
From Nature to Treatment: The Impact of Pterostilbene on Mitigating Retinal Ischemia–Reperfusion Damage by Reducing Oxidative Stress, Inflammation, and Apoptosis


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
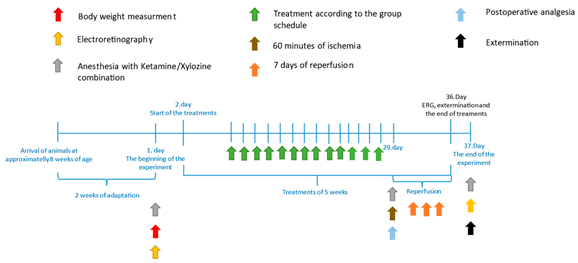
2.1. Animals and Groups
2.2. Ocular Ischemia–Reperfusion
2.3. Electroretinography
2.4. Western Blot
2.5. Histology
2.6. Statistical Analyses
3. Results
3.1. Electroretinography
3.2. Western Blot
3.3. Histology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinskey, N.D.; Besirli, C.G.; Zacks, D.N. Retinal cell death and current strategies in retinal neuroprotection. Curr. Opin. Ophthalmol. 2014, 25, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.N.; Casson, R.J.; Wood, J.P.; Chidlow, G.; Graham, M.; Melena, J. Retinal ischemia: Mechanisms of damage and potential therapeutic strategies. Prog. Retin. Eye Res. 2004, 23, 91–147. [Google Scholar] [CrossRef] [PubMed]
- Bresnick, G.H.; De Venecia, G.; Myers, F.L.; Harris, J.A.; Davis, M.D. Retinal ischemia in diabetic retinopathy. Arch. Ophthalmol. 1975, 93, 1300–1310. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; An, Q.; Ji, H.; Dai, J.; Suo, L.; Zhang, C. Retinal ischemia-reperfusion injury induces intense lipid synthesis and remodeling. Biochem. Biophys. Res. Commun. 2023, 689, 149232. [Google Scholar] [CrossRef]
- Qin, Q.; Yu, N.; Gu, Y.; Ke, W.; Zhang, Q.; Liu, X.; Wang, K.; Chen, M. Inhibiting multiple forms of cell death optimizes ganglion cells survival after retinal ischemia reperfusion injury. Cell Death Dis. 2022, 13, 507. [Google Scholar] [CrossRef]
- Jurcau, A.; Ardelean, I.A. Molecular pathophysiological mechanisms of ischemia/reperfusion injuries after recanalization therapy for acute ischemic stroke. J. Integr. Neurosci. 2021, 20, 727–744. [Google Scholar] [CrossRef]
- Li, R.; Jia, Z.; Trush, M.A. Defining ROS in Biology and Medicine. React. Oxyg. Species 2016, 1, 9–21. [Google Scholar] [CrossRef]
- Chandrasekaran, A.; Idelchik, M.; Melendez, J.A. Redox control of senescence and age-related disease. Redox Biol. 2017, 11, 91–102. [Google Scholar] [CrossRef]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39, 73–82. [Google Scholar] [CrossRef]
- Carrera-Juliá, S.; Moreno, M.L.; Barrios, C.; de la Rubia Ortí, J.E.; Drehmer, E. Antioxidant Alternatives in the Treatment of Amyotrophic Lateral Sclerosis: A Comprehensive Review. Front. Physiol. 2020, 11, 63. [Google Scholar] [CrossRef]
- Abtahi, S.H.; Nourinia, R.; Mazloumi, M.; Nouri, H.; Arevalo, J.F.; Ahmadieh, H. Retinal ischemic cascade: New insights into the pathophysiology and imaging findings. Surv. Ophthalmol. 2023, 68, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Han, Y.; Lee, J.E.; Yenari, M.A. The 70-kDa heat shock protein (Hsp70) as a therapeutic target for stroke. Expert. Opin. Ther. Targets 2018, 22, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Jurga, A.M.; Paleczna, M.; Kadluczka, J.; Kuter, K.Z. Beyond the GFAP-Astrocyte Protein Markers in the Brain. Biomolecules 2021, 11, 1361. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, S.H.; Pierce, E.A. Parthanatos as a Cell Death Pathway Underlying Retinal Disease. Adv. Exp. Med. Biol. 2019, 1185, 323–327. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef]
