
Haloferax volcanii, as a Novel Tool for Producing Mammalian Olfactory Receptors Embedded in Archaeal Lipid Bilayer

Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Strains and Culture Condition.
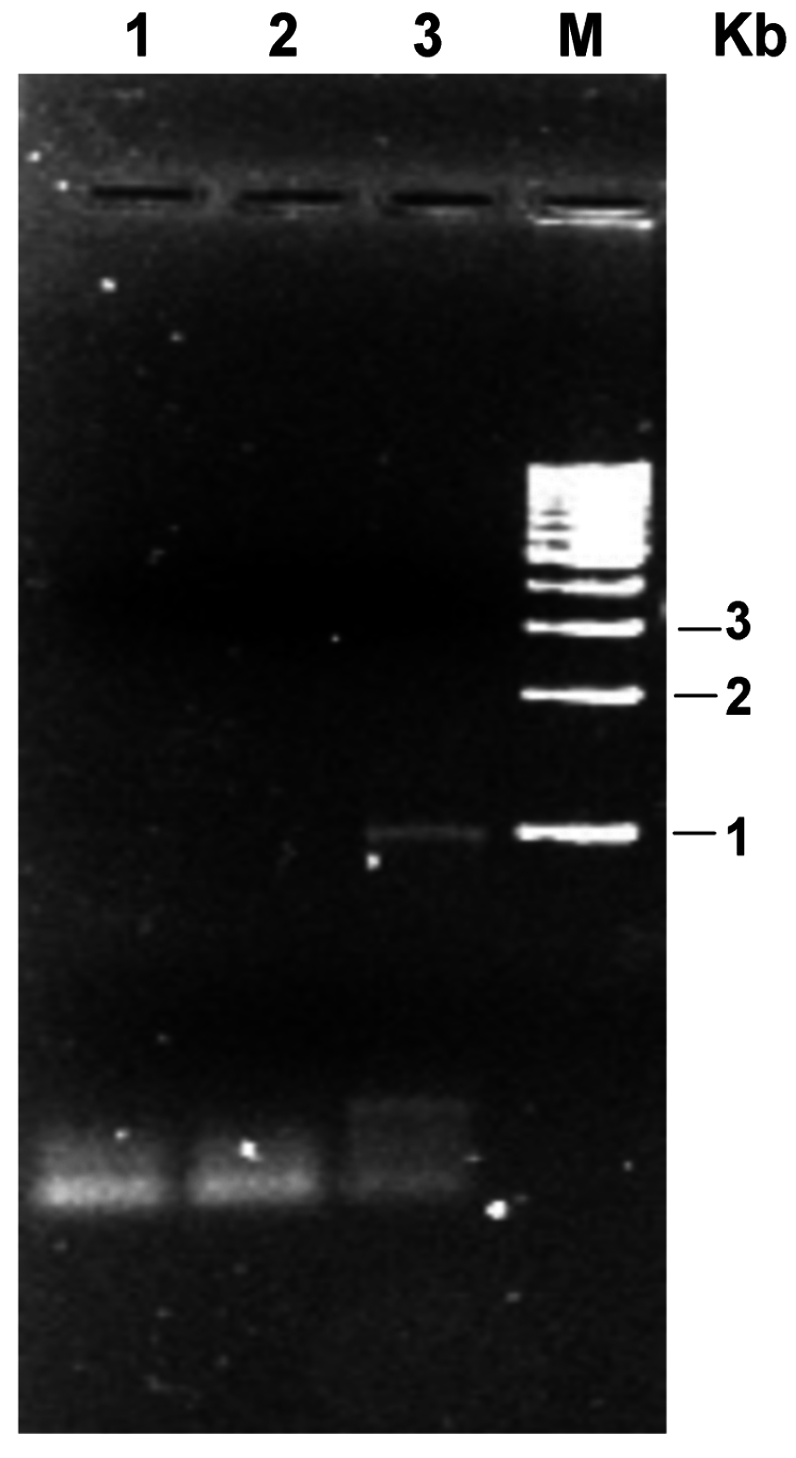
2.3. Extraction of Total Norway Rat Genomic DNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5'→3') |
|---|---|
| I7 for | gctagcatggagcgaaggaaccacag |
| I7 His rev | ggtaccctaaccaattttgctgcctttgt |
| Linker 1 | catgggggctagcgattgcgatccgattcggtac |
| Linker 2 | cgaatcggatcgcaatcgctagccc |
2.4. Vectors
2.5. Transformation of Hfx. volcanii
2.6. Reverse Transcription—Polymerase Chain Reaction (RT-PCR)
2.7. Sequencing
2.8. Isolation of Hfx. volcanii Membranes
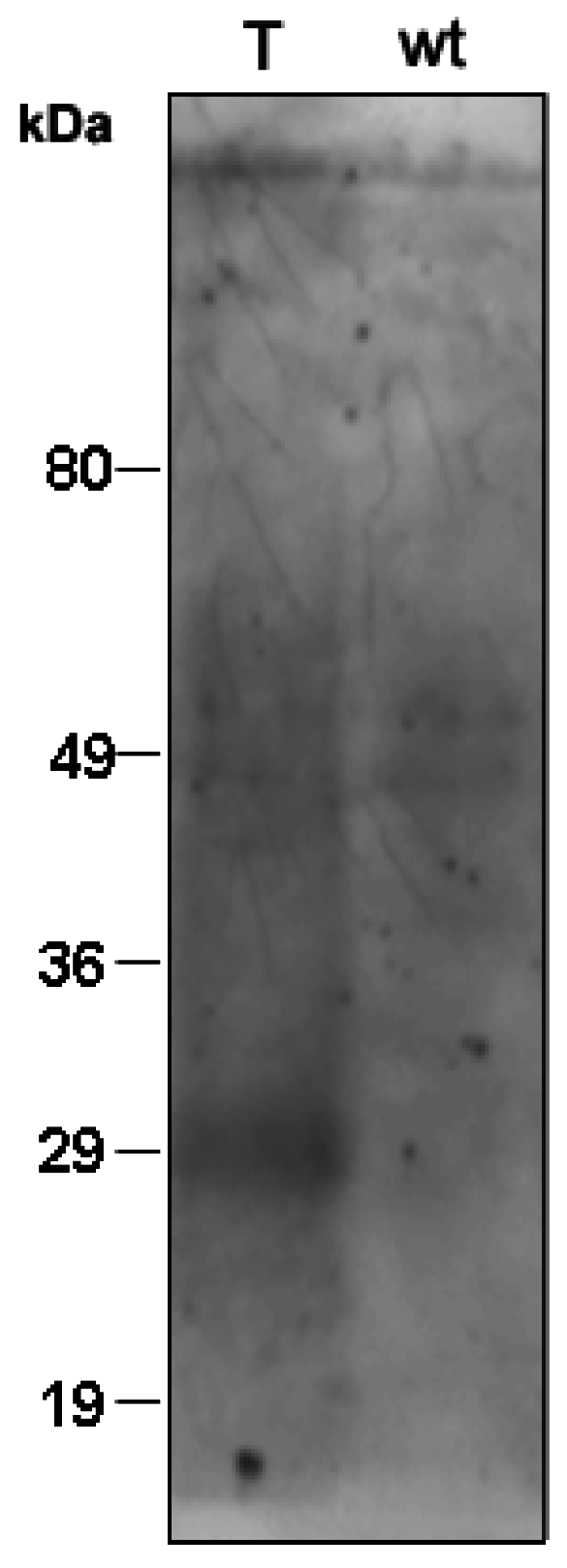
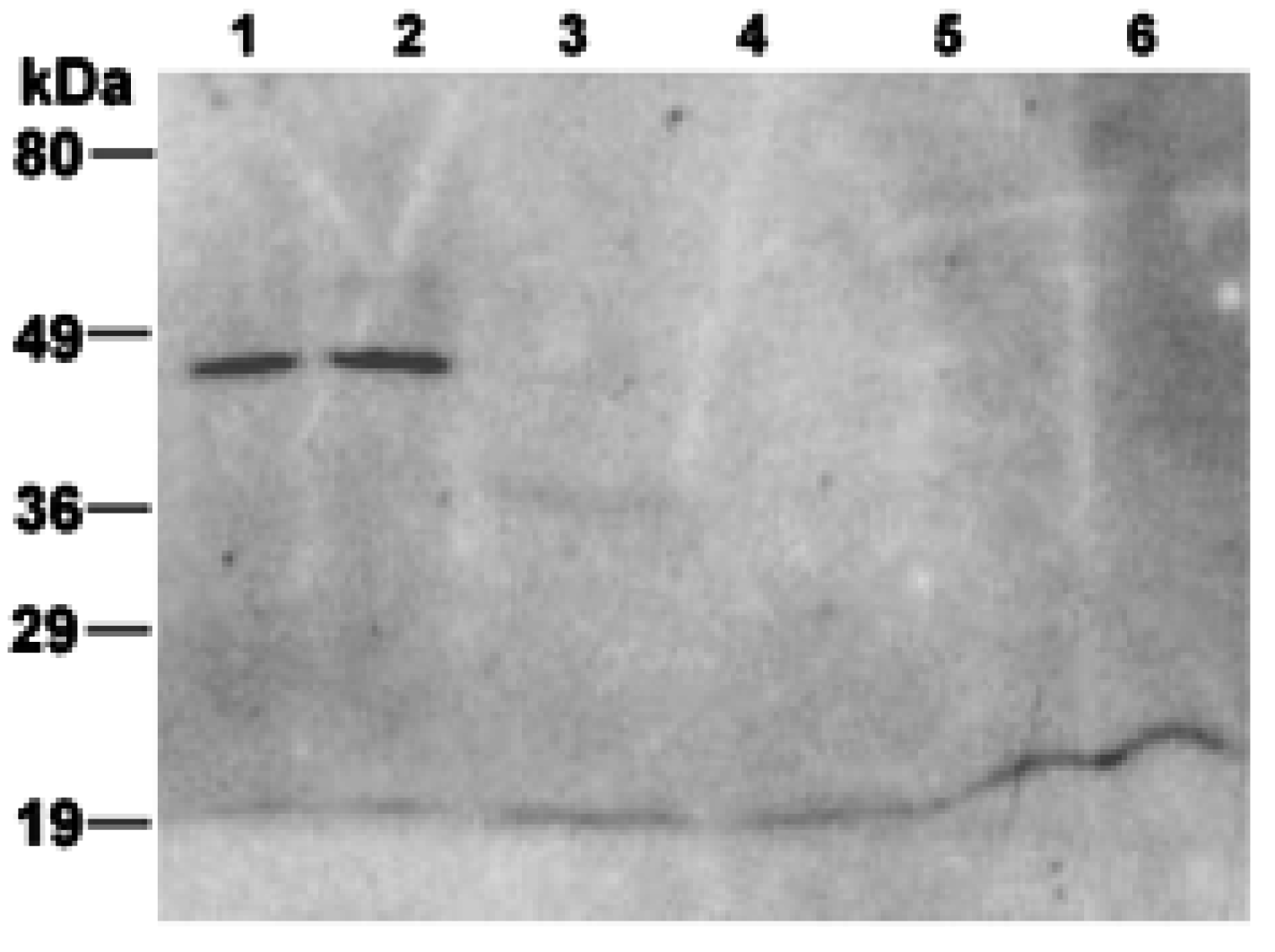
2.9. Immunoblot Analysis
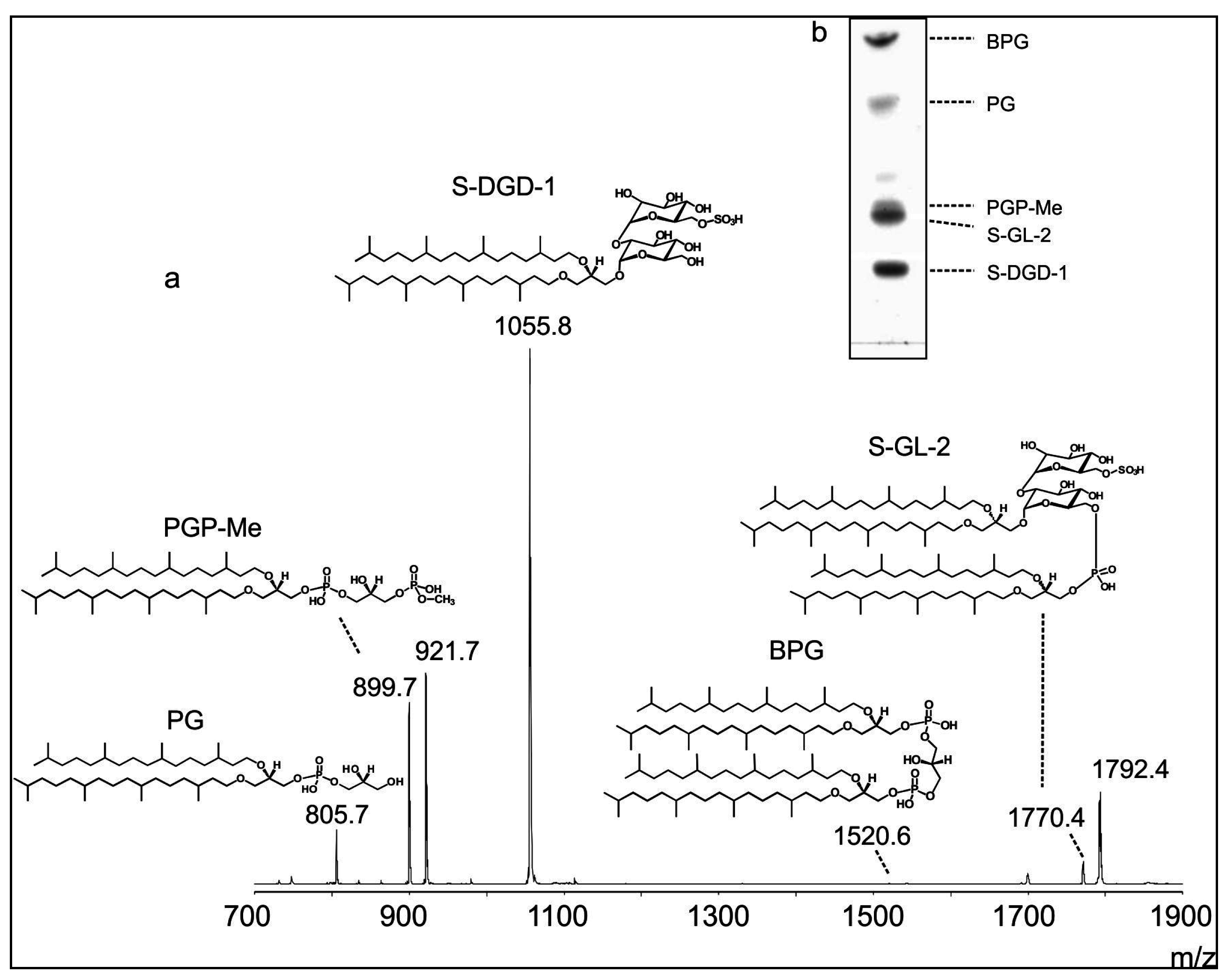
2.10. Lipid Extraction and HPTLC Analysis
2.11. MALDI-TOF/MS Lipid Analysis
3. Results and Discussion





Acknowledgments
Author Contributions
Conflicts of Interest
References
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Touhara, K. Mammalian olfactory receptors: Pharmacology, G protein coupling and desensitization. Cell. Mol. Life Sci. 2009, 66, 3743–3753. [Google Scholar] [CrossRef] [PubMed]
- Hoover, K.C. Evolution of olfactory receptors. Methods Mol. Biol. 2013, 1003, 241–249. [Google Scholar] [PubMed]
- Pierce, K.L.; Premont, R.T.; Lefkowitz, R.J. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Firestein, S. How the olfactory system makes sense of scents. Nature 2001, 413, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Ivic, L.; Otaki, J.; Hashimoto, M.; Mikoshiba, K.; Firestein, S. Functional