Abstract
Sarcopenia, the age-related decline in muscle mass and function, poses a significant health challenge as the global population ages. Mitochondrial dysfunction is a key factor in sarcopenia, as evidenced by the role of mitochondrial reactive oxygen species (mtROS) in mitochondrial biogenesis and dynamics, as well as mitophagy. Resistance exercise training (RET) is a well-established intervention for sarcopenia; however, its effects on the mitochondria in aging skeletal muscles remain unclear. This review aims to elucidate the relationship between mitochondrial dynamics and sarcopenia, with a specific focus on the implications of RET. Although aerobic exercise training (AET) has traditionally been viewed as more effective for mitochondrial enhancement, emerging evidence suggests that RET may also confer beneficial effects. Here, we highlight the potential of RET to modulate mtROS, drive mitochondrial biogenesis, optimize mitochondrial dynamics, and promote mitophagy in aging skeletal muscles. Understanding this interplay offers insights for combating sarcopenia and preserving skeletal muscle health in aging individuals.
1. Introduction
A decline in physical fitness naturally occurs as individuals age, leading to increased vulnerability to both physical and psychological impairments [1]. This is reflected in an increased prevalence of chronic health issues. A key feature of aging is the progressive loss of muscle mass, strength, and function, known as sarcopenia [2,3].
Loss of muscle mass in older adults is associated not only with falls and fractures but also with an increased risk of chronic diseases such as diabetes, hypertension, heart disease, and cancer [2,4,5]. The etiology of sarcopenia involves a range of factors and pathways, ranging from environmental influences such as physical inactivity and poor diet to cellular changes such as reduced satellite cell counts, activation of apoptotic pathways, and mitochondrial dysfunction [2]. Emerging evidence indicates that mitochondrial dysfunction and the activation of apoptotic pathways play pivotal roles in the development of age-related sarcopenia [6].
Mitochondria serve as the powerhouses of cells, responsible for producing adenosine triphosphate (ATP), and thus play a critical role in central metabolic pathways and cellular functions, ultimately impacting the overall health of the individual. Deficiencies in these processes can lead to various diseases, such as Alzheimer’s and cancer [7]. Furthermore, the mitochondria play a crucial role in regulating the metabolic function of skeletal muscles, and mitochondrial dysfunction directly affects normal skeletal muscle functioning [8]. In older adults with sarcopenia, skeletal muscle mitochondria demonstrate a characteristic decline in the expression and activity of mitochondrial respiratory complexes, accompanied by suppressed oxidative phosphorylation and downregulation of genes responsible for mitochondrial protein quality control [9].
Exercise provides numerous benefits, including improved cardiovascular health, enhanced skeletal muscle and bone health, chronic disease prevention, longevity, and improved immune function in aging individuals with sarcopenia [10]. Resistance exercise training (RET) and aerobic exercise training (AET) are two distinct exercise modalities with well-documented health benefits in aging patients with sarcopenia. Typically, the adaptation of skeletal muscles to exercise is often viewed through a dichotomous lens, where AET induces increased mitochondrial adaptations, whereas RET leads to myofibrillar adaptations through mechanical tension [11]. AET improves endurance, oxidative capacity, mitochondrial content, and function [12,13]. A systematic review and meta-analysis of 20 randomized controlled trials (RCTs) demonstrated that regular AET significantly reduces oxidative stress markers and increases antioxidant levels in older adults [14]. Furthermore, Konopka et al. examined mitochondrial quality control in young and older men. They found that 12 weeks of AET promoted mitochondrial biogenesis, as evidenced by increased levels of related proteins and elevated metabolic enzyme activity in both age groups [15]. Interestingly, the authors also reported an increase in markers of both mitochondrial fusion and fission following AET, regardless of age [15]. However, the effects of AET on mitophagy and selective removal of damaged mitochondria remain a topic of debate [16]. Researchers found in one study of an aged animal model of sarcopenia a decrease in mitophagic flux, potentially due to improved overall mitochondrial quality following endurance exercise training [17]; another report contradicted this finding. These results suggest that 4 to 5 weeks of voluntary wheel-running exercise promotes elevated basal mitophagy in trained skeletal muscles, potentially reflecting a heightened rate of mitochondrial turnover in skeletal muscle [18]. In contrast, RET research has primarily focused on adaptations related to the enhancement of skeletal muscle hypertrophy and strength [19]. The impact of RET on mitochondrial adaptation in aging skeletal muscles remains inconclusive. Traditionally viewed as less effective than endurance exercise in improving mitochondrial function, recent studies have reported enhanced markers of mitochondrial content and function following RET in both young [20,21] and old populations [22,23,24]. Further research is required to determine how much RET contributes to mitochondrial adaptation.
In this review, we discuss the underappreciated role of RET in aging skeletal muscle mitochondria, offering a new perspective on its therapeutic benefits in combating age-related sarcopenia. We summarize the effects of RET on mitochondria in aging skeletal muscles and focus on mitochondrial reactive oxygen species (mtROS), biogenesis, dynamics, and mitophagy, exploring the potential underlying mechanisms.
2. Role of Mitochondria in Sarcopenia
Skeletal muscle loss is influenced by several factors and pathways, including chronic inflammation, reduced growth hormone signaling, and inadequate protein intake [25,26,27]. Furthermore, sarcopenia entails a complex interplay of factors, such as mitochondrial dysfunction, leading to impaired skeletal muscle fiber energy production, malfunctioning muscle satellite cell function, and neurological impairments affecting motor unit recruitment and innervation [28]. Recent research suggests that changes in mitochondrial biogenesis, dynamics, function, and structure may be the primary factors influencing skeletal muscle quality and performance [29]. Understanding how these factors affect skeletal muscle metabolism is key to developing effective strategies for preventing and treating sarcopenia and, ultimately, promoting healthy aging.
2.1. Mitochondrial Reactive Oxygen Species and Mitochondrial Respiration
The overexpression of mtROS causes oxidative stress and is a major contributor to skeletal muscle weakness and loss during sarcopenia. mtROS are produced as a by-product of the electron transport chain (ETC). The excessive production of mtROS leads to loss of muscle mass and skeletal muscle strength, protein damage, mitochondrial dysfunction, and mitochondrial DNA (mtDNA) damage [30]. Skeletal muscle fibers experiencing higher levels of oxidative stress appear to have a greater prevalence of mtDNA deletions and mutations. The mtDNA genome is particularly vulnerable to mutational damage because of its proximity to the electron transport system, a major source of free radicals [31,32]. The accumulation of unrepaired mtDNA damage can exacerbate mitochondrial dysfunction by promoting increased mtROS production [33]. mtDNA mutations can impair mitochondrial respiration by damaging ETC complexes, leading to the presence of defective subunits within the ETC, which disrupts oxidative phosphorylation, reduces ATP synthesis, and elevates reactive oxygen species (ROS) generation [34,35,36,37,38]. Consequently, mitochondrial respiration, the cellular process that utilizes oxygen to generate ATP, is further impaired by mtDNA mutations [37,39].
Studies suggest elevated mtROS and impaired skeletal muscle mitochondrial respiration are associated with age-related sarcopenia [40,41]. For example, Grevendonk et al. investigated the impact of aging on mitochondrial function in older and young individuals maintaining similar levels of habitual physical activity [42]. To assess skeletal muscle mitochondrial respiration, the researchers employed high-resolution respirometry with permeabilized skeletal muscle fibers. They found approximately 15% lower ADP-stimulated mitochondrial respiration rate in older adults relative to young adults. However, no significant difference in mitochondrial content, assessed by mitochondrial oxidative phosphorylation (OXPHOS) protein levels, was observed between the groups [42]. Similarly, Joseph et al. observed a more specific association between low physical function and mitochondrial dysfunction in aging muscle [43]. They reported a decline in mitochondrial respiration, potentially leading to higher free radical formation and subsequent muscle damage, in elderly individuals with low physical function [43]. Furthermore, González-Blanco et al. reported that severely functional-dependent patients with extreme sarcopenia have lower expression of the main complexes of the ETC [44]. This finding suggests a decline in mitochondrial function and a reduced ability to produce energy (ATP) within muscle cells. Fan et al. reported higher levels of circulating cell-free mitochondrial DNA (ccf-mtDNA) in older adults with sarcopenia [45]. ccf-mtDNA, a short segment of mtDNA derived from cells, is released into systemic circulation in response to cellular injury or stress [46]. Elevated ccf-mtDNA levels may indicate mitochondrial damage due to abnormal mitochondrial leakage [45]. Damage to mtDNA caused by ROS is thought to initiate a cycle of increasing oxidative stress within the mitochondria [47].
