

Expression of 9-O-Acetylated Sialic Acid in HPV+ Oral Squamous Cell Carcinoma Cells


 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
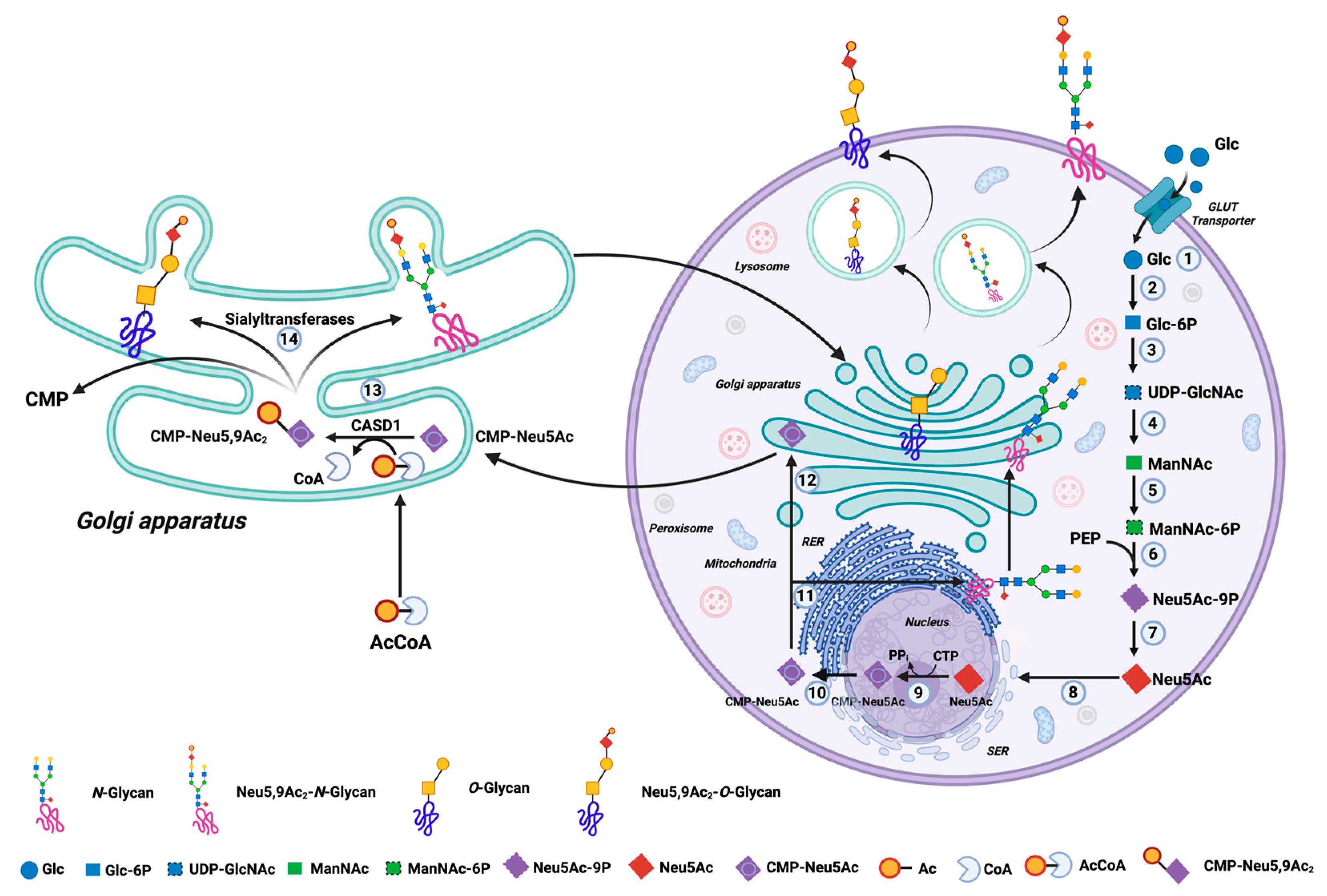
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Macrobrachium Rosenbergii Lectin
2.3. Immunocytochemistry
2.4. Sialidase Treatment of Cells
2.5. Cell Lysate
2.6. Protein Quantification
2.7. One-Dimensional Electrophoresis and Western Blot
2.8. Cellular Proliferation
2.9. Colony Formation Assay
2.10. Statistical Analysis
3. Results
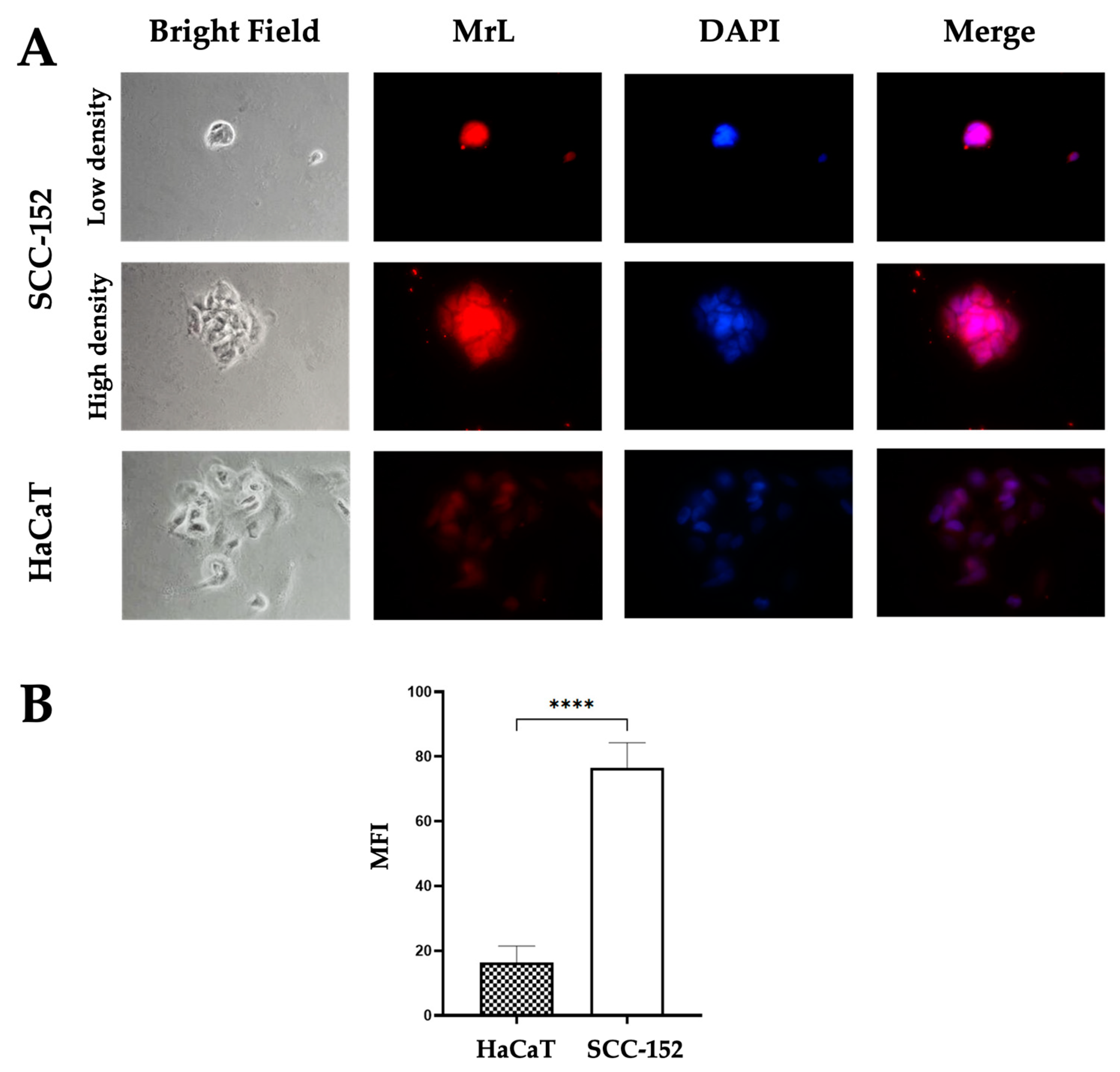
3.1. SCC-152 and HaCaT Cells Express Neu5,9Ac2
3.2. MrL Recognition Is Mediated by Neu5Ac
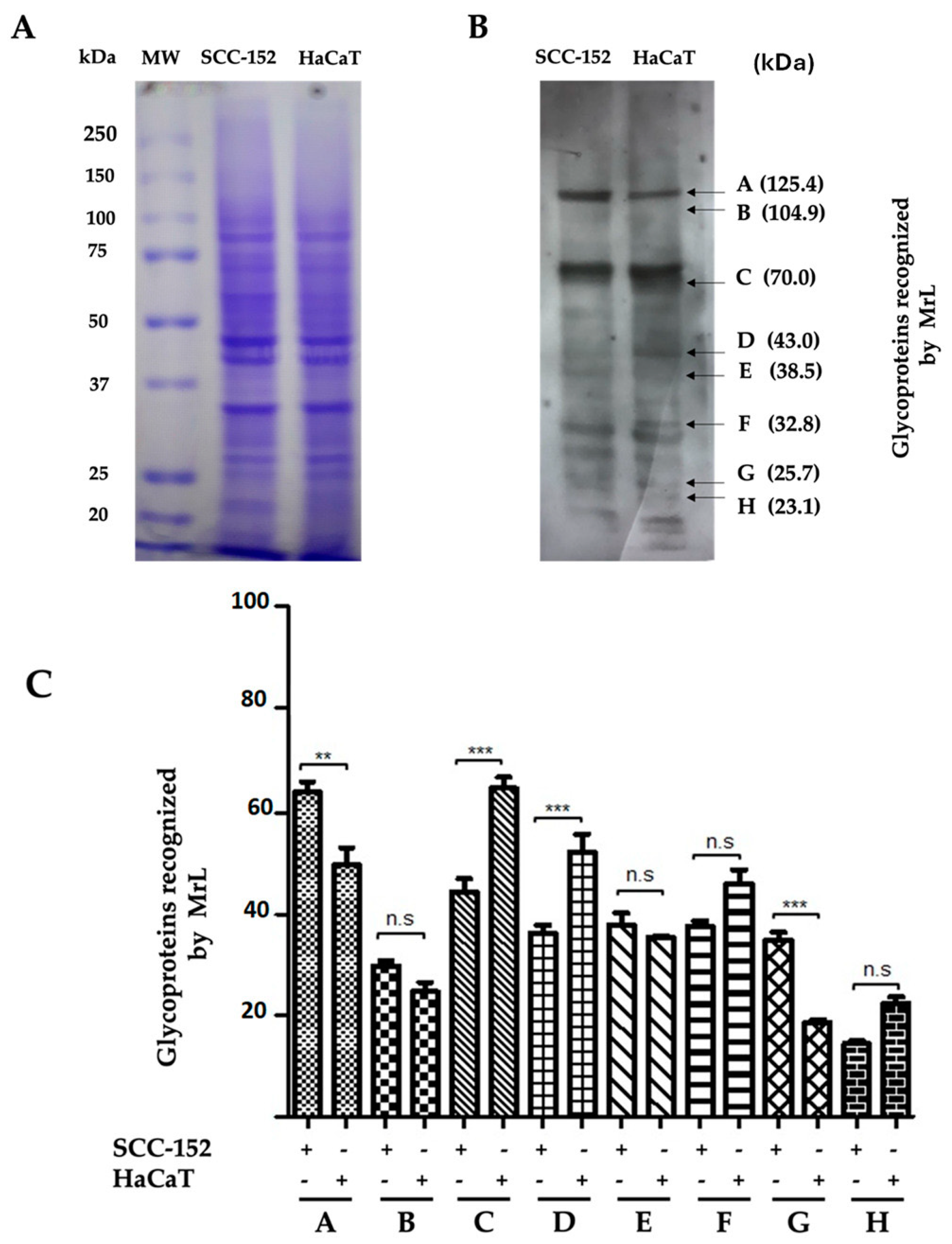
3.3. MrL Lectin Differentially Recognizes Glycoproteins in SCC-152 and HaCaT Cells
3.4. MrL Induces Proliferation of SCC-152 Cells, While It Does Not Affect HaCaT Cells
3.5. MrL Maintains the Structural and Morphological Integrity of SCC-152 Cell Colonies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AcCoa | Acetyl-CoA |
| Achatinin-H | Achatina fulica |
| Asn | Asparagine |
| ATCC | American Type Culture Collection |
| BCA | Bicinchoninic acid |
| BSA | Bovine serum albumin |
| CA-125 | Cancer antigen 125 |
| CASD1 | N-acetylneuraminate (7)-9-O-acetyltransferase |
| CFUs | Colony Forming Units |
| CMP | Cytidine monophosphate |
| CTP | Cytidine triphosphate |
| DAPI | 4′,6-diamidino-2-fenilindol |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DU145 | Prostate cancer cell line |
| E6 | E6 oncoprotein |
| E6p | Ubiquitin ligase E6p |
| EDTA | Ethylenediaminetetraacetic acid |
| EMEM | Earle’s Minimum Essential Medium |
| FBS | Fetal Bovine Serum |
| GalNAc | N-Acetylgalactosamine |
| GD2 | Ganglioside GD2 |
| GD3-9-O-AcSA | Disialoganglioside GD3-O-Acetylate |
| Glc | Glucose |
| Glc-6P | Glucose-6-phosphate |
| GlcNAc | N-Acetylglucosamine |
| HaCaT | Non-tumorigenic human cutaneous keratinocyte |
| HPV | Human papillomavirus |
| MAA | Maackia amurensis |
| ManNAc | N-Acetylmonosamine |
| ManNAc-6-Phosphate | N-Acetylmonosamine-6-phosphate |
| MrL | Macrobrachium rosenbergii lectin |
| MTT | (4,5-dimethyl-2-thiazolyl)-2,5-diphenium-2H-tetrazolium bromide |
| NaCl | Sodium chloride |
| Neu5,9Ac | 9-O-acetylated sialic acid |
| Neu5Ac | Sialic acid |
| Neu5Ac-9-phosphate | N-Acetylneuraminic acid-9-phosphate |
| OAcGD2 | Ganglioside GD2-9-O-acetylated |
| OPMDs | Oral potentially malignant disorders |
| OSCC | Oral squamous cell carcinoma |
| PBMC | Peripheral blood mononuclear cell |
| PBS | Phosphate-Buffered Saline |
| PBS-Ca++ | PBS-Calcio |
| PC-3 | Prostate cancer cell line |
| PEP | Phosphoenolpyruvate |
| PFA | Paraformaldehyde |
| PHA | Phytohemagglutinin |
| pRB | Retinoblastoma protein |
| pre-B ALL | B-cell acute lymphoblastic leukemia |
| PSA | Prostate-specific antigen |
| RER | Rough endoplasmic reticulum |
| SCC | Squamous cell carcinoma |
| SCC-152 | Squamous cell carcinoma cell line |
| SDS-PAGE | Sodium dodecyl-sulfate polyacrylamide gel electrophoresis |
| SER | Smooth endoplasmic reticulum |
| SIAE | Sialiac acid acetylesterase |
| SNA | Sambucus nigra |
| ST6Gal-I | α2,6-sialyltransferase I |
| Thr | Threonine |
| TP | Total protein |