expression of a mammalian odorant receptor. Science 1998, 279, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Kurland, M.D.; Newcomer, M.B.; Peterlin, Z.; Ryan, K.; Firestein, S.; Batista, V.S. Discrimination of saturated aldehydes by the rat I7 olfactory receptor. Biochemistry 2010, 49, 6302–6304. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.S. Analysis of the molecular basis for octanal interactions in the expressed rat 17 olfactory receptor. Chem. Senses 2000, 25, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.E.; Floriano, W.B.; Vaidehi, N.; Goddard, W.A. Predicted 3-D structures for mouse I7 and rat I7 olfactory receptors and comparison of predicted odor recognition profiles with experiment. Chem. Senses 2004, 29, 595–616. [Google Scholar] [CrossRef] [PubMed]
- Minic, J.; Persuy, M.A.; Godel, E.; Aioun, J.; Connerton, I.; Salesse, R.; Pajot-Augy, E. Functional expression of olfactory receptors in yeast and development of a bioassay for odorant screening. FEBS J. 2005, 272, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Matarazzo, V.; Clot-Faybesse, O.; Marcet, B.; Guiraudie-Capraz, G.; Atanasova, B.; Devauchelle, G.; Cerutti, M.; Etiévant, P.; Ronin, C. Functional characterization of two human olfactory receptors expressed in the baculovirus Sf9 insect cell system. Chem. Senses 2005, 30, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Kubota, M.; Roberts, R.W.; Chi, Q.; Matsunami, H. RTP family members induce functional expression of mammalian odorant receptors. Cell 2004, 119, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Von Dannecker, L.E.; Mercadante, A.F.; Malnic, B. Ric-8B promotes functional expression of odorant receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 9310–9314. [Google Scholar] [CrossRef] [PubMed]