2.2. Mitochondrial Biogenesis
Reduced mitochondrial biogenesis, which involves not only generating new mitochondria but also increasing their size and mass, is a key factor in the loss of skeletal muscle quality and the development of sarcopenia [29,48,49]. Mitochondrial biogenesis is a complex process that involves building proteins encoded by mtDNA, importing nuclear-encoded mitochondrial proteins, and replicating mtDNA [50]. Studies have shown that peroxisome proliferator-activated receptor-gamma coactivator 1 alpha (PGC-1α) plays a pivotal role in mitochondrial biogenesis by regulating the transcriptional machinery responsible for increasing mitochondrial mass [51,52]. The levels of PGC-1α protein and mRNA decline with age, and this decrease is associated with skeletal muscle weakness and poor exercise performance in older adults [49]. Joseph et al. demonstrated that lower physical function in older adults correlates with weaker mitochondrial function and reduced PGC-1α protein levels compared with both younger adults and high-functioning older adults [43]. Cannavino et al. reported that mice genetically modified to have high levels of PGC-1α maintained skeletal muscle mass even when the hindlimb was unloaded, potentially by suppressing autophagy and proteasome degradation pathways that break down skeletal muscle proteins [53]. However, a recent study investigating hindlimb unloading in mice overexpressing PGC-1α did not observe a protective effect against disuse atrophy with respect to muscle mass or cross-sectional area [54]. This discordance in results may be attributable to differences in the age of the mice used in the studies [54]. Aging in skeletal muscles is associated with altered mitochondrial biogenesis pathways, which may contribute to skeletal muscle loss and reduced physical function commonly observed in older adults [43].
2.3. Mitochondrial Dynamics
Mitochondrial dysfunction, caused by altered dynamics and morphology, is a major contributor to the decline in skeletal muscle function observed in sarcopenia [29,55]. Mitochondria are dynamic organelles that constantly undergo fusion and fission, which are important for maintaining mitochondrial function. Mitochondrial fusion is the merging of two mitochondria at the interfaces of their outer and inner membranes. In contrast, mitochondrial fission refers to the process by which a mitochondrion is divided into two separate mitochondria [56]. The constant interplay between fusion and fission determines the size, shape, and number of mitochondria in a cell [57]. In healthy cells, these processes are balanced to maintain the mitochondrial morphology. Mitochondrial fusion occurs in a two-step process. The fusion process involves the facilitation of the outer mitochondrial membrane by Mitofusin 1 (Mfn1) and Mitofusin 2 (Mfn2), followed by the mediation of inner mitochondrial membrane fusion by the Optic Atrophy 1 protein (OPA1) [8]. Mitochondrial fission involves dynamin-related protein 1 (Drp1), which forms constricting rings around the organelle and is recruited by mitochondrial Fission-1 protein (Fis1) to the outer membrane [57]. The results of recent studies suggest that disruptions in the balance of mitochondrial dynamics can impair mitochondrial function and health in skeletal muscles [29,56,57]. This imbalance is associated with senescence and muscle atrophy. For example, Sebastián et al. reported that Mfn2 deletion leads to the accumulation of damaged mitochondria, increased production of harmful molecules, impaired skeletal muscle function, reduced energy production, and general weakness [58]. In addition, Favaro et al. showed that the absence of Drp1 leads to abnormal enlargement and dysfunction of mitochondria and causes skeletal muscle atrophy and degeneration [59]. Dulac et al. demonstrated that Drp1 knockdown in middle-aged mice leads to severe skeletal muscle atrophy, oxidative stress, and impaired autophagy, whereas Drp1 overexpression results in mild skeletal muscle atrophy and negatively affects mitochondrial quality [60]. This suggests that Drp1 content in skeletal muscle must be maintained within a certain physiological range to ensure that the rate of mitochondrial fission is neither excessive nor deficient [60].
2.4. Mitophagy
Mitophagy selectively removes damaged or unnecessary mitochondria through the autophagy-lysosomal system. Initiated post-mitochondrial fission, this process facilitates the encapsulation of smaller mitochondrial fragments for the selective removal of damaged mitochondria, essentially constituting a “quality control” mechanism [61]. Cells employ various mechanisms for mitophagy, including ubiquitin- and receptor-dependent pathways [62]. Several authors have proposed correlations between alterations in signaling pathways associated with autophagy and mitophagy and skeletal muscle atrophy and decreased physical function in the elderly. For instance, Picca et al. observed elevated levels of the autophagy protein p62 and the mitophagy-targeting protein BCL2/adenovirus E1B 19 kDa protein-interacting protein-3 (BNIP3) in older adults compared with younger controls, linking them to increased mitochondrial dysfunction [63]. However, Drummond et al. reported reduced expression levels of skeletal muscle mitophagy regulators BNIP3, Drp1, and Parkin in sedentary older women, attributing it to mitochondrial dysfunction [64]. Ito et al. demonstrated the crucial role of Parkin-mediated mitophagy in modulating skeletal muscle myotube atrophy by regulating mtROS production [65]. While there are some contradictory data on mitophagy, the literature results increasingly suggest a decline in mitophagy in skeletal muscle with age, which is associated with impaired skeletal muscle function. Consequently, targeting mitophagy in skeletal muscle emerges as crucial for preserving mitochondrial function and skeletal muscle mass.
3. Effects of Resistance Exercise Training on Aging Skeletal Muscle Mitochondria
Exercise is recognized as the most effective intervention for sarcopenia, as no specific drugs have been approved for its treatment. Evidence-based clinical practice guidelines strongly recommend physical activity as the primary treatment for sarcopenia [66]. Traditionally, AET has been recommended to improve mitochondrial adaptations [11]. However, a growing body of research suggests that RET also has a positive effect on mitochondria [67]. Here, we briefly describe the effects of RET on aging skeletal muscle mitochondria in relation to sarcopenia (Table 1).

Table 1.
Effects of resistance exercise training on skeletal muscle mitochondria in the elderly.
3.1. Mitochondrial Reactive Oxygen Species and Mitochondrial Respiration
The effects of exercise-induced oxidative stress remain a topic of ongoing debate. The relationship between physical activity, the generation of ROS, and their potential impact is a complex and multifaceted issue. Exercise-induced ROS production can have both beneficial and detrimental effects. Moderate levels of ROS generated during exercise can promote positive physiological adaptations in active skeletal muscles. These adaptations include stimulating mitochondrial biogenesis, increasing the synthesis of antioxidant enzymes, and upregulating stress response proteins [77,78,79,80]. On the other hand, high levels of ROS production during exercise can damage macromolecular structures such as proteins, lipids, and DNA [77,78,81]. While speculation exists regarding the hormetic effect of exercise-induced ROS production in skeletal muscles, insufficient evidence suggests that extended periods of high-intensity exercise lead to tissue damage and compromised physiological functions [82,83]. Numerous studies consistently indicate that prolonged high-intensity exercise yields significant health advantages [82,84].
The widely accepted antioxidant benefits of exercise training have led some authors to propose that exercise itself can be considered an effective antioxidant [85,86] (Figure 1). Parise et al. examined oxidative stress, antioxidant enzyme protein levels, mitochondrial enzymes, and mtDNA levels in older adults who underwent RET for 14 weeks [68]. The researchers reported that RET did not significantly alter the levels of antioxidant proteins. However, they observed a significant attenuation in DNA damage caused by oxidative stress. An analysis of mitochondrial enzyme activity revealed a significant increase in complex IV activity, suggesting a potential indirect antioxidant role for complex IV after RET. Electron leakage from the ETC can directly interact with oxygen, resulting in enhanced ROS [87]. However, efficient electron transfer at complex IV reduces ROS formation in mitochondria by ensuring complete oxygen reduction and preventing electron leaks from upstream complexes [36,88].
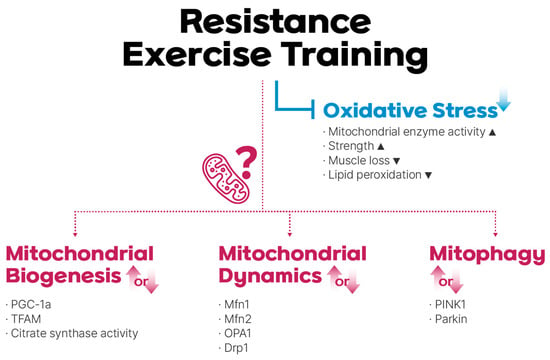
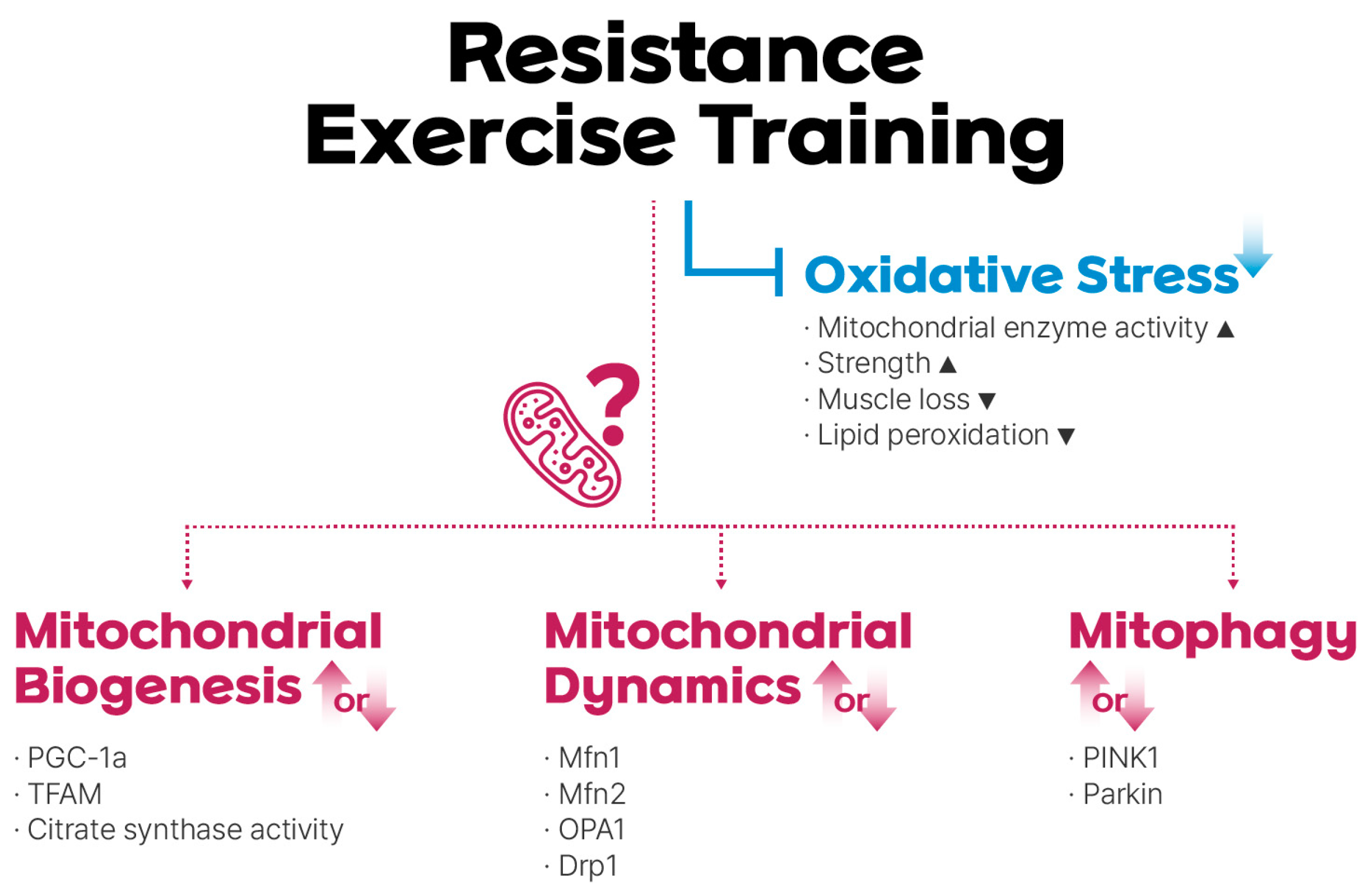
Figure 1.