- King, R.E.; Kent, K.D.; Bomser, J.A. Resveratrol reduces oxidation and proliferation of human retinal pigment epithelial cells via extracellular signal-regulated kinase inhibition. Chem. Biol. Interact. 2005, 151, 143–149. [Google Scholar] [CrossRef]
- He, Q.; Xiao, L.; Shi, Y.; Li, W.; Xin, X. Natural products: Protective effects against ischemia-induced retinal injury. Front. Pharmacol. 2023, 14, 1149708. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Qin, X.; Zhang, J.; Zhang, Z. Echinacoside protects retinal ganglion cells from ischemia/reperfusion-induced injury in the rat retina. Mol. Vis. 2018, 24, 746–758. [Google Scholar]
- Silfen, J.; Yanai, P.; Cabantchik, Z.I. Bioflavonoid effects on in vitro cultures of Plasmodium falciparum. Inhibition of permeation pathways induced in the host cell membrane by the intraerythrocytic parasite. Biochem. Pharmacol. 1988, 37, 4269–4276. [Google Scholar] [CrossRef]
- Roupe, K.A.; Remsberg, C.M.; Yáñez, J.A.; Davies, N.M. Pharmacometrics of stilbenes: Seguing towards the clinic. Curr. Clin. Pharmacol. 2006, 1, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.Y.; Ho, C.T.; Chen, Y.K. Biological actions and molecular effects of resveratrol, pterostilbene, and 3'-hydroxypterostilbene. J. Food Drug Anal. 2017, 25, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yan, Z.; Li, D.; Ma, Y.; Zhou, J.; Sui, Z. Antioxidant and Anti-Inflammatory Effects of Blueberry Anthocyanins on High Glucose-Induced Human Retinal Capillary Endothelial Cells. Oxid. Med. Cell Longev. 2018, 2018, 1862462. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ruzhi, D.; Hua, X.; Zhang, L.; Lu, F.; Coursey, T.G.; Pflugfelder, S.C.; Li, D.Q. Blueberry Component Pterostilbene Protects Corneal Epithelial Cells from Inflammation via Anti-oxidative Pathway. Sci. Rep. 2016, 6, 19408. [Google Scholar] [CrossRef] [PubMed]
- Surien, O.; Masre, S.F.; Basri, D.F.; Ghazali, A.R. Potential Chemopreventive Role of Pterostilbene in Its Modulation of the Apoptosis Pathway. Int. J. Mol. Sci. 2023, 24, 9707. [Google Scholar] [CrossRef]
- Rimando, A.M.; Kalt, W.; Magee, J.B.; Dewey, J.; Ballington, J.R. Resveratrol, pterostilbene, and piceatannol in vaccinium berries. J. Agric. Food Chem. 2004, 52, 4713–4719. [Google Scholar] [CrossRef]
- Xue, E.X.; Lin, J.P.; Zhang, Y.; Sheng, S.R.; Liu, H.X.; Zhou, Y.L.; Xu, H. Pterostilbene inhibits inflammation and ROS production in chondrocytes by activating Nrf2 pathway. Oncotarget 2017, 8, 41988–42000. [Google Scholar] [CrossRef]
- de Queiroz, K.B.; Dos Santos Fontes Pereira, T.; Araújo, M.S.S.; Gomez, R.S.; Coimbra, R.S. Resveratrol Acts Anti-Inflammatory and Neuroprotective in an Infant Rat Model of Pneumococcal Meningitis by Modulating the Hippocampal miRNome. Mol. Neurobiol. 2018, 55, 8869–8884. [Google Scholar] [CrossRef]
- Paul, S.; Rimando, A.M.; Lee, H.J.; Ji, Y.; Reddy, B.S.; Suh, N. Anti-inflammatory action of pterostilbene is mediated through the p38 mitogen-activated protein kinase pathway in colon cancer cells. Cancer Prev. Res. 2009, 2, 650–657. [Google Scholar] [CrossRef]