- Shirokova, E.; Schmiedeberg, K.; Bedner, P.; Niessen, H.; Willecke, K.; Raguse, J.D.; Meyerhof, W.; Krautwurst, D. Identification of specific ligands for orphan olfactory receptors. G protein-dependent agonism and antagonism of odorants. J. Biol. Chem. 2005, 280, 11807–11815. [Google Scholar] [CrossRef] [PubMed]
- Söhlemann, P.; Soppa, J.; Oesterhelt, D.; Lohse, M.J. Expression of beta-2-adrenoceptors in halobacteria. Nauny. Schmiedebergs Arch. Pharmacol. 1997, 355, 150–160. [Google Scholar] [CrossRef]
- Patenge, N.; Soppa, J. Extensive proteolysis inhibits high-level production of eukaryal G-protein coupled receptors in the archeon Haloferax volcanii. FEMS Microbiol. Lett. 1999, 171, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Hartman, A.L.; Norais, C.; Badger, J.H.; Delmas, S.; Haldenby, S.; Madupu, R.; Robinson, J.; Khouri, H.; Ren, Q.; Lowe, T.M.; et al. The complete genome sequence of Haloferax volcanii DS2, a model archaeon. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Christian, J.H.; Waltho, J.A. Solute concentrations within cells of halophilic and non-halophilic bacteria. Biochim. Biophys. Acta 1962, 65, 506–508. [Google Scholar] [CrossRef] [PubMed]
- Allers, T.; Ngo, H.P.; Mevarech, M.; Lloyd, R.G. Development of additional selectable markers for the halophilic archaeon Haloferax volcanii based on the leuB and trpA genes. Appl. Environ. Microbiol. 2004, 70, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Bitan-Banin, G.; Ortenberg, R.; Mevarech, M. Development of a gene knockout system for the halophilic archaeon Haloferax volcanii by use of the pyrE gene. J. Bacteriol. 