Resistance exercise training (RET)-mediated regulation of mitochondrial reactive oxidative stress (mtROS), biogenesis, dynamics, and mitophagy. RET reduces oxidative stress, although the role of mitochondrial adaptations in this process remains under debate.
In a study where older adults underwent 12 weeks of moderate-intensity RET, indicators of oxidative stress were measured in blood and urine samples [89]. The authors reported that moderate-intensity RET resulted in a significant decrease in ROS production by nonenzymatic antioxidant capacity and a significant decrease in all measures of oxidative stress, including the accumulation of oxidized proteins, degree of lipid peroxidation, and oxidative DNA damage. Interestingly, the authors of this study suggested that reduced oxidative stress may be associated with increased strength and a slower rate of skeletal muscle loss.
More recently, Mesquita et al. examined the effect of 6 weeks of RET on the mRNA expression, protein levels, and enzymatic activity of several endogenous antioxidants in the skeletal muscle of 13 older males [90]. Additionally, lipid peroxidation and levels of heat-shock proteins, known for their protective role against oxidative stress, were measured. The results showed that protein levels varied, with enzymes such as catalase showing a decrease, whereas other enzymes remained unchanged. However, mRNA expression and enzymatic activity increased, and lipid peroxidation decreased. The researchers concluded that RET may be a viable approach to counteract age-related disruptions in muscle redox homeostasis through multilevel regulation of the antioxidant system.
In contrast to research suggesting that resistance exercise reduces oxidative stress, two studies found no significant improvement in mitochondrial respiratory function following RET [23,72]. Notably, these studies assessed mitochondrial respiration in skeletal muscle using high-resolution respirometry. Irving et al. investigated the effects of 8 weeks of combined exercise training (CET) compared with AET and RET on mitochondrial physiology in both young and older adults [72]. Interestingly, while all three training modalities improved muscle strength and cardiorespiratory fitness, only AET and CET led to enhanced mitochondrial respiratory function. Similarly, Robinson et al. investigated the effects of high-intensity aerobic interval training (HIIT), RET, and CET on skeletal muscle adaptations in young and older adults following a 12-week intervention [23]. This study revealed that HIIT significantly enhanced maximal mitochondrial respiration in both young and older adults, with a greater increase observed in the older group. Conversely, CET only led to a significant rise in young adults, with no improvement in the older population. RET did not elicit a statistically significant change in mitochondrial respiration for either age group. Researchers explored the potential link between changes in mitochondrial protein synthesis rate and alterations in mitochondrial respiration. Their investigation revealed no significant difference in the baseline mitochondrial protein synthesis rate between young and older participants. However, HIIT intervention led to a significant increase in the mitochondrial protein synthesis rate for both young and older groups.
3.2. Mitochondrial Biogenesis
Exercise is an effective stimulus for mitochondrial biogenesis, a process that involves intricate molecular pathways that promote mitochondrial growth [19,91,92,93,94]. PGC-1α is a critical regulator of mitochondrial biogenesis in skeletal muscle [95]. Exercise has been shown to increase PGC-1α expression and activity, which in turn interacts with nuclear respiratory factor 1 (Nrf1) and 2 (Nrf2) and mitochondrial transcription factor A (TFAM) to promote the transcription of mtDNA and enhance mitochondrial biogenesis [50,95]. Mitochondrial biogenesis can be measured by various methods. The first approach analyzes the expression of regulatory markers such as PGC-1α, Nrf1/2, and TFAM, using techniques such as real-time polymerase chain reaction (qPCR) or Western blotting [96]. Researchers often use the mtDNA copy number as an indicator of mitochondrial biogenesis and content, which can be quantified by qPCR [71,97]. A more accurate but complex approach involves measuring the mitochondrial content or synthesis [96]. Citrate synthase (CS) activity and the protein content of OXPHOS are commonly used as markers of mitochondrial content [98,99,100,101]. CS activity has been shown to correlate with mitochondrial volume density as measured by electron microscopy, offering a considerably faster and more practical method [100].
Although mitochondrial biogenesis that occurs in response to AET has been well studied, the effects of RET on mitochondrial adaptation remain controversial (Figure 1). Long-term (“chronic”) RET has been shown to increase the mtDNA copy number, mitochondrial contents, and PGC-1α mRNA and protein contents [70,71,72]. In a study by Ogborn et al., both young and elderly individuals underwent one-time unilateral RET [73]; the researchers observed significant increases in PGC-1α mRNA levels 3 h after exercise, followed by a rise in TFAM mRNA 24 h later in the vastus lateralis muscle. Notably, there were no age-related differences in the mitochondrial response.
It should be noted that there are conflicting findings in terms of the response of mitochondrial biogenesis and mitochondrial content to RET in older adults. Parise et al. examined the effects of unilateral RET on mitochondrial enzymes in the skeletal muscles of older adults [69]. CS activity was measured in the vastus lateralis muscle of both trained and untrained legs. Over the 12-week training period, neither the trained nor the untrained leg showed any changes in CS activity. This finding adds to the growing body of evidence suggesting no change in CS activity following RET [68,74,75]. Similarly, in three recent investigations by Flack et al., Mesquita et al., and Berg et al., the authors observed no change in the levels of mitochondrial biogenesis proteins PGC-1α, TFAM, and total OXPHOS, or mRNA expression of PGC-1α and TFAM in vastus lateralis muscle after 8–12 weeks of RET [22,74,75]. These results suggest that unchanged mitochondrial protein expression, despite greater muscle volume after RET, indicates that mitochondrial biogenesis may have occurred [75]. Moreover, decreased maximal mitochondrial respiratory rate suggests potential qualitative adaptations, such as an increased proton leak and changes in cristae morphology, rather than a simple increase in mitochondrial content [75].
3.3. Mitochondrial Dynamics
The effects of exercise on mitochondrial dynamics, including fusion and fission, have not been extensively studied [102]. Although it is expected that changes in cellular energy status during exercise can be achieved through mitochondrial network dynamics, the specific effects of exercise on these mitochondrial dynamic processes require further investigation [103] (Figure 1). Available evidence from a limited number of studies suggests that physical activity influences mitochondrial dynamics via multiple signaling pathways. For example, high-intensity interval training appears to induce a progressive increase in Mfn1 and Fis1 protein levels, whereas single exercise bouts result in increased mRNA expression of Mfn1 and Mfn2 at specific time points post-exercise [104,105,106]. Several methods exist to quantify protein or mRNA levels associated with mitochondrial fusion and fission. These include quantifying the levels of proteins using techniques such as Western blot analysis, immunofluorescence analysis, and quantitative immunofluorescence analysis or assessing mRNA levels using quantitative reverse transcription-polymerase chain reaction (RT-qPCR) [96].
There are relatively few investigations of mitochondrial dynamics in the skeletal muscles of older adults during RET, with most studies relying on molecular biology analyses. Recently, researchers have reported the response of mitochondrial dynamics markers following either acute or chronic RET in older adults [22]. Researchers reported no acute changes in mitochondrial dynamics-related proteins following RET. However, chronic RET (over 10 weeks) led to an increase in Mfn1, Mfn2, OPA1, and Drp1 protein levels. The authors suggest that the age-related increase in mitochondrial fusion observed might be a compensatory response to counteract the negative effects associated with aging. Age-related increases in mitochondrial fission led to fragmented networks and dysfunction [107,108]. The authors propose that fusion could act to dilute damaged mitochondrial contents by combining them with healthier mitochondria. Additionally, their data show that RET counteracts age-related dysregulation of mitochondrial dynamics, as evidenced by changes in markers of mitochondrial fusion, potentially leading to improved mitochondrial function.