- Wang, B.; Liu, H.; Yue, L.; Li, X.; Zhao, L.; Yang, X.; Wang, X.; Yang, Y.; Qu, Y. Neuroprotective effects of pterostilbene against oxidative stress injury: Involvement of nuclear factor erythroid 2-related factor 2 pathway. Brain Res. 2016, 1643, 70–79. [Google Scholar] [CrossRef]
- Liu, H.; Wu, X.; Luo, J.; Wang, X.; Guo, H.; Feng, D.; Zhao, L.; Bai, H.; Song, M.; Liu, X.; et al. Pterostilbene Attenuates Astrocytic Inflammation and Neuronal Oxidative Injury After Ischemia-Reperfusion by Inhibiting NF-κB Phosphorylation. Front. Immunol. 2019, 10, 2408. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Zhang, L.; Wang, L. Pterostilbene as a Therapeutic Alternative for Central Nervous System Disorders: A Review of the Current Status and Perspectives. J. Agric. Food Chem. 2023, 71, 14432–14457. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.; Mohandas, S.; Ganesan, K.; Xu, B.; Ramkumar, K.M. New Insights into Dietary Pterostilbene: Sources, Metabolism, and Health Promotion Effects. Molecules 2022, 27, 6316. [Google Scholar] [CrossRef] [PubMed]
- Szilágyi, A.; Takács, B.; Szekeres, R.; Tarjányi, V.; Bombicz, M.; Priksz, D.; Kovács, A.; Juhász, B.; Frecska, E.; Szilvássy, Z.; et al. Therapeutic Properties of Ayahuasca Components in Ischemia/Reperfusion Injury of the Eye. Biomedicines 2022, 10, 997. [Google Scholar] [CrossRef] [PubMed]
- Varga, B.; Gesztelyi, R.; Bombicz, M.; Haines, D.; Szabo, A.M.; Kemeny-Beke, A.; Antal, M.; Vecsernyes, M.; Juhasz, B.; Tosaki, A. Protective effect of alpha-melanocyte-stimulating hormone (α-MSH) on the recovery of ischemia/reperfusion (I/R)-induced retinal damage in a rat model. J. Mol. Neurosci. 2013, 50, 558–570. [Google Scholar] [CrossRef]
- Ozawa, Y.; Toda, E.; Kawashima, H.; Homma, K.; Osada, H.; Nagai, N.; Abe, Y.; Yasui, M.; Tsubota, K. Aquaporin 4 Suppresses Neural Hyperactivity and Synaptic Fatigue and Fine-Tunes Neurotransmission to Regulate Visual Function in the Mouse Retina. Mol. Neurobiol. 2019, 56, 8124–8135. [Google Scholar] [CrossRef]
- Lawrenson, J.G.; Hull, C.C.; Downie, L.E. The effect of blue-light blocking spectacle lenses on visual performance, macular health and the sleep-wake cycle: A systematic review of the literature. Ophthalmic Physiol. Opt. 2017, 37, 644–654. [Google Scholar] [CrossRef]
- Luo, H.; Zhuang, J.; Hu, P.; Ye, W.; Chen, S.; Pang, Y.; Li, N.; Deng, C.; Zhang, X. Resveratrol Delays Retinal Ganglion Cell Loss and Attenuates Gliosis-Related Inflammation From Ischemia-Reperfusion Injury. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3879–3888. [Google Scholar] [CrossRef]
- Chronopoulos, P.; Manicam, C.; Zadeh, J.K.; Laspas, P.; Unkrig, J.C.; Göbel, M.L.; Musayeva, A.; Pfeiffer, N.; Oelze, M.; Daiber, A.; et al. Effects of Resveratrol on Vascular Function in Retinal Ischemia-Reperfusion Injury. Antioxidants 2023, 12, 853. [Google Scholar] [CrossRef]