| TSA | Total Neu5Ac |
| UDP-GlcNAc | UDP-N-Acetyl-D-glucosamine |
References
- Fatima, J.; Fatima, E.; Mehmood, F.; Ishtiaq, I.; Khan, M.A.; Khurshid, H.M.S.; Kashif, M. Comprehensive Analysis of Oral Squamous Cell Carcinomas: Clinical, Epidemiological, and Histopathological Insights with a Focus on Prognostic Factors and Survival Time. Cureus 2024, 16, e54394. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, D.; George, N.A.; Thomas, S.; Iype, E.M. Factors associated with delay in diagnosis of oral cancers. Cancer Treat. Res. Commun. 2024, 40, 100831. [Google Scholar] [CrossRef] [PubMed]
- Dhull, A.K.; Atri, R.; Dhankhar, R.; Chauhan, A.K.; Kaushal, V. Major Risk Factors in Head and Neck Cancer: A Retrospective Analysis of 12-Year Experiences. World J. Oncol. 2018, 9, 80–84. [Google Scholar] [CrossRef]
- Dong, L.; Xue, L.; Cheng, W.; Tang, J.; Ran, J.; Li, Y. Comprehensive survival analysis of oral squamous cell carcinoma patients undergoing initial radical surgery. BMC Oral Health 2024, 24, 919. [Google Scholar] [CrossRef]
- Lenoci, D.; Moresco, E.; Cavalieri, S.; Bergamini, C.; Torchia, E.; Botta, L.; Canevari, S.; Trama, A.; Licitra, L.; De Cecco, L. Oral cancer in young adults: Incidence, risk factors, prognosis, and molecular biomarkers. Front. Oncol 2024, 14, 1452909. [Google Scholar] [CrossRef] [PubMed]
- Ajila, V.; Babu, S.; Shetty, V.; Shetty, P.; Devegowda, D.; Ramesh, P.; Natarajan, S. Human papillomavirus in oral squamous cell carcinoma: An institutional study. Clin. Cancer Investig. J. 2021, 10, 102–107. [Google Scholar] [CrossRef]
- Rebello, N.E.; Spadigam, A.E.; Dhupar, A. Burden of High-Risk Human Papillomavirus 16- and 18-Associated Oral Squamous Cell Carcinoma in the Indian Population: A Multiplex Polymerase Chain Reaction Study. Cureus 2024, 16, e73427. [Google Scholar] [CrossRef]
- de Menezes, S.A.F.; Miranda, Y.M.S.; da Silva, Y.M.; Carvalho, T.R.B.; Alves, F.R.S.; Silvestre, R.V.D.; Oliveira-Filho, A.B.; de Alencar Menezes, T.O.; de Souza Fonseca, R.R.; Laurentino, R.V.; et al. Prevalence and Genotyping of HPV in Oral Squamous Cell Carcinoma in Northern Brazil. Pathogens 2022, 11, 1106. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, B.Y.; Lynch, C.F.; Chan, O.T.M.; Goodman, M.T.; Unger, E.R.; Steinau, M.; Thompson, T.D.; Gillison, M.; Lyu, C.; Saraiya, M. HPV Typing of Cancer Workgroup. Human papillomavirus DNA detection, p16INK4a, and oral cavity cancer in a U.S. population. Oral Oncol. 2019, 91, 92–96. [Google Scholar] [CrossRef]
- Ibieta, B.R.; Lizano, M.; Fras-Mendivil, M.; Barrera, J.L.; Carrillo, A.; Ma Ruz-Godoy, L.; Mohar, A. Human papilloma virus in oral squamous cell carcinoma in a Mexican population. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2005, 99, 311–315. [Google Scholar] [CrossRef]
- Das, R.; Kumar, R.; Rai, A.K.; Sarma, A.; Kakoti, L.; Kataki, A.C.; Bhattacharyya, M. HPV and p16 expression association with 5-year survival in oral squamous cell carcinoma patients of North-East India. Adv. Cancer Biol. Metastasis 2024, 10, 100115. [Google Scholar] [CrossRef]