In contrast, Marshall et al. did not observe significant effects after a single bout of RET on markers of mitochondrial fission/fusion dynamics [76] in a study comparing post-exercise recovery of master endurance athletes with that of untrained healthy age-matched older adults at both the 1- and 48-h time points. The researchers found that the only significant effect of RET was observed in the phosphorylation/total Drp1 ratio, a marker of mitochondrial fission, at the 1-h post-exercise time point. Importantly, this effect was observed only in the master endurance athlete group, with no changes in this or any other mitochondrial dynamics marker detected in the control group. Collectively, these findings suggest that master endurance athletes maintain more efficient mechanisms for skeletal muscle mitochondrial quality control, facilitating the fragmentation of damaged or dysfunctional organelles. Given the conflicting findings in the recent literature, the influence of RET on mitochondrial volume in older adults requires further investigation.
3.4. Mitophagy
The signaling pathway mediated by PTEN-induced putative kinase protein 1 (PINK1) and Parkin is recognized as an extensively investigated pathway responsible for transmitting signals related to mitochondrial damage, ultimately leading to the initiation of mitophagy [109]. Mitophagy is also partially mediated by BNIP3 and BNIP3L/NIX, which are located on the outer mitochondrial membrane [110,111]. Exercise induces mitophagy through various mechanisms, including the PINK1/Parkin pathway [112]. A recent cross-sectional study found that older individuals who engaged in regular exercise exhibited elevated levels of key proteins involved in mitophagy, specifically Parkin and BNIP3 [113]. Although some studies have suggested that exercise promotes mitophagy, the overall effect on this process during aging requires further investigation [16] (Figure 1). Studies have shown that Parkin protein levels may remain unchanged or increase at the mRNA level in elderly individuals in response to exercise [64,114]. This suggests that exercise may not strongly stimulate the PINK1/Parkin pathway in this population. However, PINK1 did not show a clear response, with some studies reporting no significant changes [114,115]. The current methods for assessing mitophagy are limited. Transmission electron microscopy (TEM) and Western blot analysis of mitochondrial protein levels are established methods for directly visualizing and quantifying mitophagy [96,116]. However, these methods lack accuracy and the ability to specifically quantify mitophagy [96,116]. In comparison, fluorescent-based strategies offer enhanced reliability and potential for live-cell imaging [96].
One limitation of the existing literature is the paucity of studies investigating the effects of RET on mitophagy in older adults. The previously mentioned study by Ogborn et al. compared the acute response of young and aged skeletal muscle mitochondria to RET [73]. They found that the levels of PINK1 and Parkin proteins in the cytosolic fraction remained unchanged after exercise, and neither age nor exercise regimen appeared to influence their interactions significantly. Given this confirmation of a mitophagy response in the mitochondria following acute RET exposure, it is critical to determine if this response persists in chronic settings [117].
A recent study by Mesquita et al. further supported these findings by demonstrating a lack of statistically significant differences in the mitophagic response of skeletal muscles in older adults following acute and chronic RET [22]. There was no change in the levels of PINK1 and Parkin proteins in the vastus lateralis tissue either 24 h after the first RET session or 72 h after 10 weeks of RET. In fact, the authors argued that the results presented do not definitively show unaltered mitophagy, as the study did not analyze the levels of phosphorylated PINK1 and Parkin proteins, which can affect their activity [118]. These results suggest that RET does not significantly affect mitophagy markers in older individuals. This finding is in contrast with the increased PINK1 protein levels observed in young skeletal muscles following chronic RET [21].
4. Conclusions
The findings of this review indicate that mitochondrial dysfunction contributes to the acceleration of age-related sarcopenia (Figure 2). Specifically, mtROS production, mitochondrial biogenesis, mitochondrial dynamics, and mitophagy are implicated in sarcopenia. Previous studies have demonstrated that AET reduces oxidative stress and enhances mitochondrial biogenesis and dynamics. While RET has also emerged as a promising preventive and therapeutic approach for improving muscle strength and potentially attenuating sarcopenia, much of the research on RET has focused on muscle hypertrophy and strength gains, with relatively few studies investigating mitochondrial responses to RET. Despite conflicting findings regarding the effects of RET on mitochondria in the elderly, numerous studies have reported the beneficial effects of RET on mitochondrial function in aging skeletal muscles. Further investigations are imperative to elucidate the specific mechanisms through which RET influences mitochondrial adaptation in aging skeletal muscles.
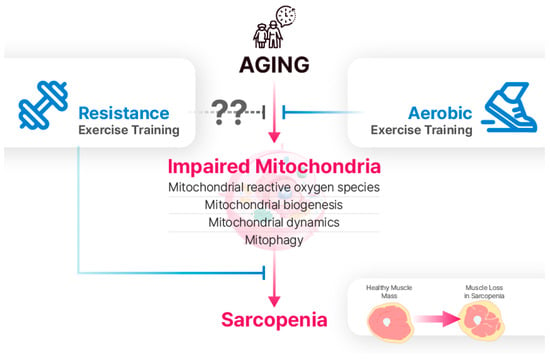
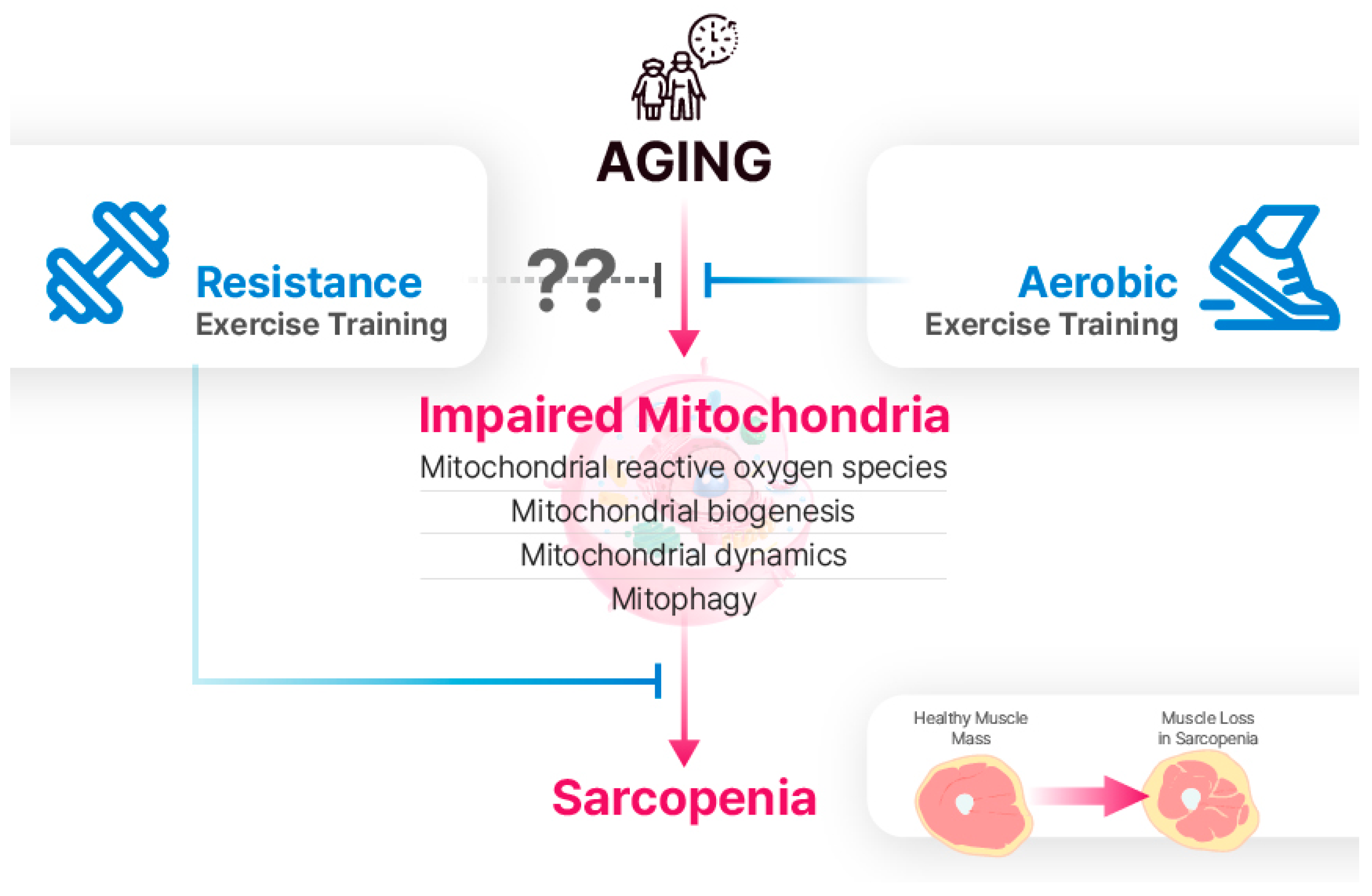
Figure 2.
Aging leads to the impairment of mitochondrial reactive oxygen species (mtROS), biogenesis, dynamics, and mitophagy. Although the effects of RET on mitochondria in aging skeletal muscle are not fully understood, it is a well-established intervention for sarcopenia. While AET is traditionally viewed as more effective for mitochondrial adaptations, recent research suggests that RET can also have positive effects on mitochondrial function.