- Fang, X.L.; Zhang, Q.; Xue, W.W.; Tao, J.H.; Zou, H.D.; Lin, Q.R.; Wang, Y.L. Suppression of cAMP/PKA/CREB signaling ameliorates retinal injury in diabetic retinopathy. Kaohsiung J. Med. Sci. 2023, 39, 916–926. [Google Scholar] [CrossRef]
- Subirada, P.V.; Vaglienti, M.V.; Joray, M.B.; Paz, M.C.; Barcelona, P.F.; Sánchez, M.C. Rapamycin and Resveratrol Modulate the Gliotic and Pro-Angiogenic Response in Müller Glial Cells Under Hypoxia. Front. Cell Dev. Biol. 2022, 10, 855178. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, J.; Guo, R.; Shao, H.; Guo, L. Pterostilbene Protects the Optic Nerves and Retina in a Murine Model of Experimental Autoimmune Encephalomyelitis via Activation of SIRT1 Signaling. Neuroscience 2022, 487, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fan, C.; Yu, L.; Yang, Y.; Jiang, S.; Ma, Z.; Hu, W.; Li, T.; Yang, Z.; Tian, T.; et al. Pterostilbene exerts an anti-inflammatory effect via regulating endoplasmic reticulum stress in endothelial cells. Cytokine 2016, 77, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.R.; Li, S.; Lin, C.C. Effect of resveratrol and pterostilbene on aging and longevity. Biofactors 2018, 44, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Rong, H. Pterostilbene impact on retinal endothelial cells under high glucose environment. Int. J. Clin. Exp. Pathol. 2015, 8, 12589–12594. [Google Scholar] [PubMed]
- Lee, D.; Fu, Z.; Hellstrom, A.; Smith, L.E.H. Therapeutic Effects of Anti-Inflammatory and Anti-Oxidant Nutritional Supplementation in Retinal Ischemic Diseases. Int. J. Mol. Sci. 2024, 25, 5503. [Google Scholar] [CrossRef]
- Remsberg, C.M.; Yáñez, J.A.; Ohgami, Y.; Vega-Villa, K.R.; Rimando, A.M.; Davies, N.M. Pharmacometrics of pterostilbene: Preclinical pharmacokinetics and metabolism, anticancer, antiinflammatory, antioxidant and analgesic activity. Phytother. Res. 2008, 22, 169–179. [Google Scholar] [CrossRef]
- Dvorakova, M.; Landa, P. Anti-inflammatory activity of natural stilbenoids: A review. Pharmacol. Res. 2017, 124, 126–145. [Google Scholar] [CrossRef]
- Lin, Y.J.; Ding, Y.; Wu, J.; Ning, B.T. Pterostilbene as treatment for severe acute pancreatitis. Genet. Mol. Res. 2016, 15, gmr.15038330. [Google Scholar] [CrossRef]
- Li, J.; Xie, C.; Zhuang, J.; Li, H.; Yao, Y.; Shao, C.; Wang, H. Resveratrol attenuates inflammation in the rat heart subjected to ischemia-reperfusion: Role of the TLR4/NF-κB signaling pathway. Mol. Med. Rep. 2015, 11, 1120–1126. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, W.; Wang, Y. PARP-1 and its associated nucleases in DNA damage response. DNA Repair 2019, 81, 102651. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Luo, W.; Wang, Y. Emerging role of PARP-1 and PARthanatos in ischemic stroke. J. Neurochem. 2022, 160, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Seo, K.H.; Yokoyama, W. Chemistry of Pterostilbene and Its Metabolic Effects. J. Agric. Food Chem. 2020, 68, 12836–12841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, T.; Sun, G.F.; Xiao, J.X.; Jiang, L.P.; Tou, F.F.; Qu, X.H.; Han, X.J. Metformin protects against retinal ischemia/reperfusion injury through AMPK-mediated mitochondrial fusion. Free Radic. Biol. Med. 2023, 205, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Gong, B.; Hatala, D.A.; Kern, T.S. Retinal ischemia and reperfusion causes capillary degeneration: Similarities to diabetes. Investig. Ophthalmol. Vis. Sci. 2007, 48, 361–367. [Google Scholar] [CrossRef]
