
A Sister Species for the Blue Crab, Callinectes sapidus? A Tale Revealed by Mitochondrial DNA





 , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
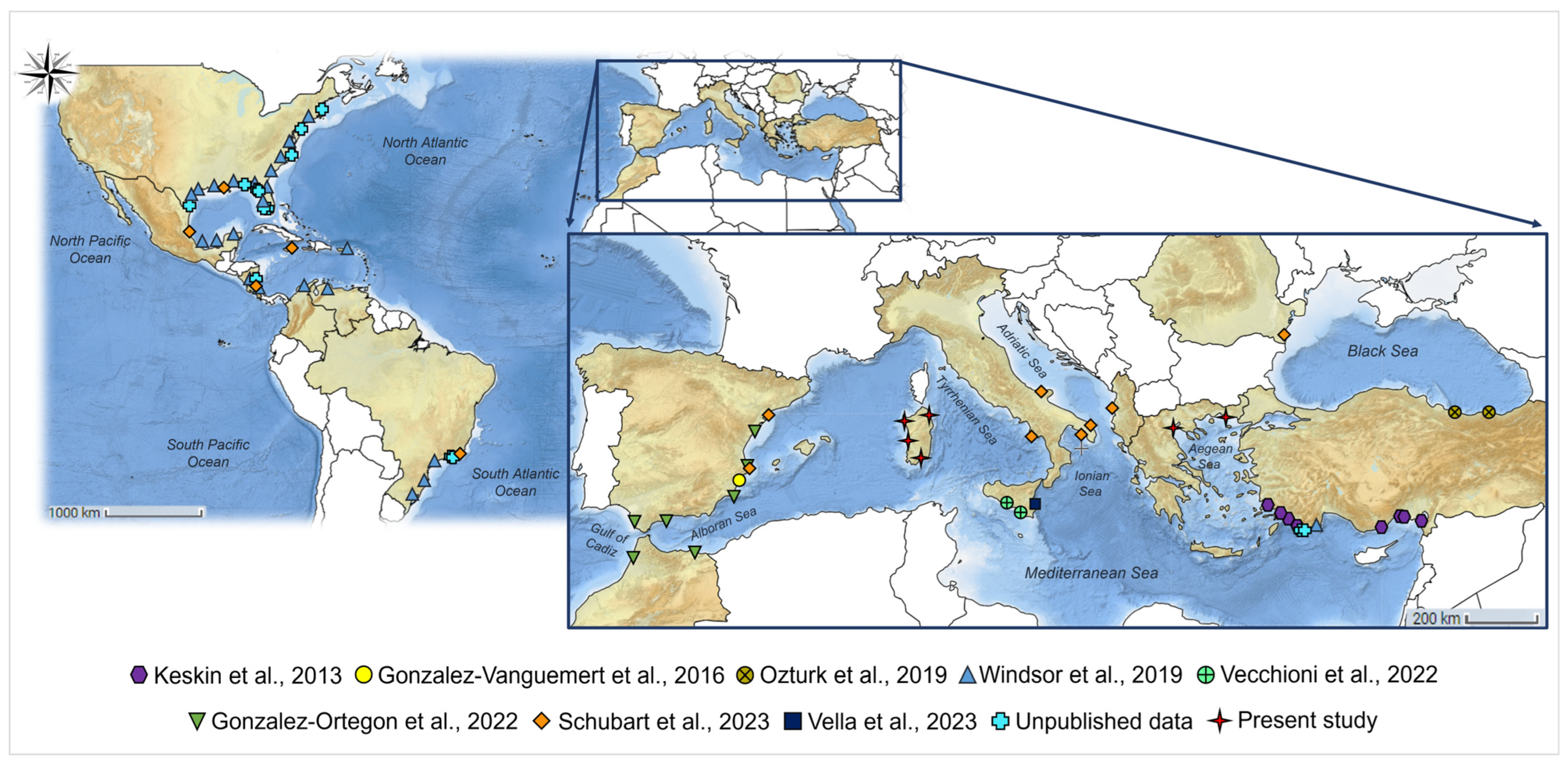
2.1. Sample Collection
2.2. Diagnostic Molecular Analysis
2.3. Phylogeographic and Phylogenetic Analysis
2.4. Estimation of Divergence Time
3. Results
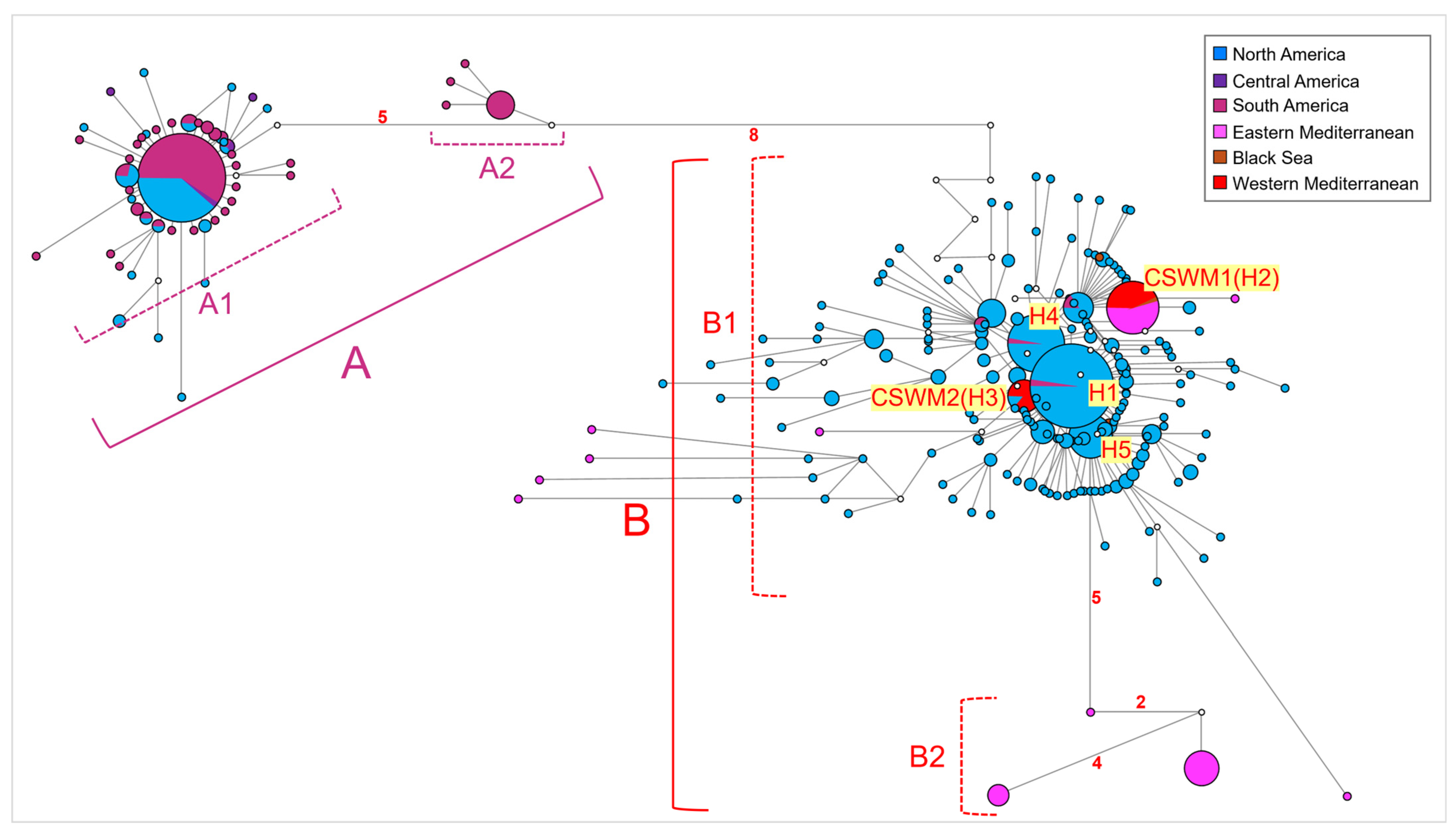
3.1. Phylogeographic and Phylogenetic Relationships among Callinectes sapidus Populations from the Whole Distribution Range
