, M.C.; Zerkle, A.L. Sulphur isotopes as biosignatures for Mars and Europa exploration. J. Geol. Soc. Lond. 2022, 179, jgs2021-134. [Google Scholar] [CrossRef]
- McKay, C.P.; Andersen, D.; Davila, A. Antarctic environments as models of planetary habitats: University Valley as a model for modern Mars and Lake Untersee as a model for Enceladus and ancient Mars. Polar J. 2017, 7, 303–318. [Google Scholar] [CrossRef]
- Zacny, K.; Paulsen, G.; Bar-Cohen, Y.; Beegle, L.; Sherrit, S.; Badescu, M.; Mellerowicz, B.; Craft, J.; Sadick, S.; Corsetti, F.; et al. Wireline deep drill for exploration of Mars, Europa, and Enceladus. In Proceedings of the 2013 IEEE Aerospace Conference, Big Sky, MT, USA, 2–9 March 2013; pp. 1–14. [Google Scholar]
- Kiang, N.Y.; Domagal-Goldman, S.; Parenteau, M.N.; Catling, D.C.; Fujii, Y.; Meadows, V.S.; Schwieterman, E.W.; Walker, S.I. Exoplanet biosignatures: At the dawn of a new era of planetary observations. Astrobiology 2018, 18, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Makuch, D.; Méndez, A.; Fairén, A.G.; von Paris, P.; Turse, C.; Boyer, G.; Davila, A.F.; de Sousa António, M.R.; Catling, D.; Irwin, L.N. A two-tiered approach to assess the habitability of exoplanets. Astrobiology 2011, 11, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Carrizo, D.; de Dios-Cubillas, A.; Sánchez-García, L.; López, I.; Prieto-Ballesteros, O. Interpreting molecular and isotopic biosignatures in methane-derived authigenic carbonates in the light of a potential carbon cycle in the icy moons. Astrobiology 2022, 22, 552–567. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.A.; Machado, P.; Ribeiro, J. From Atmospheric Evolution to the Search of Species of Astrobiological Interest in the Solar System—Case Studies Using the Planetary Spectrum Generator. Atmosphere 2022, 13, 461. [Google Scholar] [CrossRef]
- Gebauer, S.; Grenfell, J.L.; Lehmann, R.; Rauer, H. Evolution of earth-like planetary atmospheres around M Dwarf Stars: Assessing the atmospheres and biospheres with a coupled atmosphere biogeochemical model. Astrobiology 2018, 18, 856–872. [Google Scholar] [CrossRef] [PubMed]
- Keles, E.; Grenfell, J.L.; Godolt, M.; Stracke, B.; Rauer, H. The effect of varying atmospheric pressure upon habitability and biosignatures of earth-like planets. Astrobiology 2018, 18, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Pasek, M.A.; Mousis, O.; Lunine, J.I. Phosphorus chemistry on Titan. Icarus 2011, 212, 751–761. [Google Scholar] [CrossRef]
- Schulze-Makuch, D. The case (or not) for life in the Venusian clouds. Life 2021, 11, 255. [Google Scholar] [CrossRef] [PubMed]
- Taubner, R.S.; Pappenreiter, P.; Zwicker, J.; Smrzka, D.; Pruckner, C.; Kolar, P.; Bernacchi, S.; Seifert, A.H.; Krajete, A.; Bach, W.; et al. Biological methane production under putative Enceladus-like conditions. Nat. Commun. 2018, 9, 748. [Google Scholar] [CrossRef] [PubMed]
- Yung, Y.L.; Chen, P.; Nealson, K.; Atreya, S.; Beckett, P.; Blank, J.G.; Ehlmann, B.; Eiler, J.; Etiope, G.; Ferry, J.G.; et al. Methane on Mars and habitability: Challenges and responses. Astrobiology 2018, 18, 1221–1242. [Google Scholar] [CrossRef] [PubMed]
- Chopra, A.; Lineweaver, C.H. The case for a Gaian bottleneck: The biology of habitability. Astrobiology 2016, 16, 7–22. [Google Scholar] [CrossRef] [PubMed]
- McMahon, S.; O’Malley-James, J.; Parnell, J. Circumstellar Habitable Zones for Deep Terrestrial Biospheres. Planet. Space Sci. 2013, 85, 312–318. [Google Scholar] [CrossRef]
- Giovannelli, D. Subsurface life can modify volatile cycling on a planetary scale. Mem. Della Soc. Astron. Ital. 2021, 92, 60. [Google Scholar]