2003, 185, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.L.; Dyall-Smith, M.L. Sequence and expression of a halobacterial beta-galactosidase gene. Mol. Microbiol. 2000, 36, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Large, A.; Stamme, C.; Lange, C.; Duan, Z.; Allers, T.; Soppa, J.; Lund, P.A. Characterization of a tightly controlled promoter of the halophilic archaeon Haloferax volcanii and its use in the analysis of the essential cct1 gene. Mol. Microbiol. 2007, 66, 1092–1106. [Google Scholar] [CrossRef] [PubMed]
- Kates, M. Ether linked lipids in extremely halophilic bacteria. In Ether Lipids, Chemistry and Biology; Snyder, F., Ed.; Academic Press: New York, NY, USA, 1972; pp. 351–398. [Google Scholar]
- Kates, M. Membrane lipids of Archaea. In The Biochemistry of Archaea (Archaebacteria); Kates, M., Kushner, D.J., Matheson, A.T., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 261–295. [Google Scholar]
- Ingrosso, C.; Bianco, G.V.; Lopalco, P.; Tamborra, M.; Curri, M.L.; Corcelli, A.; Bruno, G.; Agostiano, A.; Siciliano, P.; Striccoli, M. Surface chemical functionalization of single walled carbon nanotubes with a bacteriorhodopsin mutant. Nanoscale 2012, 4, 6434–6441. [Google Scholar] [CrossRef] [PubMed]
- Ingrosso, C.; Bianco, G.V.; Corricelli, M.; Corcelli, A.; Lobasso, S.; Bruno, G.; Agostiano, A.; Striccoli, M.; Curri, M.L. Hybrid charge transfer complexes based on archaeal glycolipids wrapping single walled carbon nanotubes. Chem. Commun. 2013, 49, 6941–6943. [Google Scholar] [CrossRef]