3.2. Evolutionary Relationships among Callinectes sapidus Populations, along with Species of the Genus Callinectes
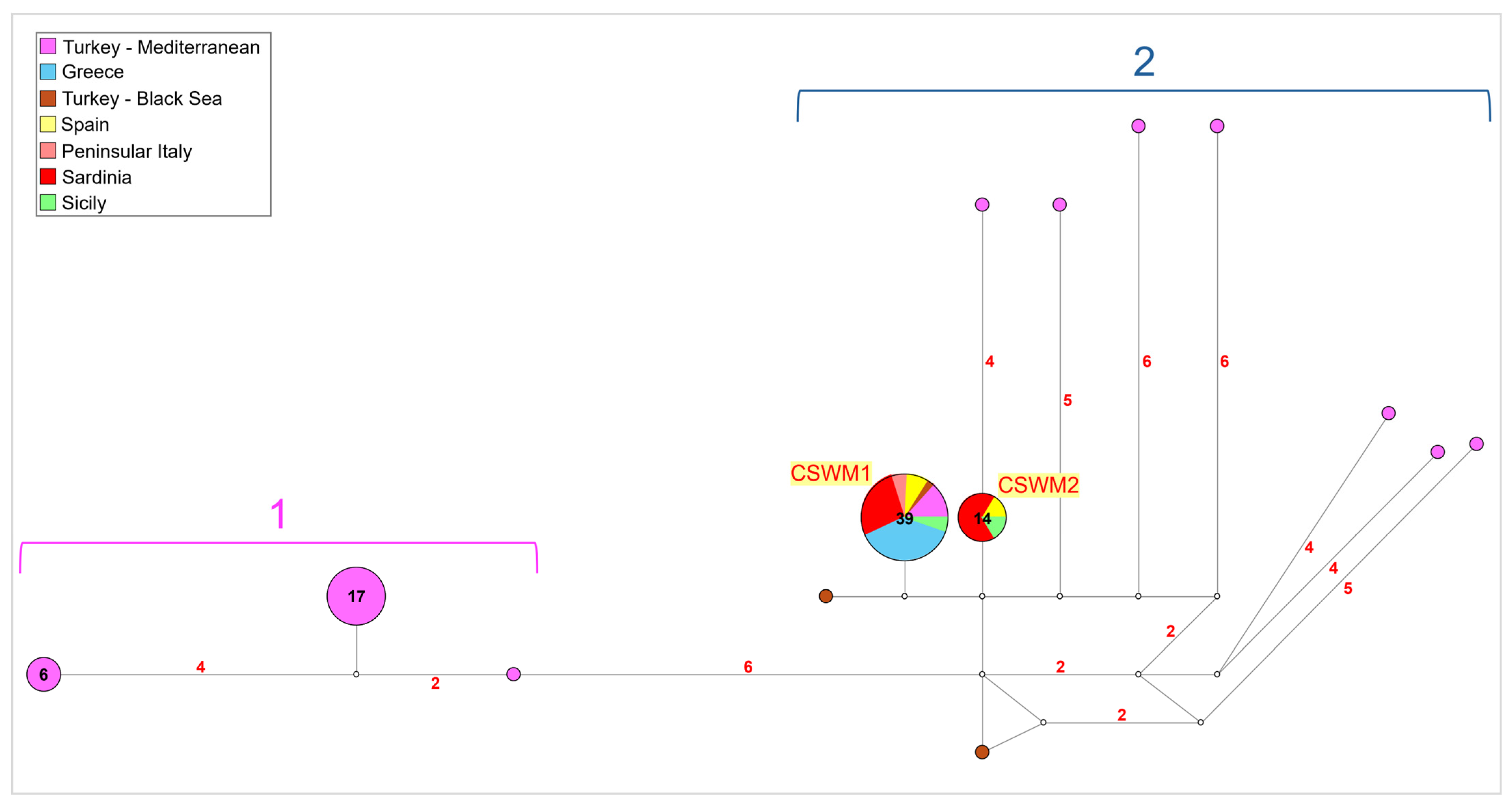
3.3. Genetic Variation of Callinectes sapidus within Its Mediterranean and Black Sea Ranges of Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN; Species Survival Commission. IUCN guidelines for the Prevention of Biodiversity Loss Caused by Alien Invasive Species. Aliens 2000, 11. Available online: https://portals.iucn.org/library/efiles/documents/Rep-2000-052.pdf (accessed on 19 March 2024).