Author Contributions
I.J., E.-J.C. and H.-B.K. contributed to the conception and design of the study. I.J., E.-J.C., J.-S.Y., Y.C., D.-H.P., J.-H.K., S.-H.L., D.-Y.S., S.-J.J. and H.-B.K. conducted the critical discussion. I.J., E.-J.C. and H.-B.K. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Ministry of Education of the Republic of Korea and the National Research Foundation of Korea (NRF-2022S1A5C2A03092407).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Colloca, G.; Di Capua, B.; Bellieni, A.; Fusco, D.; Ciciarello, F.; Tagliaferri, L.; Valentini, V.; Balducci, L. Biological and Functional Biomarkers of Aging: Definition, Characteristics, and How They Can Impact Everyday Cancer Treatment. Curr. Oncol. Rep. 2020, 22, 115. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Van Ancum, J.M.; Alcazar, J.; Meskers, C.G.M.; Nielsen, B.R.; Suetta, C.; Maier, A.B. Impact of using the updated EWGSOP2 definition in diagnosing sarcopenia: A clinical perspective. Arch. Gerontol. Geriatr. 2020, 90, 104125. [Google Scholar] [CrossRef] [PubMed]
- Anjanappa, M.; Corden, M.; Green, A.; Roberts, D.; Hoskin, P.; McWilliam, A.; Choudhury, A. Sarcopenia in cancer: Risking more than muscle loss. Tech. Innov. Patient Support Radiat. Oncol. 2020, 16, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Guo, Z.; Jiang, B.; Zhang, K.; Zhu, W.; Lian, X.; Xu, Y.; Zhao, Z.; Liu, L. Factors affecting sarcopenia in older patients with chronic diseases. Ann. Palliat. Med. 2022, 11, 972–983. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.Z.; No, M.H.; Heo, J.W.; Park, D.H.; Kang, J.H.; Kim, S.H.; Kwak, H.B. Role of exercise in age-related sarcopenia. J. Exerc. Rehabil. 2018, 14, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Annesley, S.J.; Fisher, P.R. Mitochondria in Health and Disease. Cells 2019, 8, 680. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ji, Y.; Liu, R.; Zhu, X.; Wang, K.; Yang, X.; Liu, B.; Gao, Z.; Huang, Y.; Shen, Y.; et al. Mitochondrial dysfunction: Roles in skeletal muscle atrophy. J. Transl. Med. 2023, 21, 503. [Google Scholar] [CrossRef] [PubMed]
- Migliavacca, E.; Tay, S.K.H.; Patel, H.P.; Sonntag, T.; Civiletto, G.; McFarlane, C.; Forrester, T.; Barton, S.J.; Leow, M.K.; Antoun, E.; et al. Mitochondrial oxidative capacity and NAD(+) biosynthesis are reduced in human sarcopenia across ethnicities. Nat. Commun. 2019, 10, 5808. [Google Scholar] [CrossRef]
- Ruegsegger, G.N.; Booth, F.W. Health Benefits of Exercise. Cold Spring Harb. Perspect. Med. 2018, 8, a029694. [Google Scholar] [CrossRef]
- Mang, Z.A.; Ducharme, J.B.; Mermier, C.; Kravitz, L.; de Castro Magalhaes, F.; Amorim, F. Aerobic Adaptations to Resistance Training: The Role of Time under Tension. Int. J. Sports Med. 2022, 43, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Joyner, M.J.; Coyle, E.F. Endurance exercise performance: The physiology of champions. J. Physiol. 2008, 586, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.D.; Hwa Joo, C.; Jeong, T.S.; Louhelainen, J.; Cochran, A.J.; Gibala, M.J.; Gregson, W.; Close, G.L.; Drust, B.; Morton, J.P. Matched work high-intensity interval and continuous running induce similar increases in PGC-1alpha mRNA, AMPK, p38, and p53 phosphorylation in human skeletal muscle. J. Appl. Physiol. 2012, 112, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Lin, H.; Wan, M.; Qiu, P.; Xia, R.; He, J.; Tao, J.; Chen, L.; Zheng, G. The Effects of Aerobic Exercise on Oxidative Stress in Older Adults: A Systematic Review and Meta-Analysis. Front. Physiol. 2021, 12, 701151. [Google Scholar] [CrossRef] [PubMed]
- Konopka, A.R.; Suer, M.K.; Wolff, C.A.; Harber, M.P. Markers of Human Skeletal Muscle Mitochondrial Biogenesis and Quality Control: Effects of Age and Aerobic Exercise Training. J. Gerontol. Ser. A 2013, 69, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Moreira, O.C.; Estébanez, B.; Martínez-Florez, S.; Paz, J.A.d.; Cuevas, M.J.; González-Gallego, J. Mitochondrial Function and Mitophagy in the Elderly: Effects of Exercise. Oxidative Med. Cell. Longev. 2017, 2017, 2012798. [Google Scholar] [CrossRef] [PubMed]
- Carter, H.N.; Kim, Y.; Erlich, A.T.; Zarrin-Khat, D.; Hood, D.A. Autophagy and mitophagy flux in young and aged skeletal muscle following chronic contractile activity. J. Physiol. 2018, 596, 3567–3584. [Google Scholar] [CrossRef] [PubMed]
- Lira, V.A.; Okutsu, M.; Zhang, M.; Greene, N.P.; Laker, R.C.; Breen, D.S.; Hoehn, K.L.; Yan, Z. Autophagy is required for exercise training-induced skeletal muscle adaptation and improvement of physical performance. FASEB J. 2013, 27, 4184–4193. [Google Scholar] [CrossRef] [PubMed]
- Parry, H.A.; Roberts, M.D.; Kavazis, A.N. Human Skeletal Muscle Mitochondrial Adaptations Following Resistance Exercise Training. Int. J. Sports Med. 2020, 41, 349–359. [Google Scholar] [CrossRef]
- Groennebaek, T.; Jespersen, N.R.; Jakobsgaard, J.E.; Sieljacks, P.; Wang, J.; Rindom, E.; Musci, R.V.; Bøtker, H.E.; Hamilton, K.L.; Miller, B.F.; et al. Skeletal Muscle Mitochondrial Protein Synthesis and Respiration Increase With Low-Load Blood Flow Restricted as Well as High-Load Resistance Training. Front. Physiol. 2018, 9, 1796. [Google Scholar] [CrossRef]
- Lim, C.; Kim, H.J.; Morton, R.W.; Harris, R.; Phillips, S.M.; Jeong, T.S.; Kim, C.K. Resistance Exercise-induced Changes in Muscle Phenotype Are Load Dependent. Med. Sci. Sports Exerc. 2019, 51, 2578–2585. [Google Scholar] [CrossRef]
- Mesquita, P.H.C.; Lamb, D.A.; Parry, H.A.; Moore, J.H.; Smith, M.A.; Vann, C.G.; Osburn, S.C.; Fox, C.D.; Ruple, B.A.; Huggins, K.W.; et al. Acute and chronic effects of resistance training on skeletal muscle markers of mitochondrial remodeling in older adults. Physiol. Rep. 2020, 8, e14526. [Google Scholar] [CrossRef]
- Robinson, M.M.; Dasari, S.; Konopka, A.R.; Johnson, M.L.; Manjunatha, S.; Esponda, R.R.; Carter, R.E.; Lanza, I.R.; Nair, K.S. Enhanced Protein Translation Underlies Improved Metabolic and Physical Adaptations to Different Exercise Training Modes in Young and Old Humans. Cell Metab. 2017, 25, 581–592. [Google Scholar] [CrossRef]
- Ruple, B.A.; Godwin, J.S.; Mesquita, P.H.C.; Osburn, S.C.; Vann, C.G.; Lamb, D.A.; Sexton, C.L.; Candow, D.G.; Forbes, S.C.; Frugé, A.D.; et al. Resistance training rejuvenates the mitochondrial methylome in aged human skeletal muscle. FASEB J. 2021, 35, e21864. [Google Scholar] [CrossRef]
- Salles, J.; Chanet, A.; Guillet, C.; Vaes, A.M.; Brouwer-Brolsma, E.M.; Rocher, C.; Giraudet, C.; Patrac, V.; Meugnier, E.; Montaurier, C.; et al. Vitamin D status modulates mitochondrial oxidative capacities in skeletal muscle: Role in sarcopenia. Commun. Biol. 2022, 5, 1288. [Google Scholar] [CrossRef]
- Dalle, S.; Rossmeislova, L.; Koppo, K. The Role of Inflammation in Age-Related Sarcopenia. Front. Physiol. 2017, 8, 1045. [Google Scholar] [CrossRef]
- Bian, A.; Ma, Y.; Zhou, X.; Guo, Y.; Wang, W.; Zhang, Y.; Wang, X. Association between sarcopenia and levels of growth hormone and insulin-like growth factor-1 in the elderly. BMC Musculoskelet. Disord. 2020, 21, 214. [Google Scholar] [CrossRef]
- Yang, Q.; Chan, P. Skeletal Muscle Metabolic Alternation Develops Sarcopenia. Aging Dis. 2022, 13, 801–814. [Google Scholar] [CrossRef]