- Giraldi, L.; Collatuzzo, G.; Hashim, D.; Franceschi, S.; Herrero, R.; Chen, C.; Schwartz, S.M.; Smith, E.; Kelsey, K.; McClean, M.; et al. Infection with Human Papilloma Virus (HPV) and risk of subsites within the oral cancer. Cancer Epidemiol. 2021, 75, 102020. [Google Scholar] [CrossRef]
- Pimolbutr, K.; Poomsawat, S.; Na-Ek, N.; Warnakulasuriya, S.; Buajeeb, W. Prevalence of human papillomavirus in oral cancer in Asia: A systematic review and meta-analysis. Oral Dis. 2024. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Ferrall, L.; Gaillard, S.; Wang, C.; Chi, W.Y.; Huang, C.H.; Roden, R.B.S.; Wu, T.C.; Chang, Y.N.; Hung, C.F. Development of DNA Vaccine Targeting E6 and E7 Proteins of Human Papillomavirus 16 (HPV16) and HPV18 for Immunotherapy in Combination with Recombinant Vaccinia Boost and PD-1 Antibody. mBio 2021, 12, e03224-20. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dong, X.; Tang, Y.; Lao, Z.; Li, X.; Lei, J.; Wei, G. Deciphering the mechanisms of HPV E6 mutations in the destabilization of E6/E6AP/p53 complex. Biophys. J. 2022, 121, 1704–1714. [Google Scholar] [CrossRef] [PubMed]
- Magal, S.S.; Jackman, A.; Ish-Shalom, S.; Botzer, L.E.; Gonen, P.; Schlegel, R.; Sherman, L. Downregulation of Bax mRNA expression and protein stability by the E6 protein of human papillomavirus 16. J. Gen. Virol. 2005, 86 Pt 3, 611–621. [Google Scholar] [CrossRef]
- Simmonds, M.; Storey, A. Identification of the regions of the HPV 5 E6 protein involved in Bak degradation and inhibition of apoptosis. Int. J. Cancer 2008, 123, 2260–2266. [Google Scholar] [CrossRef]
- Caldeira, S.; Dong, W.; Tommasino, M. Analysis of E7/Rb associations. Methods Mol. Med. 2005, 119, 363–379. [Google Scholar] [CrossRef]
- Huang, L.W.; Seow, K.M.; Lee, C.C.; Lin, Y.H.; Pan, H.S.; Chen, H.J. Decreased p21 expression in HPV-18 positive cervical carcinomas. Pathol. Oncol. Res. POR 2010, 16, 81–86. [Google Scholar] [CrossRef]
- Santacroce, L.; Di Cosola, M.; Bottalico, L.; Topi, S.; Charitos, I.A.; Ballini, A.; Inchingolo, F.; Cazzolla, A.P.; Dipalma, G. Focus on HPV Infection and the Molecular Mechanisms of Oral Carcinogenesis. Viruses 2021, 13, 559. [Google Scholar] [CrossRef]
- Peng, Q.; Wang, L.; Zuo, L.; Gao, S.; Jiang, X.; Han, Y.; Lin, J.; Peng, M.; Wu, N.; Tang, Y.; et al. HPV E6/E7: Insights into their regulatory role and mechanism in signaling pathways in HPV-associated tumor. Cancer Gene Ther. 2024, 31, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Eichler, J. Protein glycosylation. Curr. Biol. 2019, 29, R229–R231. [Google Scholar] [CrossRef] [PubMed]
- Maccioni, H.J. Glycosylation of glycolipids in the Golgi complex. J. Neurochem. 2007, 103, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R.A.; Pedram, K.; Malaker, S.A.; Batista, P.J.; Smith, B.A.H.; Johnson, A.G.; George, B.M.; Majzoub, K.; Villalta, P.W.; Carette, J.E.; et al. Small RNAs are modified with N-glycans and displayed on the surface of living cells. Cell 2021, 184, 3109–3124.e22. [Google Scholar] [CrossRef]
- Liu, X.; Gao, J.; Sun, Y.; Zhang, D.; Liu, T.; Yan, Q.; Yang, X. Mutation of N-linked glycosylation in EpCAM affected cell adhesion in breast cancer cells. Biol. Chem. 2017, 398, 1119–1126. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, H.; Wu, J.; Yuan, C.; Chen, S.; Liu, S.; Huo, M.; Zhang, C.; He, Y. GALNT1 Enhances Malignant Phenotype of Gastric Cancer via Modulating CD44 Glycosylation to Activate the Wnt/β-catenin Signaling Pathway. Int. J. Biol. Sci. 2022, 18, 6068–6083. [Google Scholar] [CrossRef]