- Gange, W.S.; Qiao, J.B.; Park, P.J.; McDonnell, J.F.; Tan, Z.; Perlman, J.I.; Bu, P. Protection of Retinal Function by Nucleoside Reverse Transcriptase Inhibitors Following Retinal Ischemia/Reperfusion Injury. J. Ocul. Pharmacol. Ther. 2021, 37, 485–491. [Google Scholar] [CrossRef]
- Renner, M.; Stute, G.; Alzureiqi, M.; Reinhard, J.; Wiemann, S.; Schmid, H.; Faissner, A.; Dick, H.B.; Joachim, S.C. Optic Nerve Degeneration after Retinal Ischemia/Reperfusion in a Rodent Model. Front. Cell Neurosci. 2017, 11, 254. [Google Scholar] [CrossRef]
- Kwan, R.Y.C.; Kwan, C.W.; Kor, P.P.K.; Chi, I. Cognitive decline, sensory impairment, and the use of audio-visual aids by long-term care facility residents. BMC Geriatr. 2022, 22, 216. [Google Scholar] [CrossRef]
- Aubin, G.; Phillips, N.; Jaiswal, A.; Johnson, A.P.; Joubert, S.; Bachir, V.; Kehayia, E.; Wittich, W. Visual and cognitive functioning among older adults with low vision before vision rehabilitation: A pilot study. Front. Psychol. 2023, 14, 1058951. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Number of Animals | Treatments | Examinations |
|---|---|---|---|
| 1 | 10 male SD rats | Pterostilbene-treated group =
| Electroretinography, Western blot, and histology |
| 2 | 10 male SD rats | Vehicle (Mucilago)-treated group =
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelles-Taskó, B.; Szekeres, R.; Takács, B.; Szilágyi, A.; Ujvárosy, D.; Bombicz, M.; Priksz, D.; Varga, B.; Gesztelyi, R.; Szabó, Z.; et al. From Nature to Treatment: The Impact of Pterostilbene on Mitigating Retinal Ischemia–Reperfusion Damage by Reducing Oxidative Stress, Inflammation, and Apoptosis. Life 2024, 14, 1148. https://doi.org/10.3390/life14091148
Pelles-Taskó B, Szekeres R, Takács B, Szilágyi A, Ujvárosy D, Bombicz M, Priksz D, Varga B, Gesztelyi R, Szabó Z, et al. From Nature to Treatment: The Impact of Pterostilbene on Mitigating Retinal Ischemia–Reperfusion Damage by Reducing Oxidative Stress, Inflammation, and Apoptosis. Life. 2024; 14(9):1148. https://doi.org/10.3390/life14091148
Chicago/Turabian StylePelles-Taskó, Beáta, Réka Szekeres, Barbara Takács, Anna Szilágyi, Dóra Ujvárosy, Mariann Bombicz, Dániel Priksz, Balázs Varga, Rudolf Gesztelyi, Zoltán Szabó, and et al. 2024. "From Nature to Treatment: The Impact of Pterostilbene on Mitigating Retinal Ischemia–Reperfusion Damage by Reducing Oxidative Stress, Inflammation, and Apoptosis" Life 14, no. 9: 1148. https://doi.org/10.3390/life14091148
APA StylePelles-Taskó, B., Szekeres, R., Takács, B., Szilágyi, A., Ujvárosy, D., Bombicz, M., Priksz, D., Varga, B., Gesztelyi, R., Szabó, Z., Szilvássy, Z., & Juhász, B. (2024). From Nature to Treatment: The Impact of Pterostilbene on Mitigating Retinal Ischemia–Reperfusion Damage by Reducing Oxidative Stress, Inflammation, and Apoptosis. Life, 14(9), 1148. https://doi.org/10.3390/life14091148