- Bellanti, F.; Lo Buglio, A.; Vendemiale, G. Mitochondrial Impairment in Sarcopenia. Biology 2021, 10, 31. [Google Scholar] [CrossRef]
- Chen, M.; Wang, Y.; Deng, S.; Lian, Z.; Yu, K. Skeletal muscle oxidative stress and inflammation in aging: Focus on antioxidant and anti-inflammatory therapy. Front. Cell Dev. Biol. 2022, 10, 964130. [Google Scholar] [CrossRef]
- McKenzie, D.; Bua, E.; McKiernan, S.; Cao, Z.; Wanagat, J.; Aiken, J. Mitochondrial DNA deletion mutations: A causal role in sarcopenia. Eur. J. Biochem. 2002, 269, 1995–2265. [Google Scholar] [CrossRef]
- Aiken, J.; Bua, E.; Cao, Z.; Lopez, M.; Wanagat, J.; McKenzie, D.; McKiernan, S. Mitochondrial DNA deletion mutations and sarcopenia. Ann. N. Y. Acad. Sci. 2002, 959, 412–423. [Google Scholar] [CrossRef]
- Rong, Z.; Tu, P.; Xu, P.; Sun, Y.; Yu, F.; Tu, N.; Guo, L.; Yang, Y. The Mitochondrial Response to DNA Damage. Front. Cell Dev. Biol. 2021, 9, 669379. [Google Scholar] [CrossRef]
- Hebert, S.L.; Lanza, I.R.; Nair, K.S. Mitochondrial DNA alterations and reduced mitochondrial function in aging. Mech. Ageing Dev. 2010, 131, 451–462. [Google Scholar] [CrossRef]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial electron transport chain: Oxidative phosphorylation, oxidant production, and methods of measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef]
- Raimondi, V.; Ciccarese, F.; Ciminale, V. Oncogenic pathways and the electron transport chain: A dangeROS liaison. Br. J. Cancer 2020, 122, 168–181. [Google Scholar] [CrossRef]
- Lu, J.; Sharma, L.K.; Bai, Y. Implications of mitochondrial DNA mutations and mitochondrial dysfunction in tumorigenesis. Cell Res. 2009, 19, 802–815. [Google Scholar] [CrossRef]
- Mito, T.; Kikkawa, Y.; Shimizu, A.; Hashizume, O.; Katada, S.; Imanishi, H.; Ota, A.; Kato, Y.; Nakada, K.; Hayashi, J. Mitochondrial DNA mutations in mutator mice confer respiration defects and B-cell lymphoma development. PLoS ONE 2013, 8, e55789. [Google Scholar] [CrossRef]
- Szczepanowska, J.; Malinska, D.; Wieckowski, M.R.; Duszynski, J. Effect of mtDNA point mutations on cellular bioenergetics. Biochim. Biophys. Acta BBA—Bioenerg. 2012, 1817, 1740–1746. [Google Scholar] [CrossRef]
- Hadjispyrou, S.; Giannopoulos, A.; Philippou, A.; Theos, A. Mitochondrial Dysfunction and Sarcopenic Obesity: The Role of Exercise. J. Clin. Med. 2023, 12, 5628. [Google Scholar] [CrossRef]
- Affourtit, C.; Carré, J.E. Mitochondrial involvement in sarcopenia. Acta Physiol. 2024, 240, e14107. [Google Scholar] [CrossRef] [PubMed]
- Grevendonk, L.; Connell, N.J.; McCrum, C.; Fealy, C.E.; Bilet, L.; Bruls, Y.M.H.; Mevenkamp, J.; Schrauwen-Hinderling, V.B.; Jörgensen, J.A.; Moonen-Kornips, E.; et al. Impact of aging and exercise on skeletal muscle mitochondrial capacity, energy metabolism, and physical function. Nat. Commun. 2021, 12, 4773. [Google Scholar] [CrossRef]
- Joseph, A.M.; Adhihetty, P.J.; Buford, T.W.; Wohlgemuth, S.E.; Lees, H.A.; Nguyen, L.M.; Aranda, J.M.; Sandesara, B.D.; Pahor, M.; Manini, T.M.; et al. The impact of aging on mitochondrial function and biogenesis pathways in skeletal muscle of sedentary high- and low-functioning elderly individuals. Aging Cell 2012, 11, 801–809. [Google Scholar] [CrossRef]
- Gonzalez-Blanco, L.; Bermudez, M.; Bermejo-Millo, J.C.; Gutierrez-Rodriguez, J.; Solano, J.J.; Antuna, E.; Menendez-Valle, I.; Caballero, B.; Vega-Naredo, I.; Potes, Y.; et al. Cell interactome in sarcopenia during aging. J. Cachexia Sarcopenia Muscle 2022, 13, 919–931. [Google Scholar] [CrossRef]
- Fan, Z.; Yang, J.Y.; Guo, Y.; Liu, Y.X.; Zhong, X.Y. Altered levels of circulating mitochondrial DNA in elderly people with sarcopenia: Association with mitochondrial impairment. Exp. Gerontol. 2022, 163, 111802. [Google Scholar] [CrossRef]
- Blalock, Z.N.; Wu, G.W.Y.; Lindqvist, D.; Trumpff, C.; Flory, J.D.; Lin, J.; Reus, V.I.; Rampersaud, R.; Hammamieh, R.; Gautam, A.; et al. Circulating cell-free mitochondrial DNA levels and glucocorticoid sensitivity in a cohort of male veterans with and without combat-related PTSD. Transl. Psychiatry 2024, 14, 22. [Google Scholar] [CrossRef] [PubMed]
- Brunetta, H.S.; Holwerda, A.M.; van Loon, L.J.C.; Holloway, G.P. Mitochondrial ROS and Aging: Understanding Exercise as a Preventive Tool. J. Sci. Sport Exerc. 2020, 2, 15–24. [Google Scholar] [CrossRef]
- Jornayvaz, F.R.; Shulman, G.I. Regulation of mitochondrial biogenesis. Essays Biochem. 2010, 47, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Fan, Y.B.; Tao, X.H.; Pan, W.L.; Wu, Y.X.; Wang, X.H.; He, Y.Q.; Xiao, W.F.; Li, Y.S. Mitochondrial Quality Control in Sarcopenia: Updated Overview of Mechanisms and Interventions. Aging Dis. 2021, 12, 2016–2030. [Google Scholar] [CrossRef]
- Chen, L.; Qin, Y.; Liu, B.; Gao, M.; Li, A.; Li, X.; Gong, G. PGC-1α-Mediated Mitochondrial Quality Control: Molecular Mechanisms and Implications for Heart Failure. Front. Cell Dev. Biol. 2022, 10, 871357. [Google Scholar] [CrossRef]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms Controlling Mitochondrial Biogenesis and Respiration through the Thermogenic Coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef]
- Li, L.; Pan, R.; Li, R.; Niemann, B.; Aurich, A.C.; Chen, Y.; Rohrbach, S. Mitochondrial biogenesis and peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) deacetylation by physical activity: Intact adipocytokine signaling is required. Diabetes 2011, 60, 157–167. [Google Scholar] [CrossRef]
- Cannavino, J.; Brocca, L.; Sandri, M.; Bottinelli, R.; Pellegrino, M.A. PGC1-α over-expression prevents metabolic alterations and soleus muscle atrophy in hindlimb unloaded mice. J. Physiol. 2014, 592, 4575–4589. [Google Scholar] [CrossRef]
- Rosa-Caldwell, M.E.; Lim, S.; Haynie, W.S.; Jansen, L.T.; Westervelt, L.C.; Amos, M.G.; Washington, T.A.; Greene, N.P. Altering aspects of mitochondrial quality to improve musculoskeletal outcomes in disuse atrophy. J. Appl. Physiol. 2020, 129, 1290–1303. [Google Scholar] [CrossRef]
- Djalalvandi, A.; Scorrano, L. Mitochondrial dynamics: Roles in exercise physiology and muscle mass regulation. Curr. Opin. Physiol. 2022, 27, 100550. [Google Scholar] [CrossRef]
- Adebayo, M.; Singh, S.; Singh, A.P.; Dasgupta, S. Mitochondrial fusion and fission: The fine-tune balance for cellular homeostasis. FASEB J. 2021, 35, e21620. [Google Scholar] [CrossRef]
- Detmer, S.A.; Chan, D.C. Functions and dysfunctions of mitochondrial dynamics. Nat. Rev. Mol. Cell Biol. 2007, 8, 870–879. [Google Scholar] [CrossRef]
- Sebastián, D.; Sorianello, E.; Segalés, J.; Irazoki, A.; Ruiz-Bonilla, V.; Sala, D.; Planet, E.; Berenguer-Llergo, A.; Muñoz, J.P.; Sánchez-Feutrie, M.; et al. Mfn2 deficiency links age-related sarcopenia and impaired autophagy to activation of an adaptive mitophagy pathway. EMBO J. 2016, 35, 1677–1693. [Google Scholar] [CrossRef]
- Favaro, G.; Romanello, V.; Varanita, T.; Andrea Desbats, M.; Morbidoni, V.; Tezze, C.; Albiero, M.; Canato, M.; Gherardi, G.; De Stefani, D.; et al. DRP1-mediated mitochondrial shape controls calcium homeostasis and muscle mass. Nat. Commun. 2019, 10, 2576. [Google Scholar] [CrossRef]
- Dulac, M.; Leduc-Gaudet, J.-P.; Cefis, M.; Ayoub, M.-B.; Reynaud, O.; Shams, A.; Moamer, A.; Nery Ferreira, M.F.; Hussain, S.N.; Gouspillou, G. Regulation of muscle and mitochondrial health by the mitochondrial fission protein Drp1 in aged mice. J. Physiol. 2021, 599, 4045–4063. [Google Scholar] [CrossRef]