- Lin, W.L.; Lin, Y.S.; Shi, G.Y.; Chang, C.F.; Wu, H.L. Lewisy promotes migration of oral cancer cells by glycosylation of epidermal growth factor receptor. PLoS ONE 2015, 10, e0120162. [Google Scholar] [CrossRef]
- Saitou, A.; Hasegawa, Y.; Fujitani, N.; Ariki, S.; Uehara, Y.; Hashimoto, U.; Saito, A.; Kuronuma, K.; Matsumoto, K.; Chiba, H.; et al. N-glycosylation regulates MET processing and signaling. Cancer Sci. 2022, 113, 1292–1304. [Google Scholar] [CrossRef]
- Li, H.W.; Liu, M.B.; Jiang, X.; Song, T.; Feng, S.X.; Wu, J.Y.; Deng, P.F.; Wang, X.Y. GALNT14 regulates ferroptosis and apoptosis of ovarian cancer through the EGFR/mTOR pathway. Future Oncol. 2022, 18, 149–161. [Google Scholar] [CrossRef]
- Stanley, P.; Moremen, K.W.; Lewis, N.E.; Taniguchi, N.; Aebi, M. N-Glycans. In Essentials of Glycobiology, 4th ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, New York, NY, USA, 2022; pp. 103–116. [Google Scholar]
- Brockhausen, I.; Wandall, H.H.; Hagen, K.G.T.; Stanley, P. O-GalNAc Glycans. In Essentials of Glycobiology, 4th ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2022; pp. 117–128. [Google Scholar]
- Zhu, W.; Zhou, Y.; Guo, L.; Feng, S. Biological function of sialic acid and sialylation in human health and disease. Cell Death Discov. 2024, 10, 415. [Google Scholar] [CrossRef]
- Harduin-Lepers, A.; Krzewinski-Recchi, M.A.; Colomb, F.; Foulquier, F.; Groux-Degroote, S.; Delannoy, P. Sialyltransferases functions in cancers. Front. Biosci. 2012, 4, 499–515. [Google Scholar] [CrossRef]
- Schauer, R.; Kamerling, J.P. Exploration of the Sialic Acid World. Adv. Carbohydr. Chem. Biochem. 2018, 75, 1–213. [Google Scholar] [CrossRef]
- Lewis, A.L.; Chen, X.; Schnaar, R.L.; Varki, A. Sialic Acids and Other Nonulosonic Acids. In Essentials of Glycobiology, 4th ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2022; Chapter 15. Available online: https://www.ncbi.nlm.nih.gov/books/NBK579976/ (accessed on 7 January 2025). [CrossRef]
- Raval, G.N.; Patel, D.D.; Parekh, L.J.; Patel, J.B.; Shah, M.H.; Patel, P.S. Evaluation of serum sialic acid, sialyltransferase and sialoproteins in oral cavity cancer. Oral Dis. 2023, 9, 119–128. [Google Scholar] [CrossRef]
- Dobie, C.; Skropeta, D. Insights into the role of sialylation in cancer progression and metastasis. Br. J. Cancer 2021, 124, 76–90. [Google Scholar] [CrossRef]
- Zhang, Z.; Wuhrer, M.; Holst, S. Serum sialylation changes in cancer. Glycoconj. J. 2018, 35, 139–160. [Google Scholar] [CrossRef]
- Ghosh, S. Sialic acid and biology of life: An introduction. In Sialic Acids and Sialoglycoconjugates in the Biology of Life, Health and Disease; Academic Press: Cambridge, MA, USA, 2020; pp. 1–61. [Google Scholar] [CrossRef]
- İliklerden, Ü.H.; Peksen, C.; Kalayci, T.; Kemik, O. Evaluation of preoperative and postoperative total serum sialic acid levels in patients with colon cancer. Ann. Ital. Chir. 2020, 91, 649–657. [Google Scholar]