- Streftaris, N.; Zenetos, A. Alien marine species in the Mediterranean-the 100 ‘Worst Invasives’ and their impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef]
- Lucy, F.E.; Caffrey, J.; Dick, J.T.; Davis, E.; Coughlan, N.E. Prevention, Control and Eradication of Invasive Alien Species (No. 368). EPA Research Report. 2021. Available online: https://www.epa.ie/publications/research/biodiversity/Research_Report_368 (accessed on 19 March 2024).
- European Commission; Directorate-General for Environment; Sundseth, K. Invasive Alien Species—A European Union Response. Publications Office, 2016. Available online: https://data.europa.eu/doi/10.2779/374800 (accessed on 19 March 2024).
- Genovesi, P. Guidelines for Eradication of Terrestrial Vertebrates: A European Contribution to the Invasive Alien Species Issue. Other Publications in Wildlife Management. 2000. Available online: https://digitalcommons.unl.edu/icwdmother/24 (accessed on 19 March 2024).
- Mancinelli, G.; Carrozzo, L.; Costantini, M.L.; Rossi, L.; Marini, G.; Pinna, M. Occurrence of the Atlantic blue crab Callinectes sapidus Rathbun, 1896 in two Mediterranean coastal habitats: Temporary visitor or permanent resident? Estuar. Coast. Shelf Sci. 2013, 135, 46–56. [Google Scholar] [CrossRef]
- Zenetos, A.; Çinar, M.E.; Crocetta, F.; Golani, D.; Rosso, A.; Servello, G.; Shenkar, N.; Turon, X.; Verlaque, M. Uncertainties and validation of alien species catalogues: The Mediterranean as an example. Estuar. Coast. Shelf Sci. 2017, 191, 171–187. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Vilà, M.; Hulme, P.E. Impact of Biological Invasions on Ecosystem Services; Springer: Dordrecht, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Mancinelli, G.; Bardelli, R.; Zenetos, A. A global occurrence database of the Atlantic blue crab Callinectes sapidus. Sci. Data 2021, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Hines, A.H.; Haddon, A.M.; Wiechert, L.A. Guild structure and foraging impact of blue crabs and epibenthic fish in a subestuary of Chesapeake Bay. Mar. Ecol. Prog. Ser. 1990, 67, 105–126. [Google Scholar] [CrossRef]
- Laughlin, R.A. Feeding habits of the blue crab, Callinectes sapidus Rathbun, in the Apalachicola estuary, Florida. Bull. Mar. Sci. 1982, 32, 807–822. [Google Scholar]
- Kampouris, T.E.; Porter, J.S.; Sanderson, W.G. Callinectes sapidus Rathbun, 1896 (Brachyura: Portunidae): An assessment on its diet and foraging behaviour, Thermaikos Gulf, NW Aegean Sea, Greece: Evidence for ecological and economic impacts. Crustac. Res. 2019, 48, 23–37. [Google Scholar] [CrossRef]
- Millikin, M.R.; Williams, A.B. Synopsis of biological data on blue crab, Callinectes sapidus Rathbun. In FAO Fisheries Synopsis; NMFS (National Marine Fisheries Service): Silver Spring, MD, USA, 1984; Volume 138, pp. 32–39. [Google Scholar]
- Marchessaux, G.; Chevalier, C.; Mangano, M.C.; Sarà, G. Larval connectivity of the invasive blue crabs Callinectes sapidus and Portunus segnis in the Mediterranean Sea: A step toward improved cross border management. Mar. Pollut. Bull. 2023, 194, 115272. [Google Scholar] [CrossRef]
- Marchessaux, G.; Gjoni, V.; Sarà, G. Environmental drivers of size-based population structure, sexual maturity and fecundity: A study of the invasive blue crab Callinectes sapidus (Rathbun, 1896) in the Mediterranean Sea. PLoS ONE 2023, 18, e0289611. [Google Scholar] [CrossRef] [PubMed]
- Mizerek, T.; Regan, H.M.; Hovel, K.A. Seagrass habitat loss and fragmentation influence management strategies for a blue crab Callinectes sapidus fishery. Mar. Ecol. Prog. Ser. 2011, 427, 247–257. [Google Scholar] [CrossRef]
- Hines, A.H. Ecology of juvenile and adult blue crabs: Summary of discussion of research themes and directions. Bull. Mar. Sci. 2003, 72, 423–433. [Google Scholar]
- Walters, E.A.; Bojko, J.; Crowley, C.E.; Gandy, R.L.; Martin, C.W.; Shea, C.P.; Bateman, K.S.; Stentiford, G.D.; Behringer, D.C. Salinity and temperature affect the symbiont profile and host condition of Florida USA blue crabs Callinectes sapidus. J. Invertebr. Pathol. 2023, 198, 107930. [Google Scholar] [CrossRef] [PubMed]
- Marchessaux, G.; Bosch-Belmar, M.; Cilenti, L.; Lago, N.; Mangano, M.C.; Marsiglia, N.; Sarà, G. The invasive blue crab Callinectes sapidus thermal response: Predicting metabolic suitability maps under future warming Mediterranean scenarios. Front. Mar. Sci. 2022, 9, 1055404. [Google Scholar] [CrossRef]
- Lewis, D.H.; Roer, R.D. Thermal preference in distribution of blue crabs, Callinectes sapidus, in a power plant cooling pond. J. Crustac. Biol. 1988, 8, 283–289. [Google Scholar] [CrossRef]
- García-Rueda, A.; Tremblay, N.; Mascaró, M.; Díaz, F.; Paschke, K.; Caamal-Monsreal, C.; Rosas, C. The thermal tolerance of a tropical population of blue crab (Callinectes sapidus) modulates aerobic metabolism during hypoxia. J. Therm. Biol. 2021, 102, 103078. [Google Scholar] [CrossRef] [PubMed]
- Nehring, S. Invasion history and success of the American blue crab Callinectes sapidus in European and adjacent waters. In The Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts; Galil, B., Clark, P., Carlton, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 607–624. [Google Scholar] [CrossRef]
- Guillory, V.; Perry, H.; Steele, P.; Wagner, T.; Keithly, W.; Pellegrin, B.; Petterson, J.; Floyd, T.; Buckson, B.; Hartman, L.; et al. The Blue Crab Fishery of the Gulf of Mexico, United States: A Regional Management Plan; Guillory, V., Perry, H.M., VanderKooy, S., Eds.; Gulf States Marine Fisheries Commission: Ocean Springs, MS, USA, 2001; Publication 96. Available online: https://www.impactassessment.com/reports/Petterson_2001_Sociocultural_Characterization_Gulf_Mexico_Blue_Crab_Fishery.pdf (accessed on 19 March 2024).