- Cline, S.W.; Lam, W.L.; Charlebois, R.L.; Schalkwyk, L.C.; Doolittle, W.F. Transformation methods for halophilic archaebacteria. Can. J. Microbiol. 1989, 35, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef] [PubMed]
- Kates, M. Techniques of Lipidology; Elsevier: Amsterdam, The Netherlands, 1986. [Google Scholar]
- Corcelli, A.; Lobasso, S. Characterization of lipids of halophilic Archaea. In Methods in Microbiology—Extremophile Microorganisms and the Methods to Handle Them; Rainey, F.A., Oren, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 35, pp. 591–613. [Google Scholar]
- Lobasso, S.; Lopalco, P.; Mascolo, G.; Corcelli, A. Lipids of the ultra-thin square microorganism Haloquadratum walsbyi. Archaea 2008, 2, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Angelini, R.; Vitale, R.; Patil, V.A.; Cocco, T.; Ludwig, B.; Greenberg, M.L.; Corcelli, A. Lipidomics of intact mitochondria by MALDI-TOF/MS. J. Lipid Res. 2012, 53, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, S.C.; Kates, M.; Juez, G.; Rodriguez-Valera, F.; Kushner, D.J. Polar lipids of an extremely halophilic bacterial strain (R-4) isolated from salt ponds in Spain. Biochim. Biophys. Acta 1982, 711, 19–25. [Google Scholar] [CrossRef]
- Sprott, G.D.; Laroque, S.; Cadotte, N.; Dicaire, C.J.; McGee, M.; Brisson, J.I. Novel polar lipids of halophilic eubacterium Planococcus H8 and archaeon Haloferax volcanii. Biochim. Biophys. Acta 2003, 1633, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Corcelli, A.; Colella, M.; Mascolo, G.; Fanizzi, F.P.; Kates, M. A novel glycolipid and phospholipid in the purple membrane. Biochemistry 2000, 39, 3318–3326. [Google Scholar] [CrossRef] [PubMed]