- Sata, T.; Roth, J.; Zuber, C.; Stamm, B.; Heitz, P.U. Expression of alpha 2,6-linked sialic acid residues in neoplastic but not in normal human colonic mucosa. A lectin-gold cytochemical study with Sambucus nigra and Maackia amurensis lectins. Am. J. Pathol. 1991, 139, 1435–1448. [Google Scholar]
- Cao, Y.; Karsten, U.; Otto, G.; Bannasch, P. Expression of MUC1, Thomsen-Friedenreich antigen, Tn, sialosyl-Tn, and alpha2,6-linked sialic acid in hepatocellular carcinomas and preneoplastic hepatocellular lesions. Virchows Arch. Int. J. Pathol. 1999, 434, 503–509. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, Y.; Wang, L.; Chen, X.; Su, Z.; Zhang, H.; Yuan, Q.; Wang, S. Sialylated β1, 6 branched N-glycans modulate the adhesion, invasion and metastasis of hepatocarcinoma cells. Biomed. Pharmacother. = Biomed. Pharmacother. 2016, 84, 1654–1661. [Google Scholar] [CrossRef]
- Pally, D.; Pramanik, D.; Hussain, S.; Verma, S.; Srinivas, A.; Kumar, R.V.; Everest-Dass, A.; Bhat, R. Heterogeneity in 2,6-Linked Sialic Acids Potentiates Invasion of Breast Cancer Epithelia. ACS Cent. Sci. 2021, 7, 110–125. [Google Scholar] [CrossRef]
- Zamudio Cañas, R.; Jaramillo Flores, M.E.; Vallejo Ruiz, V.; Delgado Macuil, R.J.; López Gayou, V. Detection of Sialic Acid to Differentiate Cervical Cancer Cell Lines Using a Sambucus nigra Lectin Biosensor. Biosensors 2024, 14, 34. [Google Scholar] [CrossRef]
- López-Morales, D.; Reyes-Leyva, J.; Santos-López, G.; Zenteno, E.; Vallejo-Ruiz, V. Increased expression of sialic acid in cervical biopsies with squamous intraepithelial lesions. Diagn. Pathol. 2010, 5, 74. [Google Scholar] [CrossRef]
- Daniel, D.; Jose, J.; Harish Kumar, A. Is Salivary Sialic Acid a Reliable Biomarker in the Detection of Oral Potentially Malignant Disorder and Oral Squamous Cell Carcinoma. J. Maxillofac. Oral Surg. 2021, 20, 83–89. [Google Scholar] [CrossRef]
- Chaudhari, V.; Pradeep, G.L.; Prakash, N.; Mahajan, A.M. Estimation of salivary sialic acid in oral premalignancy and oral squamous cell carcinoma. Contemp. Clin. Dent. 2016, 7, 451–456. [Google Scholar] [CrossRef]
- Jacob, T.V.; Ramesh, M.; Murali, S.; Ramesh, K.; Sanjay, P.R.; Abraham, P. A non-invasive study to estimate and compare salivary sialic acid level as tumor marker in patients with pre-cancer and oral cancer. J. Cancer Res. Ther. 2016, 12, 634–639. [Google Scholar] [CrossRef]
- Guruaribam, V.D.; Sarumathi, T. Relevance of serum and salivary sialic acid in oral cancer diagnostics. J. Cancer Res. Ther. 2020, 16, 401–404. [Google Scholar] [CrossRef]
- Dhakar, N.; Astekar, M.; Jain, M.; Saawarn, S.; Saawarn, N. Total sialic acid, total protein and total sugar levels in serum and saliva of oral squamous cell carcinoma patients: A case control study. Dent. Res. J. 2013, 10, 343–347. [Google Scholar] [CrossRef]
- Rajaram, S.; Danasekaran, B.P.; Venkatachalapathy, R.; Prashad, K.V.; Rajaram, S. N-acetylneuraminic acid: A scrutinizing tool in oral squamous cell carcinoma diagnosis. Dent. Res. J. 2017, 14, 267–271. [Google Scholar] [CrossRef]
- Shah, M.H.; Telang, S.D.; Shah, P.M.; Patel, P.S. Tissue and serum alpha 2-3- and alpha 2-6-linkage specific sialylation changes in oral carcinogenesis. Glycoconj. J. 2008, 25, 279–290. [Google Scholar] [CrossRef]