- Churchill, E.P. Life history of the blue crab. Bull. Bur. Fish 1919, 36, 95–128. [Google Scholar]
- Van Engel, W.A. The blue crab and its fishery in Chesapeake Bay. Part 1. Reproduction, early development, growth and migration. Comm. Fish. Rev. 1958, 20, 6. [Google Scholar]
- Hill, J.; Fowler, D.L.; Avyle, M.V. Species Profiles: Life Histories and Environmental Requirements of Coastal Fishes and Invertebrates (Mid-Atlantic)—Blue Crab. U.S. Army Corps of Engineers, Vicksburg, 1989. Available online: https://apps.dtic.mil/sti/pdfs/ADA210181.pdf (accessed on 19 March 2024).
- Kevrekidis, K.; Kevrekidis, T.; Mogias, A.; Boubonari, T.; Kantaridou, F.; Kaisari, N.; Malea, P.; Dounas, C.; Thessalou-Legaki, M. Fisheries Biology and Basic Life-Cycle Characteristics of the Invasive Blue Crab Callinectes sapidus Rathbun in the Estuarine Area of the Evros River (Northeast Aegean Sea, Eastern Mediterranean). J. Mar. Sci. Eng. 2023, 11, 462. [Google Scholar] [CrossRef]
- Gennaio, R.; Scordella, G.; Pastore, M. Occurrence of blue crab Callinectes sapidus (Rathbun, 1986, Crustacea, Brachyura), in the Ugento ponds area (Lecce, Italy). Thalass. Salentina 2006, 29, 29–39. [Google Scholar] [CrossRef]
- Lipcius, R.N.; Eggleston, D.B.; Heck Jr, K.L.; Seitz, R.D.; Van Montfrans, J. Post-settlement abundance, survival, and growth of postlarvae and young juvenile blue crabs in nursery habitats. In The Blue Crab: Callinectes sapidus; Kennedy, V.S., Cronin, L.E., Eds.; Maryland Sea Grant College: College Park, MD, USA, 2007; Volume 13, pp. 535–565. Available online: https://www.researchgate.net/publication/284418968 (accessed on 20 March 2024).
- Hines, A.H. Ecology of Juvenile and Adult Blue Crabs. In The Blue Crab: Callinectes sapidus; Kennedy, V.S., Cronin, L.E., Eds.; Maryland Sea Grant College: College Park, MD, USA, 2007; pp. 565–654. Available online: https://repository.si.edu/bitstream/handle/10088/18447/serc_Hines_2007_Chapter_14_565_654.pdf (accessed on 20 March 2024).
- Epifanio, C.E. Early life history of the blue crab Callinectes sapidus: A review. J. Shellfish Res. 2019, 38, 1–22. [Google Scholar] [CrossRef]
- Taylor, D.L.; Fehon, M.M. Blue crab (Callinectes sapidus) population structure in southern New England tidal rivers: Patterns of shallow water, unvegetated habitat use and quality. Estuar. Coast. 2021, 44, 1320–1343. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, E.L. Sur un Callinectes sapidus M. Rathbun trouvé à Rochefort. Bull. Mus. Hist. Nat. Paris 1901, 7, 16–17. [Google Scholar]
- Ribeiro, F.; Veríssimo, A. A new record of Callinectes sapidus in a western European estuary (Portuguese coast). Mar. Biodivers. Rec. 2014, 7, e36. [Google Scholar] [CrossRef]
- Eldredge, L.G. First record of the blue crab (Callinectes sapidus) in Hawaii (Decapoda: Brachyura). Bishop Mus. Occas. Pap. 1995, 42, 55–58. [Google Scholar]
- Monin, V. New find of the blue crab Callinectes sapidus (Decapoda, Brachyura) in the Black Sea. Zool. Zh. 1984, 63, 1100–1101. [Google Scholar]
- Labrune, C.; Amilhat, E.; Amouroux, J.; Jabouin, C.; Gigou, A.; Noël, P. The arrival of the American blue crab, Callinectes sapidus Rathbun, 1896 (Decapoda: Brachyura: Portunidae), in the Gulf of Lions (Mediterranean Sea). BioInvasions Rec. 2019, 8, 876–881. [Google Scholar] [CrossRef]
- Mancinelli, G.; Chainho, P.; Cilenti, L.; Falco, S.; Kapiris, K.; Katselis, G.; Ribeiro, F. The Atlantic blue crab Callinectes sapidus in southern European coastal waters: Distribution, impact and prospective invasion management strategies. Mar. Pollut. Bull. 2017, 119, 5–11. [Google Scholar] [CrossRef]
- Cerri, J.; Chiesa, S.; Bolognini, L.; Mancinelli, G.; Grati, F.; Dragičević, B.; Dulčic, J.; Azzurro, E. Using online questionnaires to assess marine bio-invasions: A demonstration with recreational fishers and the Atlantic blue crab Callinectes sapidus (Rathbun, 1986) along three Mediterranean countries. Mar. Pollut. Bull. 2020, 156, 111209. [Google Scholar] [CrossRef]
- Serbetis, C. Un nouveau Crustacé comestible en Mer Egée Callinectes sapidus Rathb. (Décapode brachyoure). Proc. Gen. Fish. Comm. Mediterr. 1959, 5, 505–507. [Google Scholar]
- Georgiadis, C.; Georgiadis, G. Zur Kenntnis der Crustacea Decapoda des Golfes von Thessaloniki. Crustaceana 1974, 26, 239–248. [Google Scholar] [CrossRef]
- Giordani Soika, A. Il Neptunus pelagicus (L.) nell’Alto Adriatico. Natura 1951, 42, 18–20. [Google Scholar]
- Mizzan, L. Presence of swimming crabs of the genus Callinectes (Stimpson) (Decapoda, Portunidae) in the Venice Lagoon (North Adriatic Sea-Italy): First record of Callinectes danae Smith in European waters. Boll. Mus. Civ. Stor. Nat. Venezia 1993, 42, 31–43. [Google Scholar]
- Holthuis, L.B.; Gottlieb, L. The occurrence of the American blue crab, Callinectes sapidus Rathbun, in Israel waters. Bull. Res. Counc. Isr. B 1955, 5, 154–156. [Google Scholar]