- Corcelli, A. The cardiolipin analogues of Archaea. Biochim. Biophys. Acta 2009, 1788, 2101–2106. [Google Scholar] [CrossRef] [PubMed]
- Vidic, J.; Grosclaude, J.; Persuy, M.A.; Aioun, J.; Salesse, R.; Pajot-Augy, E. Quantitative assessment of olfactory receptors activity in immobilized nanosomes: A novel concept for bioelectronic nose. Lab Chip 2006, 6, 1026–1032. [Google Scholar] [CrossRef] [PubMed]
- Glatz, R.; Bailey-Hill, K. Mimicking nature’s noses: From receptor deorphaning to olfactory biosensing. Prog. Neurobiol. 2011, 93, 270–296. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Wu, C.; Liu, Q.; Huang, L.; Wang, P. Recent advances in olfactory receptor-based biosensors. Biosens. Bioelectron. 2013, 42, 570–580. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobasso, S.; Vitale, R.; Lopalco, P.; Corcelli, A. Haloferax volcanii, as a Novel Tool for Producing Mammalian Olfactory Receptors Embedded in Archaeal Lipid Bilayer. Life 2015, 5, 770-782. https://doi.org/10.3390/life5010770
Lobasso S, Vitale R, Lopalco P, Corcelli A. Haloferax volcanii, as a Novel Tool for Producing Mammalian Olfactory Receptors Embedded in Archaeal Lipid Bilayer. Life. 2015; 5(1):770-782. https://doi.org/10.3390/life5010770
Chicago/Turabian StyleLobasso, Simona, Rita Vitale, Patrizia Lopalco, and Angela Corcelli. 2015. "Haloferax volcanii, as a Novel Tool for Producing Mammalian Olfactory Receptors Embedded in Archaeal Lipid Bilayer" Life 5, no. 1: 770-782. https://doi.org/10.3390/life5010770
APA StyleLobasso, S., Vitale, R., Lopalco, P., & Corcelli, A. (2015). Haloferax volcanii, as a Novel Tool for Producing Mammalian Olfactory Receptors Embedded in Archaeal Lipid Bilayer. Life, 5(1), 770-782. https://doi.org/10.3390/life5010770