- Elgendi, M.; Lyzwinski, L.; Kübler, E.; Shokurov, A.V.; Howard, N.; Menon, C. Advancing cancer detection with portable salivary sialic acid testing. Npj Biosens. 2024, 1, 3. [Google Scholar] [CrossRef]
- Arthisri, A.S.; Sathiyamoorthy, A.; Meenakshi, B.; Chandran, C.R. Ratio of Salivary Sialic Acid to Fucose as Tumor Markers in Potentially Malignant Disorders and Oral Cancer. Contemp Clin Dent. 2020, 11, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Araújo, J.R.C.; Coelho, C.B.; Campos, A.R.; de Azevedo Moreira, R.; de Oliveira Monteiro-Moreira, A.C. Animal Galectins and Plant Lectins as Tools for Studies in Neurosciences. Curr. Neuropharmacol. 2020, 18, 202–215. [Google Scholar] [CrossRef]
- Singh, K.; Agrawal, L.; Gupta, R.; Singh, D.; Kathpalia, M.; Kaur, N. Lectins as a promising therapeutic agent for breast cancer: A review. Breast Dis. 2024, 43, 193–211. [Google Scholar] [CrossRef] [PubMed]
- Visser, E.A.; Moons, S.J.; Timmermans, S.B.P.E.; de Jong, H.; Boltje, T.J.; Büll, C. Sialic acid O-acetylation: From biosynthesis to roles in health and disease. J. Biol. Chem. 2021, 297, 100906. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Mandal, C. O-acetylated sialic acids: Multifaceted role in childhood acute lymphoblastic leukaemia. Biotechnol. J. 2009, 4, 361–374. [Google Scholar] [CrossRef]
- Cheresh, D.A.; Reisfeld, R.A.; Varki, A.P. O-acetylation of disialoganglioside GD3 by human melanoma cells creates a unique antigenic determinant. Science 1984, 225, 844–846. [Google Scholar] [CrossRef]
- Fahr, C.; Schauer, R. Detection of sialic acids and gangliosides with special reference to 9-O-acetylated species in basaliomas and normal human skin. J. Investig. Dermatol. 2001, 116, 254–260. [Google Scholar] [CrossRef]
- Cavdarli, S.; Dewald, J.H.; Yamakawa, N.; Guérardel, Y.; Terme, M.; Le Doussal, J.M.; Delannoy, P.; Groux-Degroote, S. Identification of 9-O-acetyl-N-acetylneuraminic acid (Neu5,9Ac2) as main O-acetylated sialic acid species of GD2 in breast cancer cells. Glycoconj. J. 2019, 36, 79–90. [Google Scholar] [CrossRef]
- Zenteno, R.; Vazquez, L.; Sierra, C.; Pereyra, A.; Slomianny, M.C.; Bouquelet, S.; Zenteno, E. Chemical characterization of the lectin from the freshwater prawn Macrobrachium rosenbergii (De Man) by MALDI-TOF. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2000, 127, 243–250. [Google Scholar] [CrossRef]
- Zenteno, R.; Vázquez, L.; Martínez-Cairo, S.; Bouquelet, S.; Agundis, C.; Zenteno, E. Identification of lectin isoforms in juvenile freshwater prawns Macrobrachium rosenbergii (DeMan, 1879). Glycoconj. J. 2000, 17, 339–347. [Google Scholar] [CrossRef]
- Pérez-Campos-Mayoral, L.; Ruiz-Argüelles, A.; Pérez-Romano, B.; Zenteno, E.; Hernández-Cruz, P.; Martínez-Cruz, R.; Martínez-Cruz, M.; Pina-Canseco, S.; Pérez-Campos, E. Potential use of the Macrobrachium rosenbergii lectin for diagnosis of T-cell acute lymphoblastic leukemia. Tohoku J. Exp. Med. 2008, 214, 11–16. [Google Scholar] [CrossRef] [PubMed]
- American Type Culture Collection. Available online: https://www.atcc.org/products/crl-3240 (accessed on 7 January 2025).