- Shiber, J.G. Brachyurans from Lebanese waters. Bull. Mar. Sci. 1981, 31, 864–875. [Google Scholar]
- Abdel-Razek, F.A. Crab fishery of the Egyptian waters with notes on the bionomics of Portunus pelagicus (L.). Acta Adriat. 1987, 28, 143–154. [Google Scholar]
- Tortonese, E. La comparsa di Callinectes sapidus Rath. (Decapoda, Brachyura) nel Mar Ligure. Ann. Mus. Civ. Stor. Nat. Genova 1965, 4, 1–3. [Google Scholar]
- Suaria, G.; Pierucci, A.; Zanello, P.; Fanelli, E.; Chiesa, S.; Azzurro, E. Percnon gibbesi (H. Milne Edwards, 1853) and Callinectes sapidus (Rathbun, 1896) in the Ligurian Sea: Two additional invasive species detections made in collaboration with local fishermen. BioInvasions Rec. 2017, 6, 147–151. [Google Scholar] [CrossRef]
- Cavaliere, A.; Berdar, A. Presenza di Callinectes sapidus Rathbun (Decapoda, Brachyura) nello Stretto di Messina. Boll. Pesca Piscicolt. Idrobiol. 1975, 30, 315–322. [Google Scholar]
- Bisconti, M.; Silvi, E. Prima segnalazione di Callinectes sapidus Rathbun, 1896 (Crustacea, Decapoda, Brachyura) nella provincia di Livorno. Quad. Mus. Stor. Nat. Livorno 2005, 18, 1–6. [Google Scholar]
- Stasolla, G.; Innocenti, G. New records of the invasive crabs Callinectes sapidus Rathbun, 1896 and Percnon gibbesi (H. Milne Edwards, 1853) along the Italian coasts. BioInvasions Rec. 2014, 3, 39–43. [Google Scholar] [CrossRef]
- Carrozzo, L.; Potenza, L.; Carlino, P.; Costantini, M.L.; Rossi, L.; Mancinelli, G. Seasonal abundance and trophic position of the Atlantic blue crab Callinectes sapidus Rathbun 1896 in a Mediterranean coastal habitat. Rend. Lincei 2014, 25, 201–208. [Google Scholar] [CrossRef]
- Florio, M.; Breber, P.; Scirocco, T.; Specchiulli, A.; Cilenti, L.; Lumare, L. Exotic species in Lesina and Varano lakes new guest in Lesina and Varano lakes: Gargano National Park (Italy). Transit. Waters Bull. 2008, 2, 69–79. [Google Scholar] [CrossRef]
- Mancinelli, G.; Glamuzina, B.; Petrić, M.; Carrozzo, L.; Glamuzina, L.; Zotti, M.; Raho, D.; Vizzini, S. The trophic position of the Atlantic blue crab Callinectes sapidus Rathbun 1896 in the food web of Parila lagoon (South Eastern Adriatic, Croatia): A first assessment using stable isotopes. Mediterr. Mar. Sci. 2016, 17, 634–643. [Google Scholar] [CrossRef]
- Cilenti, L.; Pazienza, G.; Scirocco, T.; Fabbrocini, A.; D’Adamo, R. First record of ovigerous Callinectes sapidus (Rathbun, 1896) in the Gargano lagoons (south-west Adriatic Sea). BioInvasions Rec. 2015, 4, 281–287. [Google Scholar] [CrossRef]
- Manfrin, C.; Turolla, E.; Chung, J.S.; Giulianini, P.G. First occurrence of Callinectes sapidus (Rathbun, 1896) within the Sacca di Goro (Italy) and surroundings. Check List 2015, 11, 1640. [Google Scholar] [CrossRef]
- Manfrin, C.; Comisso, G.; dall’Asta, A.; Bettoso, N.; Chung, J.S. The return of the Blue Crab, Callinectes sapidus Rathbun, 1896, after 70 years from its first appearance in the Gulf of Trieste, northern Adriatic Sea, Italy (Decapoda: Portunidae). Check List 2016, 12, 2006. [Google Scholar] [CrossRef]
- Mordenti, O.; Scaravelli, D. Segnalazioni Faunistiche n° 83–87 (Callinectes sapidus). Quad. Studi Notiz. Stor. Nat. Romagna 2007, 24, 155–156. [Google Scholar]
- Giansante, C. Segnalazioni faunistiche n. 119–124. Quad. Studi Notiz. Stor. Nat. Romagna 2012, 36, 207–208. [Google Scholar]
- Castriota, L.; Andaloro, F.; Costantini, R.; De Ascentiis, A. First record of the Atlantic crab Callinectes sapidus Rathbun, 1896 (Crustacea: Brachyura: Portunidae) in Abruzzi waters, central Adriatic Sea. Acta Adriat. 2012, 53, 467–470. [Google Scholar]
- Castejón, D.; Guerao, G. A new record of the American blue crab, Callinectes sapidus Rathbun, 1896 (Decapoda: Brachyura: Portunidae), from the Mediterranean coast of the Iberian Peninsula. BioInvasions Rec. 2013, 2, 141–143. [Google Scholar] [CrossRef]
- El Zrelli, R.; Rabaoui, L.; Ragkousis, M.; Abdelali, N.; Azzurro, E.; Badreddine, A.; Bariche, M.; Bitar, G.; Crocetta, F.; Denitto, F.; et al. First record of the Atlantic blue crab Callinectes sapidus Rathbun, 1896 in the Gulf of Gabès (south-eastern Tunisia). New Alien Mediterranean Biodiversity Records (October 2020). Medit. Mar. Sci. 2020, 21, 631–652. [Google Scholar] [CrossRef]
- Culurgioni, J.; Diciotti, R.; Satta, C.T.; Camedda, A.; de Lucia, G.A.; Pulina, S.; Lugliè, A.; Brundu, R.; Fois, N. First data on the appearance of Callinectes sapidus (Rathbun, 1896) in Sardinia (western Mediterranean). In Proceedings of the Atti del XXVIII Congresso Società Italiana di Ecologia, Cagliari, Italy, 12–14 September 2018; p. 112. [Google Scholar]