- Barry, P.H.; De Moor, J.M.; Giovannelli, D.; Schrenk, M.; Hummer, D.R.; Lopez, T.; Pratt, C.A.; Alpìsaz Segura, Y.; Battaglia, A.; Beaudry, P.; et al. Forearc carbon sink reduces long-term volatile recycling into the mantle. Nature 2019, 568, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, K.M.; Schrenk, M.O.; Yücel, M.; Manini, E.; Basili, M.; Rogers, T.J.; Fattorini, D.; Di Carlo, M.; d’Errico, G.; Regoli, F.; et al. Effect of tectonic processes on biosphere–geosphere feedbacks across a convergent margin. Nat. Geosci. 2021, 14, 301–306. [Google Scholar] [CrossRef]
- Rogers, T.J.; Buongiorno, J.; Jessen, G.L.; Schrenk, M.O.; Fordyce, J.A.; de Moor, J.M.; Ramírez, C.J.; Barry, P.H.; Yücel, M.; Selci, M.; et al. Chemolithoautotroph distributions across the subsurface of a convergent margin. ISME J. 2023, 17, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Upin, H.E.; Newell, D.L.; Colman, D.R.; Boyd, E.S. Tectonic settings influence the geochemical and microbial diversity of Peru hot springs. Commun. Earth Environ. 2023, 4, 112. [Google Scholar] [CrossRef] [PubMed]
- Chyba, C.F.; Hand, K.P. The study of the living universe. Annu. Rev. Astron. Astrophys. 2005, 43, 31–74. [Google Scholar] [CrossRef]
- Cockell, C.S.; Bush, T.; Bryce, C.; Direito, S.; Fox-Powell, M.; Harrison, J.P.; Lammer, H.; Landenmark, H.; Martin-Torres, J.; Nicholson, N.; et al. Habitability: A Review. Astrobiology 2016, 16, 89–117. [Google Scholar] [CrossRef] [PubMed]
- Sagan, C.; Thompson, W.R.; Carlson, R.; Gurnett, D.; Hord, C. A search for life on Earth from the Galileo spacecraft. Nature 1993, 365, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Herzing, D.L. Profiling nonhuman intelligence: An exercise in developing unbiased tools for describing other “types” of intelligence on earth. Acta Astronaut. 2014, 94, 676–680. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Bains, W. The Cosmic Zoo: Complex Life on Many Worlds; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Wright, J.T. Prior indigenous technological species. Int. J. Astrobiol. 2018, 17, 96–100. [Google Scholar] [CrossRef]
- Lingam, M.; Loeb, A. Dependence of Biological Activity on the Surface Water Fraction of Planets. Astron. J. 2019, 157, 25. [Google Scholar] [CrossRef]
- Benford, J.A. Drake equation for alien artifacts. Astrobiology 2021, 21, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Todd, P.M.; Miller, G.F. The evolutionary psychology of extraterrestrial Intelligence: Are there universal adaptations in search, aversion, and signalling? Biol. Theory 2018, 13, 131–141. [Google Scholar] [CrossRef]
- Frank, A.; Grinspoon, D.; Walker, S. Intelligence as a planetary scale process. Int. J. Astrobiol. 2022, 21, 47–61. [Google Scholar] [CrossRef]
- Vakoch, D.A.; Harrison, A.A. (Eds.) Civilizations beyond Earth: Extraterrestrial Life and Society; Berghahn Books: Oxford, NY, USA, 2022. [Google Scholar]
- Dick, S.J. The societal impact of extraterrestrial life: The relevance of history and the social sciences. In Astrobiology, History, and Society. Advances in Astrobiology and Biogeophysics; Springer: Berlin/Heidelberg, Germany, 2013; pp. 227–257. [Google Scholar] [CrossRef]
- Impey, C.; Spitz, A.H.; Stoeger, W. (Eds.) Encountering Life in the Universe: Ethical Foundations and Social Implications of Astrobiology; University of Arizona Press: Tucson, AZ, USA, 2013; pp. 1–269. [Google Scholar]
- Traphagan, J.W. Science, Culture and the Search for Life on Other Worlds; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Dick, S.J. Space, Time, and Aliens: Collected Works on Cosmos and Culture; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–781. [Google Scholar]
- Dunér, D. Swedenborg and the Plurality of Worlds: Astrotheology in the Eighteenth Century. Zygon 2016, 51, 450–479. [Google Scholar] [CrossRef]
- Haider, S.; Ansar, A.; Naqvi, S.A.A. Shīʿī Imāmī thought on existence, life, and extraterrestrials. Theol. Sci. 2023, 21, 261–272. [Google Scholar] [CrossRef]
- Ellery, A.A. Robotic astrobiology-prospects for enhancing scientific productivity of mars rover missions. Int. J. Astrobiol. 2018, 17, 203–217. [Google Scholar] [CrossRef]