- Cellosaurus UPCI-SCC-152. Available online: https://www.cellosaurus.org/CVCL_C058 (accessed on 7 January 2025).
- Xu, C.; Yan, T.; Yang, J. OVOL1 inhibits oral squamous cell carcinoma growth and metastasis by suppressing zinc finger E-box binding homeobox 1. Int. J. Clin. Exp. Pathol. 2019, 12, 2801–2808. [Google Scholar]
- Ziemann, F.; Arenz, A.; Preising, S.; Wittekindt, C.; Klussmann, J.P.; Engenhart-Cabillic, R.; Wittig, A. Increased sensitivity of HPV-positive head and neck cancer cell lines to x-irradiation ± Cisplatin due to decreased expression of E6 and E7 oncoproteins and enhanced apoptosis. Am. J. Cancer Res. 2015, 5, 1017–1031. [Google Scholar]
- Pekarek, L.; GarridoGil, M.J.; SánchezCendra, A.; Cassinello, J.; Pekarek, T.; FraileMartinez, O.; García-Montero, C.; López-González, L.; Rios-Parra, A.; Álvarez-Mon, M.; et al. Emerging histological and serological biomarkers in oral squamous cell carcinoma: Applications in diagnosis, prognosis evaluation and personalized therapeutics (Review). Oncol. Rep. 2023, 50, 213. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, S.; Sethy, M.; Bhue, S.; Mohanta, B.K.; Dixit, A. Identification of therapeutically potential targets and their ligands for the treatment of OSCC. Front. Oncol. 2022, 12, 910494. [Google Scholar] [CrossRef]
- He, K.; Baniasad, M.; Kwon, H.; Caval, T.; Xu, G.; Lebrilla, C.; Hommes, D.W.; Bertozzi, C. Decoding the glycoproteome: A new frontier for biomarker discovery in cancer. J. Hematol. Oncol. 2024, 17, 12. [Google Scholar] [CrossRef]
- Guo, Y.; Jia, W.; Yang, J.; Zhan, X. Cancer glycomics offers potential biomarkers and therapeutic targets in the framework of 3P medicine. Front. Endocrinol. 2022, 13, 970489. [Google Scholar] [CrossRef]
- Mayoral, M.A.; Mayoral, C.; Meneses, A.; Villalvazo, L.; Guzman, A.; Espinosa, B.; Ochoa, J.L.; Zenteno, E.; Guevara, J. Identification of galectin-3 and mucin-type O-glycans in breast cancer and its metastasis to brain. Cancer Investig. 2008, 26, 615–623. [Google Scholar] [CrossRef]
- Hernández-Maqueda, J.G.; Luna-Ulloa, L.B.; Santoyo-Ramos, P.; Castañeda-Patlán, M.C.; Robles-Flores, M. Protein kinase C delta negatively modulates canonical Wnt pathway and cell proliferation in colon tumor cell lines. PLoS ONE 2013, 8, e58540. [Google Scholar] [CrossRef]
- Katz, D.; Ito, E.; Lau, K.S.; Mocanu, J.D.; Bastianutto, C.; Schimmer, A.D.; Liu, F.F. Increased efficiency for performing colony formation assays in 96-well plates: Novel applications to combination therapies and high-throughput screening. BioTechniques 2008, 44, ix–xiv. [Google Scholar] [CrossRef]
- Josic, D.; Martinovic, T.; Pavelic, K. Glycosylation and metastases. Electrophoresis 2019, 40, 140–150. [Google Scholar] [CrossRef]
- Vajaria, B.N.; Patel, K.R.; Begum, R.; Patel, P.S. Sialylation: An Avenue to Target Cancer Cells. Pathol. Oncol. Res. POR 2016, 22, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ding, J. Sialylation is involved in cell fate decision during development, reprogramming and cancer progression. Protein Cell 2019, 10, 550–565. [Google Scholar] [CrossRef] [PubMed]
- Holíková, Z.; Hrdlicková-Cela, E.; Plzák, J.; Smetana, K., Jr.; Betka, J.; Dvoránková, B.; Esner, M.; Wasano, K.; André, S.; Kaltner, H.; et al. Defining the glycophenotype of squamous epithelia using plant and mammalian lectins. Differentiation-dependent expression of alpha2,6- and alpha2,3-linked N-acetylneuraminic acid in squamous epithelia and carcinomas, and its differential effect on binding of the endogenous lectins galectins-1 and -3. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2002, 110, 845–856. [Google Scholar] [CrossRef]
- Vajaria, B.N.; Patel, K.R.; Begum, R.; Patel, J.B.; Shah, F.D.; Joshi, G.M.; Patel, P.S. Salivary glyco-sialylation changes monitors oral carcinogenesis. Glycoconj. J. 2014, 31, 649–659. [Google Scholar] [CrossRef]
- Varki, N.M.; Varki, A. Diversity in cell surface sialic acid presentations: Implications for biology and disease. Lab. Investig. J. Tech. Methods Pathol. 2007, 87, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Cavdarli, S.; Schröter, L.; Albers, M.; Baumann, A.M.; Vicogne, D.; Le Doussal, J.M.; Mühlenhoff, M.; Delannoy, P.; Groux-Degroote, S. Role of Sialyl-O-Acetyltransferase CASD1 on GD2 Ganglioside O-Acetylation in Breast Cancer Cells. Cells 2021, 10, 1468. [Google Scholar] [CrossRef]