- Piras, P.; Esposito, G.; Meloni, D. On the occurrence of the blue crab Callinectes sapidus (Rathbun, 1896) in Sardinian coastal habitats (Italy): A present threat or a future resource for the regional fishery sector? BioInvasions Rec. 2019, 8, 134–141. [Google Scholar] [CrossRef]
- Culurgioni, J.; Diciotti, R.; Satta, C.T.; Camedda, A.; de Lucia, G.A.; Pulina, S.; Lugliè, A.; Brundu, R.; Fois, N. Distribution of the alien species Callinectes sapidus (Rathbun, 1896) in Sardinian waters (western Mediterranean). BioInvasions Rec. 2020, 9, 65–73. [Google Scholar] [CrossRef]
- Öztürk, R.Ç.; Terzi, Y.; Feyzioğlu, A.M.; Şahin, A.; Aydın, M. Genetic characterization of the invasive Blue crab, Callinectes sapidus (Rathbun, 1896), in the Black Sea. Reg. Stud. Mar. Sci. 2020, 39, 101412. [Google Scholar] [CrossRef]
- González-Ortegón, E.; Berger, S.; Encarnação, J.; Chairi, H.; Morais, P.; Teodosio, M.A.; Oliva-Paterna, F.J.; Schubart, C.D.; Cuesta, J.A. Free pass through the pillars of Hercules? Genetic and historical insights into the recent expansion of the Atlantic blue crab Callinectes sapidus to the West and the East of the Strait of Gibraltar. Front. Mar. Sci. 2022, 9, 918026. [Google Scholar] [CrossRef]
- Schubart, C.D.; Deli, T.; Mancinelli, G.; Cilenti, L.; Gil Fernández, A.; Falco, S.; Berger, S. Phylogeography of the Atlantic blue crab Callinectes sapidus (Brachyura: Portunidae) in the Americas versus the Mediterranean Sea: Determining origins and genetic connectivity of a large-scale invasion. Biology 2022, 12, 35. [Google Scholar] [CrossRef]
- Bilen, C.T.; Yesilyurt, I.N. Growth of blue crab, Callinectes sapidus, in the Yumurtalik Cove, Turkey: A molt process approach. Cent. Eur. J. Biol. 2014, 9, 49–57. [Google Scholar] [CrossRef]
- Öndes, F.; Gökçe, G. Distribution and Fishery of the Blue Crab (Callinectes sapidus Rathbun, 1896) in Turkey Based on Local Ecological Knowledge of Fishers. J. Anatol. Environ. Animal Sci. 2021, 6, 325–332. [Google Scholar] [CrossRef]
- Lattos, A.; Papadopoulos, D.K.; Giantsis, I.A.; Stamelos, A.; Karagiannis, D. Histopathology and Phylogeny of the Dinoflagellate Hematodinium perezi and the Epibiotic Peritrich Ciliate Epistylis sp. Infecting the Blue Crab Callinectes sapidus in the Eastern Mediterranean. Microorganisms 2024, 12, 456. [Google Scholar] [CrossRef]
- Wolff, T. Occurrence of Two East American Species of Crabs in European Waters. Nature 1954, 174, 188–189. [Google Scholar] [CrossRef]
- ICES WGITMO. Report of the Working Group on Introductions and Transfers of Marine Organisms. In Proceedings of the ICES CM 2006/ACME:05, Oostende, Belgium, 16–17 March 2006; p. 334. [Google Scholar]
- Small, H.J.; Shields, J.D.; Hudson, K.L.; Reece, K.S. Molecular detection of Hematodinium sp. infecting the blue crab, Callinectes sapidus. J. Shellfish Res. 2007, 26, 131–139. [Google Scholar] [CrossRef]
- Lohan, K.M.P. Molecular Genetic Studies of Hematodinium sp., a Parasitic Dinoflagellate of the Blue Crab, Callinectes sapidus; The College of William & Mary, Virginia Institute of Marine Science: Gloucester Point, VA, USA, 2011. [Google Scholar] [CrossRef]
- Pagliara, P.; Mancinelli, G. Parasites affect hemocyte functionality in the hemolymph of the invasive Atlantic blue crab Callinectes sapidus from a coastal habitat of the Salento Peninsula (SE Italy). Mediterr. Mar. Sci. 2018, 19, 193–200. [Google Scholar] [CrossRef]
- Small, H.J.; Huchin-Mian, J.P.; Reece, K.S.; Pagenkopp Lohan, K.M.; Butler, M.J., IV; Shields, J.D. Parasitic dinoflagellate Hematodinium perezi prevalence in larval and juvenile blue crabs Callinectes sapidus from coastal bays of Virginia. Dis. Aquat. Org. 2019, 134, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Flowers, E.M.; Johnson, A.F.; Aguilar, R.; Schott, E.J. Prevalence of the pathogenic crustacean virus Callinectes sapidus reovirus 1 near flow-through blue crab aquaculture in Chesapeake Bay, USA. Dis. Aquat. Org. 2018, 129, 135–144. [Google Scholar] [CrossRef]
- Lively, J.A.; Spitznagel, M.I.; Schott, E.J.; Small, H.J. Investigating conspecific CsRV1 transmission in Callinectes sapidus. J. Invertebr. Pathol. 2023, 201, 107987. [Google Scholar] [CrossRef]
- Zhao, M.; Plough, L.V.; Behringer, D.C.; Bojko, J.; Kough, A.S.; Alper, N.W.; Xu, L.; Schott, E.J. Cross-hemispheric genetic diversity and spatial genetic structure of Callinectes sapidus reovirus 1 (CsRV1). Viruses 2023, 15, 563. [Google Scholar] [CrossRef]
- McMillen-Jackson, A.L.; Bert, T.M.; Steele, P. Population genetics of the blue crab Callinectes sapidus: Modest population structuring in a background of high gene flow. Mar. Biol. 1994, 118, 53–65. [Google Scholar] [CrossRef]
- Place, A.R.; Feng, X.; Steven, C.R.; Fourcade, H.M.; Boore, J.L. Genetic markers in blue crabs (Callinectes sapidus): II. Complete mitochondrial genome sequence and characterization of genetic variation. J. Exp. Mar. Biol. Ecol. 2005, 319, 15–27. [Google Scholar] [CrossRef]