- Sherwood, B.; Lunine, J.; Sotin, C.; Cwik, T.; Naderi, F. Program options to explore ocean worlds. Acta Astronaut. 2018, 143, 285–296. [Google Scholar] [CrossRef]
- Baxter, S. Missions to the edge of the visible. JBIS 2020, 73, 126–129. [Google Scholar]
- Goldsmith, D.; Owen, T.C. The Search for Life in the Universe, 2nd ed.; Addison-Wesley: Boston, MA, USA, 1980; p. 247. [Google Scholar]
- Otlewska, A.; Migliore, M.; Dybka-Stepien, K.; Manfredini, A.; Struszczyk-Swita, K.; Napoli, R.; Białkowska, A.; Canfora, L.; Pinzari, F. When salt meddles between plant, soil, and microorganisms. Front. Plant Sci. 2020, 11, 1429. [Google Scholar] [CrossRef]
- Cockell, C.S.; Stevens, A.H.; Prescott, R. Habitability is a binary property. Nat. Astron. 2019, 3, 956–957. [Google Scholar] [CrossRef]
- Coker, J.A. Recent advances in understanding extremophiles. F1000Research 2019, 8, 1917. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, S.J.; Beckett, P. Probing complexity: Thermodynamics and computational mechanics approaches to origins studies. Interface Focus 2019, 9, 20190058. [Google Scholar] [CrossRef] [PubMed]
- Knoch, T.A. How Genomes Emerge, Function, and Evolve: Living Systems Emergence-Genotype-Phenotype-Multilism-Genome/Systems Ecology. In Nuclear, Chromosomal, and Genomic Architecture in Biology and Medicine; Springer International Publishing: Cham, Switzerland, 2022; Volume 70, pp. 103–156. [Google Scholar] [CrossRef]
- Neidhöfer, C. On the evolution of the biological framework for insight. Philosophies 2021, 6, 43. [Google Scholar] [CrossRef]
- Noor, M.A.F. Thinking outside Earth’s box—How might heredity and evolution differ on other worlds? Evol. Educ. Outreach 2022, 15, 13. [Google Scholar] [CrossRef]
- Von Hegner, I. First principles of terrestrial life: Exemplars for potential extraterrestrial biology. Theory Biosci. 2022, 141, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Perl, S.M.; Celestian, A.J.; Cockell, C.S.; Corsetti, F.A.; Barge, L.M.; Bottjer, D.; Filiberto, J.; Baxter, B.K.; Kanik, I.; Potter-McIntyre, S.; et al. A proposed geobiology-driven nomenclature for astrobiological in-situ observations and sample analyses. Astrobiology 2021, 21, 954–967. [Google Scholar] [CrossRef] [PubMed]
- De la Higuera, I.; Lázaro, E. Viruses in astrobiology. Front. Microbiol. 2022, 13, 1032918. [Google Scholar] [CrossRef] [PubMed]
- Clements, D.L. Life before Fermi—Back to the Solar System. J. Br. Interplanet. Soc. 2018, 71, 222–224. [Google Scholar]
- Mason, P.A.; Biermann, P.L. Challenges for life in the local universe. Proc. Sci. 2019, 368, 59. [Google Scholar]
- Stevenson, D.S.; Wallace, R. Biogeographical modeling of alien worlds. Astrobiology 2021, 21, 831–844. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguzzi, J.; Cuadros, J.; Dartnell, L.; Costa, C.; Violino, S.; Canfora, L.; Danovaro, R.; Robinson, N.J.; Giovannelli, D.; Flögel, S.; et al. Marine Science Can Contribute to the Search for Extra-Terrestrial Life. Life 2024, 14, 676. https://doi.org/10.3390/life14060676
Aguzzi J, Cuadros J, Dartnell L, Costa C, Violino S, Canfora L, Danovaro R, Robinson NJ, Giovannelli D, Flögel S, et al. Marine Science Can Contribute to the Search for Extra-Terrestrial Life. Life. 2024; 14(6):676. https://doi.org/10.3390/life14060676
Chicago/Turabian StyleAguzzi, Jacopo, Javier Cuadros, Lewis Dartnell, Corrado Costa, Simona Violino, Loredana Canfora, Roberto Danovaro, Nathan Jack Robinson, Donato Giovannelli, Sascha Flögel, and et al. 2024. "Marine Science Can Contribute to the Search for Extra-Terrestrial Life" Life 14, no. 6: 676. https://doi.org/10.3390/life14060676
APA StyleAguzzi, J., Cuadros, J., Dartnell, L., Costa, C., Violino, S., Canfora, L., Danovaro, R., Robinson, N. J., Giovannelli, D., Flögel, S., Stefanni, S., Chatzievangelou, D., Marini, S., Picardi, G., & Foing, B. (2024). Marine Science Can Contribute to the Search for Extra-Terrestrial Life. Life, 14(6), 676. https://doi.org/10.3390/life14060676