- Youle, R.J.; Narendra, D.P. Mechanisms of mitophagy. Nat. Rev. Mol. Cell Biol. 2011, 12, 9–14. [Google Scholar] [CrossRef]
- Palikaras, K.; Lionaki, E.; Tavernarakis, N. Mechanisms of mitophagy in cellular homeostasis, physiology and pathology. Nat. Cell Biol. 2018, 20, 1013–1022. [Google Scholar] [CrossRef]
- Picca, A.; Triolo, M.; Wohlgemuth, S.E.; Martenson, M.S.; Mankowski, R.T.; Anton, S.D.; Marzetti, E.; Leeuwenburgh, C.; Hood, D.A. Relationship between Mitochondrial Quality Control Markers, Lower Extremity Tissue Composition, and Physical Performance in Physically Inactive Older Adults. Cells 2023, 12, 183. [Google Scholar] [CrossRef]
- Drummond, M.J.; Addison, O.; Brunker, L.; Hopkins, P.N.; McClain, D.A.; LaStayo, P.C.; Marcus, R.L. Downregulation of E3 ubiquitin ligases and mitophagy-related genes in skeletal muscle of physically inactive, frail older women: A cross-sectional comparison. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69, 1040–1048. [Google Scholar] [CrossRef]
- Ito, A.; Hashimoto, M.; Tanihata, J.; Matsubayashi, S.; Sasaki, R.; Fujimoto, S.; Kawamoto, H.; Hosaka, Y.; Ichikawa, A.; Kadota, T.; et al. Involvement of Parkin-mediated mitophagy in the pathogenesis of chronic obstructive pulmonary disease-related sarcopenia. J. Cachexia Sarcopenia Muscle 2022, 13, 1864–1882. [Google Scholar] [CrossRef]
- Dent, E.; Morley, J.E.; Cruz-Jentoft, A.J.; Arai, H.; Kritchevsky, S.B.; Guralnik, J.; Bauer, J.M.; Pahor, M.; Clark, B.C.; Cesari, M.; et al. International Clinical Practice Guidelines for Sarcopenia (ICFSR): Screening, Diagnosis and Management. J. Nutr. Health Aging 2018, 22, 1148–1161. [Google Scholar] [CrossRef]
- Porter, C.; Reidy, P.T.; Bhattarai, N.; Sidossis, L.S.; Rasmussen, B.B. Resistance Exercise Training Alters Mitochondrial Function in Human Skeletal Muscle. Med. Sci. Sports Exerc. 2015, 47, 1922–1931. [Google Scholar] [CrossRef]
- Parise, G.; Brose, A.N.; Tarnopolsky, M.A. Resistance exercise training decreases oxidative damage to DNA and increases cytochrome oxidase activity in older adults. Exp. Gerontol. 2005, 40, 173–180. [Google Scholar] [CrossRef]
- Parise, G.; Phillips, S.M.; Kaczor, J.J.; Tarnopolsky, M.A. Antioxidant enzyme activity is up-regulated after unilateral resistance exercise training in older adults. Free Radic. Biol. Med. 2005, 39, 289–295. [Google Scholar] [CrossRef]
- Balakrishnan, V.S.; Rao, M.; Menon, V.; Gordon, P.L.; Pilichowska, M.; Castaneda, F.; Castaneda-Sceppa, C. Resistance training increases muscle mitochondrial biogenesis in patients with chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2010, 5, 996–1002. [Google Scholar] [CrossRef]
- Sparks, L.M.; Johannsen, N.M.; Church, T.S.; Earnest, C.P.; Moonen-Kornips, E.; Moro, C.; Hesselink, M.K.; Smith, S.R.; Schrauwen, P. Nine months of combined training improves ex vivo skeletal muscle metabolism in individuals with type 2 diabetes. J. Clin. Endocrinol. Metab. 2013, 98, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Irving, B.A.; Lanza, I.R.; Henderson, G.C.; Rao, R.R.; Spiegelman, B.M.; Nair, K.S. Combined training enhances skeletal muscle mitochondrial oxidative capacity independent of age. J. Clin. Endocrinol. Metab. 2015, 100, 1654–1663. [Google Scholar] [CrossRef]
- Ogborn, D.I.; McKay, B.R.; Crane, J.D.; Safdar, A.; Akhtar, M.; Parise, G.; Tarnopolsky, M.A. Effects of age and unaccustomed resistance exercise on mitochondrial transcript and protein abundance in skeletal muscle of men. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R734–R741. [Google Scholar] [CrossRef] [PubMed]
- Flack, K.D.; Davy, B.M.; DeBerardinis, M.; Boutagy, N.E.; McMillan, R.P.; Hulver, M.W.; Frisard, M.I.; Anderson, A.S.; Savla, J.; Davy, K.P. Resistance exercise training and in vitro skeletal muscle oxidative capacity in older adults. Physiol. Rep. 2016, 4, e12849. [Google Scholar] [CrossRef]
- Berg, O.K.; Kwon, O.S.; Hureau, T.J.; Clifton, H.L.; Thurston, T.S.; Le Fur, Y.; Jeong, E.K.; Trinity, J.D.; Richardson, R.S.; Wang, E.; et al. Skeletal Muscle Mitochondrial Adaptations to Maximal Strength Training in Older Adults. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2020, 75, 2269–2277. [Google Scholar] [CrossRef]
- Marshall, R.N.; McKendry, J.; Smeuninx, B.; Seabright, A.P.; Morgan, P.T.; Greig, C.; Breen, L. Acute resistance exercise training does not augment mitochondrial remodelling in master athletes or untrained older adults. Front. Physiol. 2022, 13, 1097988. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.; Liu, Y.; Zhang, Z. Effects of Exercise-Induced ROS on the Pathophysiological Functions of Skeletal Muscle. Oxidative Med. Cell. Longev. 2021, 2021, 3846122. [Google Scholar] [CrossRef]
- Di Meo, S.; Napolitano, G.; Venditti, P. Mediators of Physical Activity Protection against ROS-Linked Skeletal Muscle Damage. Int. J. Mol. Sci. 2019, 20, 3024. [Google Scholar] [CrossRef]
- Ji, L.L. Exercise-induced modulation of antioxidant defense. Ann. N. Y. Acad. Sci. 2002, 959, 82–92. [Google Scholar] [CrossRef]
- Ji, L.L. Modulation of skeletal muscle antioxidant defense by exercise: Role of redox signaling. Free Radic. Biol. Med. 2008, 44, 142–152. [Google Scholar] [CrossRef]
- Tryfidou, D.V.; McClean, C.; Nikolaidis, M.G.; Davison, G.W. DNA Damage Following Acute Aerobic Exercise: A Systematic Review and Meta-analysis. Sports Med. 2020, 50, 103–127. [Google Scholar] [CrossRef]
- Al-horani, R. A Narrative Review of Exercise-Induced Oxidative Stress: Oxidative DNA Damage Underlined. Open Sports Sci. J. 2022, 15, e1875399X2202220. [Google Scholar] [CrossRef]
- Powers, S.K.; Deminice, R.; Ozdemir, M.; Yoshihara, T.; Bomkamp, M.P.; Hyatt, H. Exercise-induced oxidative stress: Friend or foe? J. Sport Health Sci. 2020, 9, 415–425. [Google Scholar] [CrossRef]
- Ji, L.L. Exercise at old age: Does it increase or alleviate oxidative stress? Ann. N. Y. Acad. Sci. 2001, 928, 236–247. [Google Scholar] [CrossRef]
- Viña, J.; Olaso-Gonzalez, G.; Arc-Chagnaud, C.; De la Rosa, A.; Gomez-Cabrera, M.C. Modulating Oxidant Levels to Promote Healthy Aging. Antioxid. Redox Signal. 2020, 33, 570–579. [Google Scholar] [CrossRef]
- El Assar, M.; Álvarez-Bustos, A.; Sosa, P.; Angulo, J.; Rodríguez-Mañas, L. Effect of Physical Activity/Exercise on Oxidative Stress and Inflammation in Muscle and Vascular Aging. Int. J. Mol. Sci. 2022, 23, 8713. [Google Scholar] [CrossRef]
- Jastroch, M.; Divakaruni, A.S.; Mookerjee, S.; Treberg, J.R.; Brand, M.D. Mitochondrial proton and electron leaks. Essays Biochem. 2010, 47, 53–67. [Google Scholar] [CrossRef]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef]
- Vezzoli, A.; Mrakic-Sposta, S.; Montorsi, M.; Porcelli, S.; Vago, P.; Cereda, F.; Longo, S.; Maggio, M.; Narici, M. Moderate Intensity Resistive Training Reduces Oxidative Stress and Improves Muscle Mass and Function in Older Individuals. Antioxidants 2019, 8, 431. [Google Scholar] [CrossRef]
- Mesquita, P.H.C.; Lamb, D.A.; Godwin, J.S.; Osburn, S.C.; Ruple, B.A.; Moore, J.H.; Vann, C.G.; Huggins, K.W.; Fruge, A.D.; Young, K.C.; et al. Effects of Resistance Training on the Redox Status of Skeletal Muscle in Older Adults. Antioxidants 2021, 10, 350. [Google Scholar] [CrossRef]