- Yednock, B.K.; Neigel, J.E. An investigation of genetic population structure in blue crabs, Callinectes sapidus, using nuclear gene sequences. Mar. Biol. 2014, 161, 871–886. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Dumont, L.F.C.; dos Santos, C.R.M.; D’Incao, F.; Weiss, S.; Froufe, E. Two distinct mtDNA lineages of the blue crab reveal large-scale population structure in its native Atlantic distribution. Estuar. Coast. Shelf Sci. 2017, 197, 45–53. [Google Scholar] [CrossRef]
- Windsor, A.M.; Moore, M.K.; Warner, K.A.; Stadig, S.R.; Deeds, J.R. Evaluation of variation within the barcode region of Cytochrome c Oxidase I (COI) for the detection of commercial Callinectes sapidus Rathbun, 1896 (blue crab) products of non-US origin. PeerJ 2019, 7, e7827. [Google Scholar] [CrossRef]
- Vecchioni, L.; Russotto, S.; Arculeo, M.; Marrone, F. On the occurrence of the invasive Atlantic blue crab Callinectes sapidus Rathbun 1896 (Decapoda: Brachyura: Portunidae) in Sicilian inland waters. Nat. Hist. Sci. 2022, 9, 43–46. [Google Scholar] [CrossRef]
- Vella, A.; Giarrusso, E.; Monaco, C.; Misfud, C.M.; Darmanin, S.A.; Raffa, A.; Tumino, C.; Peri, I.; Vella, N. New Records of Callinectes sapidus (Crustacea, Portunidae) from Malta and the San Leonardo River Estuary in Sicily (Central Mediterranean). Diversity 2023, 15, 679. [Google Scholar] [CrossRef]
- Keskin, E.; Atar, H.H. DNA barcoding commercially important aquatic invertebrates of Turkey. Mitochondrial DNA 2013, 24, 440–450. [Google Scholar] [CrossRef]
- Gonzalez-Wanguemert, M.; Pujol, J.A. First record of the atlantic blue crab Callinectes sapidus (Crustacea: Brachyura: Portunidae) in the Segura river mouth (Spain, southwestern Mediterranean Sea). Turk. J. Zool. 2016, 40, 615–619. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Sievers, F.; Higgins, D.G. Clustal Omega, Accurate Alignment of Very Large Numbers of Sequences. In Multiple Sequence Alignment Methods, Russell, D., Ed.; Humana Press: Totowa, NJ, USA, 2014; Volume 1079, pp. 105–116. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; Ugene Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.A.; Strimmer, K.; Vingron, M.; Von Haeseler, A. TREE-PUZZLE: Maximum likelihood phylogenetic analysis using quartets and parallel computing. Bioinformatics 2002, 18, 502–504. [Google Scholar] [CrossRef]
- Scarpa, F.; Cossu, P.; Sanna, D.; Lai, T.; Casu, M.; Curini-Galletti, M. New insights on the genus Otoplana Du Plessis, 1889 (Platyhelminthes: Proseriata), with description of two new species from the Canary Islands. Mar. Biodivers. 2019, 49, 2075–2087. [Google Scholar] [CrossRef]
- Scarpa, F.; Sanna, D.; Cossu, P.; Lai, T.; Casu, M.; Curini-Galletti, M. How to achieve internal fertilization without a vagina: The study case of the genus Archilina Ax, 1959 (Platyhelminthes, Proseriata) from Canary Islands. Mar. Biodivers. 2019, 49, 2057–2073. [Google Scholar] [CrossRef]
- Strimmer, K.; von Haeseler, A. Quartet Puzzling: A quartet Maximum-Likelihood method for reconstructing tree topologies. Mol. Biol. Evol. 1996, 13, 964–969. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Scarpa, F.; Cossu, P.; Sanna, D.; Lai, T.; Norenburg, J.L.; Curini-Galletti, M.; Casu, M. An 18S and 28S-based clock calibration for marine Proseriata (Platyhelminthes). J. Exp. Mar. Biol. Ecol. 2015, 463, 22–31. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2020, 21, 609–620. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Scarpa, F.; Cossu, P.; Lai, T.; Sanna, D.; Curini-Galletti, M.; Casu, M. Meiofaunal cryptic species challenge species delimitation: The case of the Monocelis lineata (Platyhelminthes: Proseriata) species complex. Contrib. Zool. 2016, 85, 123–145. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; Dewaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, F.; Cossu, P.; Delogu, V.; Lai, T.; Sanna, D.; Leasi, F.; Norenuburg, J.L.; Curini-Galletti, M.; Casu, M. Molecular support for morphology-based family-rank taxa: The contrasting cases of two families of Proseriata (Platyhelminthes). Zool. Scr. 2017, 46, 753–766. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, e214. [Google Scholar] [CrossRef]
- Schubart, C.D.; Diesel, R.; Hedges, S.B. Rapid evolution to terrestrial life in Jamaican crabs. Nature 1998, 393, 363–365. [Google Scholar] [CrossRef]
- Daniels, S.R.; Phiri, E.E.; Klaus, S.; Albrecht, C.; Cumberlidge, N. Multilocus phylogeny of the Afrotropical freshwater crab fauna reveals historical drainage connectivity and transoceanic dispersal since the Eocene. Syst. Biol. 2015, 64, 549–567. [Google Scholar] [CrossRef]
- Rambaut, A.; Suchard, M.A.; Xie, D. Drummond. Tracer v1.6. A.J. 2014. Available online: http://tree.bio.ed.ac.uk/software/tracer/ (accessed on 15 June 2024).