- Kawashima, I.; Nagata, I.; Tai, T. Immunocytochemical analysis of gangliosides in rat primary cerebellar cultures using specific monoclonal antibodies. Brain Res. 1996, 732, 75–86. [Google Scholar] [CrossRef]
- Wang, Y.H. Sialidases From Clostridium perfringens and Their Inhibitors. Front. Cell. Infect. Microbiol. 2020, 9, 462. [Google Scholar] [CrossRef]
- Joo, E.J.; Wasik, B.R.; Parrish, C.; Paz, H.; Mϋhlenhoff, M.; Abdel-Azim, H.; Groffen, J.; Heisterkamp, N. Pre-B acute lymphoblastic leukemia expresses cell surface nucleolin as a 9-O-acetylated sialoglycoprotein. Sci. Rep. 2018, 8, 17174. [Google Scholar] [CrossRef]
- Chowdhury, S.; Bandyopadhyay, S.; Mandal, C.; Chandra, S.; Mandal, C. Flow-cytometric monitoring of disease-associated expression of 9-O-acetylated sialoglycoproteins in combination with known CD antigens, as an index for MRD in children with acute lymphoblastic leukaemia: A two-year longitudinal follow-up study. BMC Cancer 2008, 8, 40. [Google Scholar] [CrossRef]
- Zhang, X.; Pan, C.; Zhou, L.; Cai, Z.; Zhao, S.; Yu, D. Knockdown of ST6Gal-I increases cisplatin sensitivity in cervical cancer cells. BMC Cancer 2016, 16, 949. [Google Scholar] [CrossRef]
- Wei, A.; Fan, B.; Zhao, Y.; Zhang, H.; Wang, L.; Yu, X.; Yuan, Q.; Yang, D.; Wang, S. ST6Gal-I overexpression facilitates prostate cancer progression via the PI3K/Akt/GSK-3β/β-catenin signaling pathway. Oncotarget 2016, 7, 65374–65388. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Bandyopadhyay, S.; Pal, S.; Das, B.; Bhattacharya, D.K.; Mandal, C. Increased interferon gamma production by peripheral blood mononuclear cells in response to stimulation of overexpressed disease-specific 9-O-acetylated sialoglycoconjugates in children suffering from acute lymphoblastic leukaemia. Br. J. Haematol. 2005, 128, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, M.; Wipfler, D.; Merling, A.; Cao, Y.; Claus, C.; Kniep, B.; Sadick, H.; Bergler, W.; Vlasak, R.; Schwartz-Albiez, R. Differential surface expression and possible function of 9-O- and 7-O-acetylated GD3 (CD60 b and c) during activation and apoptosis of human tonsillar B and T lymphocytes. Glycoconj. J. 2006, 23, 627–638. [Google Scholar] [CrossRef]
- Tonelli, Q.; Meints, R.H. Sialic acid: A specific role in hematopoietic spleen colony formation. J. Supramol. Struct. 1978, 8, 67–78. [Google Scholar] [CrossRef]
- Varki, A. Sialic acids in human health and disease. Trends Mol. Med. 2008, 14, 351–360. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Martínez, H.; Jiménez-Castillo, V.; Illescas-Barbosa, D.; Ávila-Curiel, B.X.; Hernández-Huerta, M.T.; Díaz-Castillejos, R.; Torres-Rosas, R.; Zenteno, E.; Pereyra-Morales, M.A.; Solórzano-Mata, C.J. Expression of 9-O-Acetylated Sialic Acid in HPV+ Oral Squamous Cell Carcinoma Cells. Life 2025, 15, 663. https://doi.org/10.3390/life15040663
Sánchez-Martínez H, Jiménez-Castillo V, Illescas-Barbosa D, Ávila-Curiel BX, Hernández-Huerta MT, Díaz-Castillejos R, Torres-Rosas R, Zenteno E, Pereyra-Morales MA, Solórzano-Mata CJ. Expression of 9-O-Acetylated Sialic Acid in HPV+ Oral Squamous Cell Carcinoma Cells. Life. 2025; 15(4):663. https://doi.org/10.3390/life15040663
Chicago/Turabian StyleSánchez-Martínez, Hugo, Victoria Jiménez-Castillo, Daniela Illescas-Barbosa, Beatriz Xochitl Ávila-Curiel, María Teresa Hernández-Huerta, Risk Díaz-Castillejos, Rafael Torres-Rosas, Edgar Zenteno, Mohamed Alí Pereyra-Morales, and Carlos Josué Solórzano-Mata. 2025. "Expression of 9-O-Acetylated Sialic Acid in HPV+ Oral Squamous Cell Carcinoma Cells" Life 15, no. 4: 663. https://doi.org/10.3390/life15040663
APA StyleSánchez-Martínez, H., Jiménez-Castillo, V., Illescas-Barbosa, D., Ávila-Curiel, B. X., Hernández-Huerta, M. T., Díaz-Castillejos, R., Torres-Rosas, R., Zenteno, E., Pereyra-Morales, M. A., & Solórzano-Mata, C. J. (2025). Expression of 9-O-Acetylated Sialic Acid in HPV+ Oral Squamous Cell Carcinoma Cells. Life, 15(4), 663. https://doi.org/10.3390/life15040663