- de Sousa Fernandes, M.S.; Aidar, F.J.; da Silva Pedroza, A.A.; de Andrade Silva, S.C.; Santos, G.C.J.; dos Santos Henrique, R.; Clemente, F.M.; Silva, A.F.; de Souza, R.F.; Ferreira, D.J.; et al. Effects of aerobic exercise training in oxidative metabolism and mitochondrial biogenesis markers on prefrontal cortex in obese mice. BMC Sports Sci. Med. Rehabil. 2022, 14, 213. [Google Scholar] [CrossRef]
- Kim, K.-I.; An, S.-M.; Park, H.-G.; Lee, K.-m.; Lee, W.-L. The Effect of either Aerobic Exercise Training or Chrysin Supplementation on Mitochondrial Biogenesis in Skeletal Muscle of High Fat Diet-Induced Obese Mice. Exerc. Sci. 2019, 28, 365–372. [Google Scholar] [CrossRef]
- Andrade-Souza, V.A.; Ghiarone, T.; Sansonio, A.; Santos Silva, K.A.; Tomazini, F.; Arcoverde, L.; Fyfe, J.; Perri, E.; Saner, N.; Kuang, J.; et al. Exercise twice-a-day potentiates markers of mitochondrial biogenesis in men. FASEB J. 2020, 34, 1602–1619. [Google Scholar] [CrossRef]
- Short, K.R.; Vittone, J.L.; Bigelow, M.L.; Proctor, D.N.; Rizza, R.A.; Coenen-Schimke, J.M.; Nair, K.S. Impact of aerobic exercise training on age-related changes in insulin sensitivity and muscle oxidative capacity. Diabetes 2003, 52, 1888–1896. [Google Scholar] [CrossRef]
- Fernandez-Marcos, P.J.; Auwerx, J. Regulation of PGC-1α, a nodal regulator of mitochondrial biogenesis. Am. J. Clin. Nutr. 2011, 93, 884S–890S. [Google Scholar] [CrossRef]
- You, W.; Li, Y.; Liu, K.; Mi, X.; Li, Y.; Guo, X.; Li, Z. Latest assessment methods for mitochondrial homeostasis in cognitive diseases. Neural Regen. Res. 2024, 19, 754–768. [Google Scholar] [CrossRef]
- Chen, N.; Ge, M.-M.; Li, D.-Y.; Wang, X.-M.; Liu, D.-Q.; Ye, D.-W.; Tian, Y.-K.; Zhou, Y.-Q.; Chen, J.-P. β2-adrenoreceptor agonist ameliorates mechanical allodynia in paclitaxel-induced neuropathic pain via induction of mitochondrial biogenesis. Biomed. Pharmacother. 2021, 144, 112331. [Google Scholar] [CrossRef]
- Szczerbinski, L.; Taylor, M.A.; Puchta, U.; Konopka, P.; Paszko, A.; Citko, A.; Szczerbinski, K.; Goscik, J.; Gorska, M.; Larsen, S.; et al. The Response of Mitochondrial Respiration and Quantity in Skeletal Muscle and Adipose Tissue to Exercise in Humans with Prediabetes. Cells 2021, 10, 3013. [Google Scholar] [CrossRef]
- Watson, E.L.; Baker, L.A.; Wilkinson, T.J.; Gould, D.W.; Graham-Brown, M.P.; Major, R.W.; Ashford, R.U.; Philp, A.; Smith, A.C. Reductions in skeletal muscle mitochondrial mass are not restored following exercise training in patients with chronic kidney disease. FASEB J. 2020, 34, 1755–1767. [Google Scholar] [CrossRef]
- Larsen, S.; Nielsen, J.; Hansen, C.N.; Nielsen, L.B.; Wibrand, F.; Stride, N.; Schroder, H.D.; Boushel, R.; Helge, J.W.; Dela, F.; et al. Biomarkers of mitochondrial content in skeletal muscle of healthy young human subjects. J. Physiol. 2012, 590, 3349–3360. [Google Scholar] [CrossRef]
- Granata, C.; Jamnick, N.A.; Bishop, D.J. Training-Induced Changes in Mitochondrial Content and Respiratory Function in Human Skeletal Muscle. Sports Med. 2018, 48, 1809–1828. [Google Scholar] [CrossRef]
- Trewin, A.J.; Berry, B.J.; Wojtovich, A.P. Exercise and Mitochondrial Dynamics: Keeping in Shape with ROS and AMPK. Antioxidants 2018, 7, 7. [Google Scholar] [CrossRef]
- Bo, H.; Zhang, Y.; Ji, L.L. Redefining the role of mitochondria in exercise: A dynamic remodeling. Ann. N. Y. Acad. Sci. 2010, 1201, 121–128. [Google Scholar] [CrossRef]
- Perry, C.G.; Lally, J.; Holloway, G.P.; Heigenhauser, G.J.; Bonen, A.; Spriet, L.L. Repeated transient mRNA bursts precede increases in transcriptional and mitochondrial proteins during training in human skeletal muscle. J. Physiol. 2010, 588, 4795–4810. [Google Scholar] [CrossRef]
- Ding, H.; Jiang, N.; Liu, H.; Liu, X.; Liu, D.; Zhao, F.; Wen, L.; Liu, S.; Ji, L.L.; Zhang, Y. Response of mitochondrial fusion and fission protein gene expression to exercise in rat skeletal muscle. Biochim. Biophys. Acta 2010, 1800, 250–256. [Google Scholar] [CrossRef]
- Cartoni, R.; Léger, B.; Hock, M.B.; Praz, M.; Crettenand, A.; Pich, S.; Ziltener, J.L.; Luthi, F.; Dériaz, O.; Zorzano, A.; et al. Mitofusins 1/2 and ERRalpha expression are increased in human skeletal muscle after physical exercise. J. Physiol. 2005, 567, 349–358. [Google Scholar] [CrossRef]
- Amartuvshin, O.; Lin, C.H.; Hsu, S.C.; Kao, S.H.; Chen, A.; Tang, W.C.; Chou, H.L.; Chang, D.L.; Hsu, Y.Y.; Hsiao, B.S.; et al. Aging shifts mitochondrial dynamics toward fission to promote germline stem cell loss. Aging Cell 2020, 19, e13191. [Google Scholar] [CrossRef]
- Spurlock, B.; Tullet, J.M.A.; Hartman, J.L.; Mitra, K. Interplay of mitochondrial fission-fusion with cell cycle regulation: Possible impacts on stem cell and organismal aging. Exp. Gerontol. 2020, 135, 110919. [Google Scholar] [CrossRef]
- Narendra, D.P.; Jin, S.M.; Tanaka, A.; Suen, D.-F.; Gautier, C.A.; Shen, J.; Cookson, M.R.; Youle, R.J. PINK1 Is Selectively Stabilized on Impaired Mitochondria to Activate Parkin. PLoS Biol. 2010, 8, e1000298. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, W.; Lu, Y.; Zheng, Y.; Pan, L.; Wu, X.; Yuan, Y.; Shen, Z.; Ma, S.; Zhang, X.; et al. BNIP3L/NIX-mediated mitophagy: Molecular mechanisms and implications for human disease. Cell Death Dis. 2021, 13, 14. [Google Scholar] [CrossRef]
- Gao, A.; Jiang, J.; Xie, F.; Chen, L. Bnip3 in mitophagy: Novel insights and potential therapeutic target for diseases of secondary mitochondrial dysfunction. Clin. Chim. Acta 2020, 506, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.M.; Lazarou, M.; Wang, C.; Kane, L.A.; Narendra, D.P.; Youle, R.J. Mitochondrial membrane potential regulates PINK1 import and proteolytic destabilization by PARL. J. Cell Biol. 2010, 191, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhou, X.; Zhu, A.; Xiong, S.; Xie, J.; Bai, Z. Advances in exercise to alleviate sarcopenia in older adults by improving mitochondrial dysfunction. Front. Physiol. 2023, 14, 1196426. [Google Scholar] [CrossRef]
- Fealy, C.E.; Mulya, A.; Lai, N.; Kirwan, J.P. Exercise training decreases activation of the mitochondrial fission protein dynamin-related protein-1 in insulin-resistant human skeletal muscle. J. Appl. Physiol. 2014, 117, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Jamart, C.; Francaux, M.; Millet, G.Y.; Deldicque, L.; Frère, D.; Féasson, L. Modulation of autophagy and ubiquitin-proteasome pathways during ultra-endurance running. J. Appl. Physiol. 2012, 112, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Mauro-Lizcano, M.; Esteban-Martínez, L.; Seco, E.; Serrano-Puebla, A.; Garcia-Ledo, L.; Figueiredo-Pereira, C.; Vieira, H.L.; Boya, P. New method to assess mitophagy flux by flow cytometry. Autophagy 2015, 11, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Lu, A.M.; Wang, Y.; Hong, A.; Chen, Y.; Hu, J.; Li, X.; Qin, Z.H. Chronic resistance training activates autophagy and reduces apoptosis of muscle cells by modulating IGF-1 and its receptors, Akt/mTOR and Akt/FOXO3a signaling in aged rats. Exp. Gerontol. 2013, 48, 427–436. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, B.; Hardy, J.; Plun-Favreau, H. PINK1: From Parkinson’s disease to mitophagy and back again. PLoS Biol. 2023, 21, e3002196. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).