- Strimmer, K.; von Haeseler, A. Likelihood-mapping: A simple method to visualize phylogenetic content of a sequence alignment. Proc. Natl. Acad. Sci. USA 1997, 94, 6815–6819. [Google Scholar] [CrossRef]
- Head, M.J.; Gibbard, P.L. Early–Middle Pleistocene transitions: Linking terrestrial and marine realms. Quat. Int. 2015, 389, 7–46. [Google Scholar] [CrossRef]
- Oliveira, D.; Desprat, S.; Yin, Q.; Rodrigues, T.; Naughton, F.; Trigo, R.; Su, Q.; Grimalt, J.O.; Alonso-Garcia, M.; Voelker, A.H.L.; et al. Enhanced humidity in SW Iberia driven by the combination of insolation and ice-sheet forcing during MIS 13 interglacial. In Proceedings of the EGU General Assembly Conference Abstracts, EGU General Assembly 2021, Online, 19–30 April 2021; p. EGU21-11408. [Google Scholar] [CrossRef]
- Ludt, W.B.; Rocha, L.A. Shifting seas: The impacts of Pleistocene sea-level fluctuations on the evolution of tropical marine taxa. J. Biogeogr. 2015, 42, 25–38. [Google Scholar] [CrossRef]
- Lacerda, A.L.F.; Kersanach, R.; Cortinhas, M.C.S.; Prata, P.F.S.; Dumont, L.F.C.; Proietti, M.C.; Maggioni, R.; D’Incao, F. High connectivity among blue crab (Callinectes sapidus) populations in the Western South Atlantic. PLoS ONE 2016, 11, e0153124. [Google Scholar] [CrossRef] [PubMed]
- Olival, K.J.; Latinne, A.; Islam, A.; Epstein, J.H.; Hersch, R.; Engstrand, R.C.; Gurley, E.S.; Amato, G.; Luby, S.P.; Daszak, P. Population genetics of fruit bat reservoir informs the dynamics, distribution and diversity of Nipah virus. Mol. Ecol. 2020, 29, 970–985. [Google Scholar] [CrossRef] [PubMed]
- Akdaş, O.; Ayaz, İ.S.; Çaylan, D.Ö. Analysing the trade between Turkey and the Aegean Islands: The hidden opportunities. Dokuz Eylül Üniv. Denizcilik Fak. Derg. 2018, 10, 109–126. [Google Scholar] [CrossRef]
- Cowen, R.K.; Sponaugle, S. Larval dispersal and marine population connectivity. Ann. Rev. Mar. Sci. 2009, 1, 443–466. [Google Scholar] [CrossRef] [PubMed]
- González-Wangüemert, M.; Basso, L.; Balau, A.; Costa, J.; Renault, L.; Serrão, E.A.; Duarte, C.M.; Hendriks, I.E. Gene pool and connectivity patterns of Pinna nobilis in the Balearic Islands (Spain, Western Mediterranean Sea): Implications for its conservation through restocking. Aquatic Conserv. Mar. Freshw. Ecosyst. 2019, 29, 175–188. [Google Scholar] [CrossRef]
- Willis, J.H.; Orr, H.A. Increased heritable variation following population bottlenecks: The role of dominance. Evolution 1993, 47, 949–957. [Google Scholar] [CrossRef]
- Sanna, D.; Scarpa, F.; Lai, T.; Cossu, P.; Falautano, M.; Castriota, L.; Andaloro, F.; Follesa, M.C.; Francalacci, P.; Curini-Galletti, M.; et al. Fistularia commersonii (Teleostea: Fistulariidae): Walking through the Lessepsian paradox of mitochondrial DNA. Ital. J. Zool. 2015, 82, 499–512. [Google Scholar] [CrossRef]
- Sanna, D.; Azzena, I.; Scarpa, F.; Cossu, P.; Pira, A.; Gagliardi, F.; Casu, M. First Record of the Alien Species Procambarus virginalis Lyko, 2017 in FreshWaters of Sardinia and Insight into Its Genetic Variability. Life 2021, 11, 606. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographic Region | N | S | H | h | π |
|---|---|---|---|---|---|
| United States of America | 397 | 100 | 146 | 0.922 | 0.00766 |
| Puerto Rico | 22 | 34 | 10 | 0.749 | 0.00600 |
| Jamaica | 1 | 0 | 0 | 0.000 | 0.00000 |
| United Mexican States | 43 | 38 | 22 | 0.844 | 0.01777 |
| North America | 463 | 108 | 162 | 0.931 | 0.01249 |
| Nicaragua | 4 | 2 | 3 | 0.833 | 0.00184 |
| Costa Rica | 1 | 0 | 0 | 0.000 | 0.00000 |
| Central America | 5 | 4 | 4 | 0.900 | 0.00284 |
| Venezuela | 90 | 45 | 30 | 0.614 | 0.00216 |
| Colombia | 4 | 2 | 3 | 0.833 | 0.00158 |
| Brazil | 19 | 22 | 7 | 0.667 | 0.01168 |
| South America | 113 | 53 | 39 | 0.729 | 0.00712 |
| Turkey—Black Sea | 3 | 5 | 3 | 1.000 | 0.00527 |
| Greece | 15 | 0 | 1 | 0.000 | 0.00000 |
| Turkey—Levantine Sea | 36 | 46 | 11 | 0.744 | 0.01317 |
| Eastern Mediterranean | 51 | 45 | 11 | 0.733 | 0.01289 |
| Spain | 5 | 3 | 2 | 0.600 | 0.00292 |
| Peninsular Italy | 2 | 0 | 1 | 0.000 | 0.00000 |
| Sardinia | 21 | 3 | 2 | 0.524 | 0.00248 |
| Sicily | 4 | 3 | 2 | 0.500 | 0.00237 |
| Western Mediterranean | 32 | 3 | 2 | 0.508 | 0.00247 |
| Whole Mediterranean | 83 | 45 | 12 | 0.725 | 0.01103 |
| Whole dataset | 667 | 124 | 196 | 0.937 | 0.01793 |
| Group | N | S | H | h | π |
|---|---|---|---|---|---|
| A | 176 | 60 | 50 | 0.685 | 0.00349 |
| B | 491 | 110 | 156 | 0.933 | 0.00699 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Locci, C.; Azzena, I.; Pascale, N.; Ciccozzi, A.; Deplano, I.; Giantsis, I.A.; Papadopoulos, D.K.; Lattos, A.; Orrù, F.; Puzzi, C.M.; et al. A Sister Species for the Blue Crab, Callinectes sapidus? A Tale Revealed by Mitochondrial DNA. Life 2024, 14, 1116. https://doi.org/10.3390/life14091116
Locci C, Azzena I, Pascale N, Ciccozzi A, Deplano I, Giantsis IA, Papadopoulos DK, Lattos A, Orrù F, Puzzi CM, et al. A Sister Species for the Blue Crab, Callinectes sapidus? A Tale Revealed by Mitochondrial DNA. Life. 2024; 14(9):1116. https://doi.org/10.3390/life14091116
Chicago/Turabian StyleLocci, Chiara, Ilenia Azzena, Noemi Pascale, Alessandra Ciccozzi, Ilaria Deplano, Ioannis A. Giantsis, Dimitrios K. Papadopoulos, Athanasios Lattos, Flavio Orrù, Cesare M. Puzzi, and et al. 2024. "A Sister Species for the Blue Crab, Callinectes sapidus? A Tale Revealed by Mitochondrial DNA" Life 14, no. 9: 1116. https://doi.org/10.3390/life14091116
APA StyleLocci, C., Azzena, I., Pascale, N., Ciccozzi, A., Deplano, I., Giantsis, I. A., Papadopoulos, D. K., Lattos, A., Orrù, F., Puzzi, C. M., Scarpa, F., Casu, M., & Sanna, D. (2024). A Sister Species for the Blue Crab, Callinectes sapidus? A Tale Revealed by Mitochondrial DNA. Life, 14(9), 1116. https://doi.org/10.3390/life14091116