Sun Exposure Shapes Functional Grouping of Fungi in Cryptoendolithic Antarctic Communities

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Area
2.2. DNA Extraction, Metabarcoding Sequencing, and Bioinformatic Analysis
2.3. Statistical Analysis
3. Results
3.1. Bionformatic Analysis and Guild Assignment
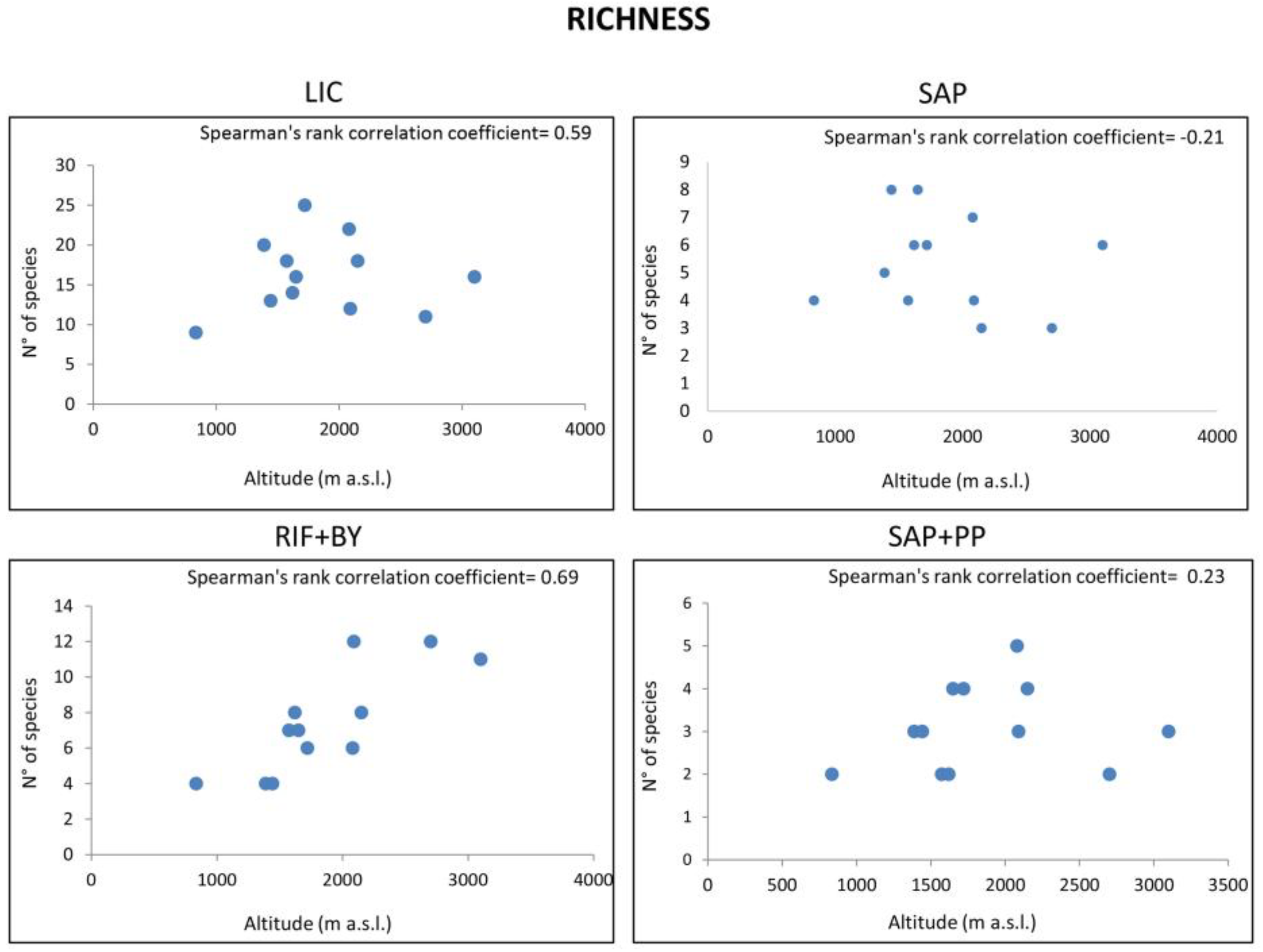
3.2. Diversity Measures
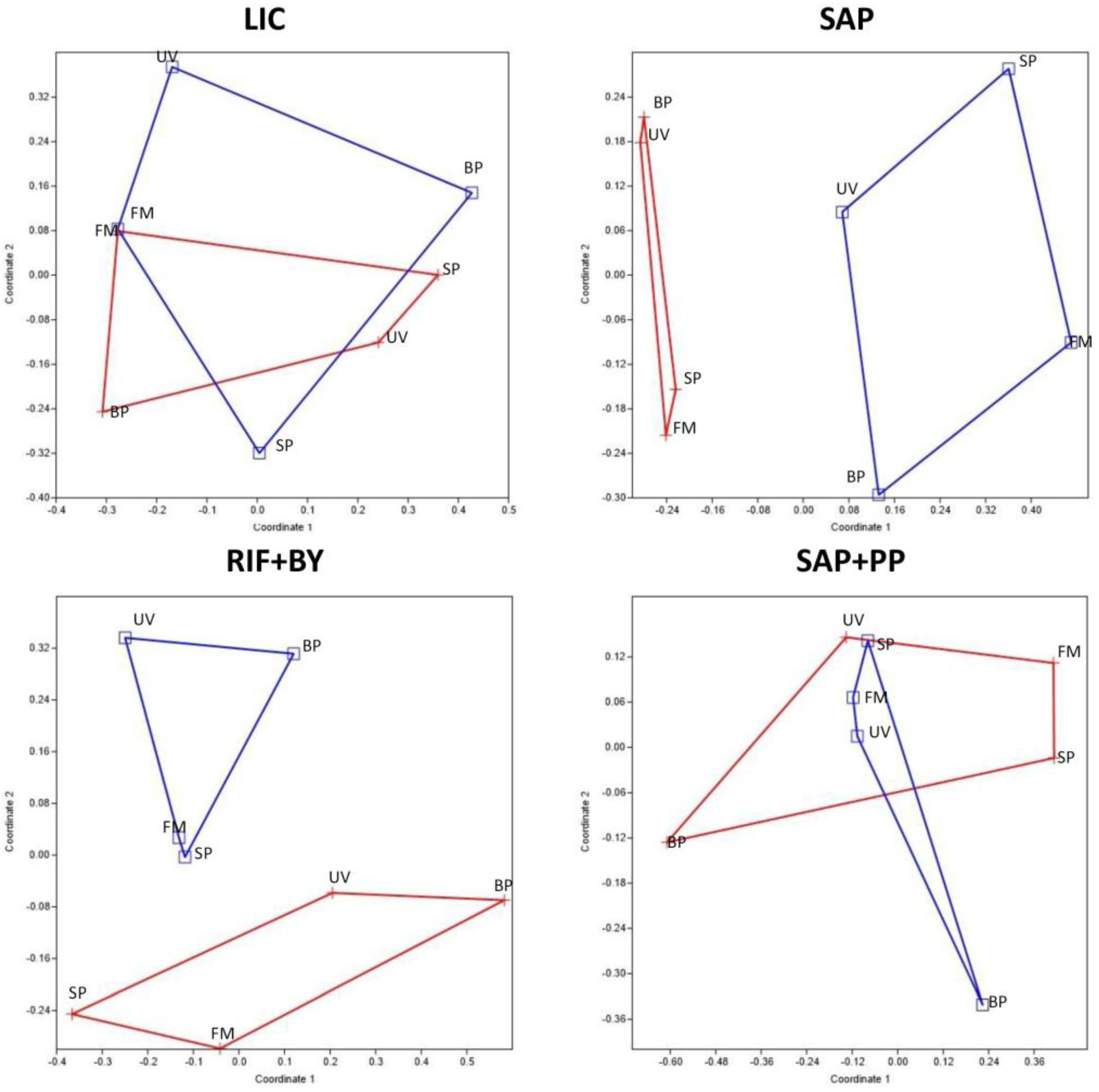
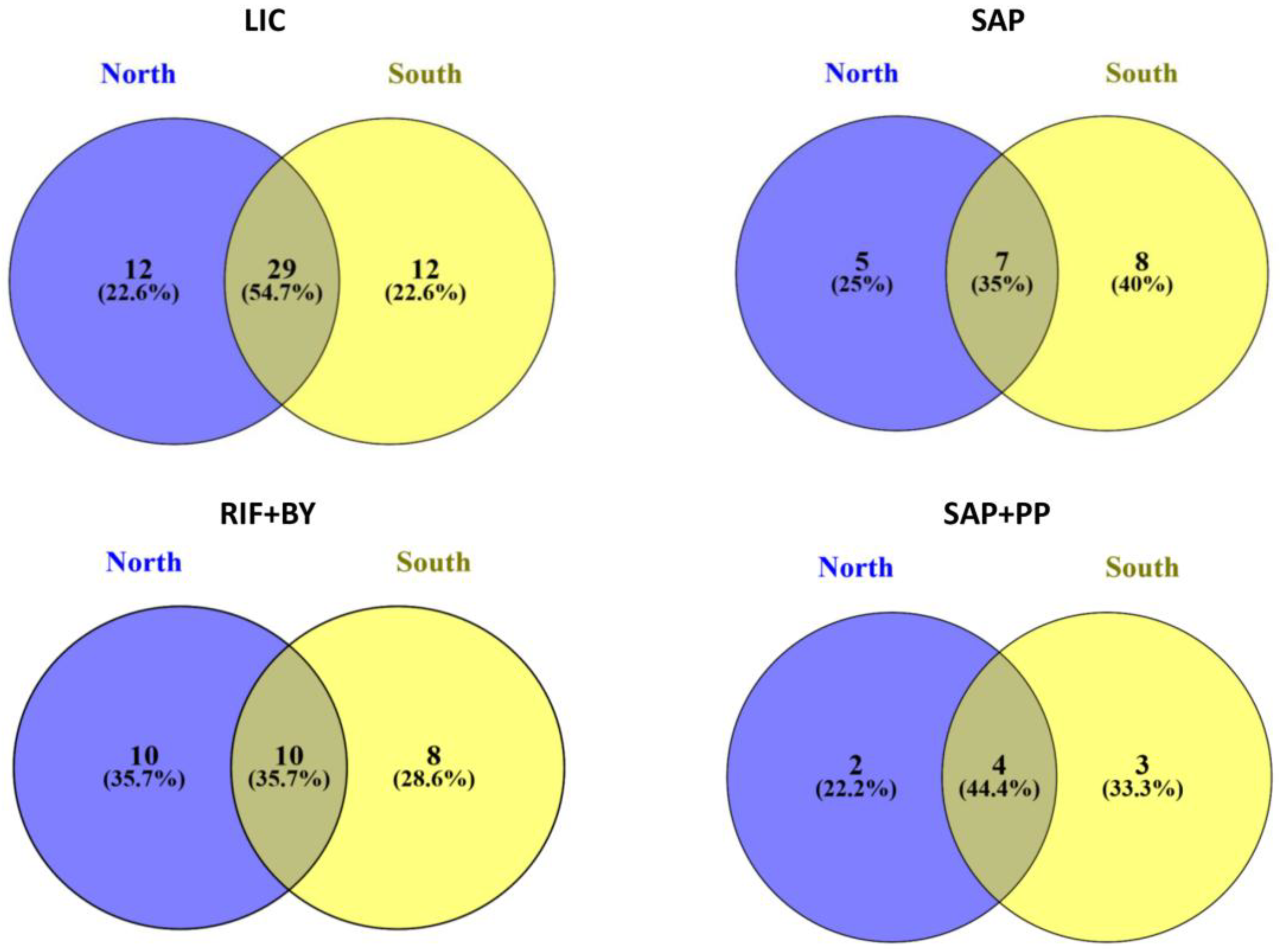
3.3. Community Composition
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Horowitz, N.H.; Cameron, R.E.; Hubbard, J.S. Microbiology of the dry valleys of Antarctica. Science 1972, 176, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Pointing, S.B.; Chan, Y.; Lacap, D.C.; Lau, M.C.; Jurgens, J.A. Highly specialized microbial diversity in hyper arid polar desert. Proc. Natl. Acad. Sci. USA 2009, 106, 19964–19969. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Chan, Y.; Lacap, D.C.; Hyde, K.D.; Pointing, S.B.; Farrell, R.L. Low-diversity fungal assemblage in an Antarctic Dry Valleys soil. Polar Biol. 2011, 35, 567–574. [Google Scholar] [CrossRef]
- Lee, C.K.; Barbier, B.A.; Bottos, E.M.; McDonald, I.R.; Cary, S.C. The inter-valley soil comparative survey: The ecology of Dry Valley edaphic microbial communities. ISME J. 2012, 6, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Cowan, D.; Tow, L. Endangered Antarctic environments. Annu. Rev. Microbiol. 2004, 58, 649–690. [Google Scholar] [CrossRef] [PubMed]
- Cary, S.C.; McDonald, I.R.; Barrett, J.E.; Cowan, D.A. On the rocks: The microbiology of Antarctic Dry Valley soils. Nat. Rev. Microbiol. 2010, 8, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Cowan, D.; Makhalanyane, T.P.; Dennis, P.G.; Hopkins, D.W. Microbial ecology and biogeochemistry of continental Antarctic soils. Front. Microbiol. 2014, 5, 154. [Google Scholar] [CrossRef] [PubMed]
- Friedmann, E.I. Endolithic microorganisms in the Antarctic cold desert. Science 1982, 215, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Nienow, J.A.; Friedmann, E.I. Terrestrial lithophytic (rock) communities. In Antarctic Microbiology; Friedmann, E.I., Ed.; Wiley-Liss: New York, NY, USA, 1993; pp. 343–412. [Google Scholar]
- Onofri, S.; Zucconi, L.; Selbmann, L.; de Hoog, S.; de los Rios, A.; Ruisi, S.; Grube, M. Fungal associations at the cold edge of life. In Algae and Cyanobacteria in Extreme Environments; Seckbach, J., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 735–757. [Google Scholar]
- Zucconi, L.; Onofri, S.; Cecchini, C.; Isola, D.; Ripa, C.; Fenice, M.; Madonna, S.; Reboleiro-Rivas, P.; Selbmann, L. Mapping the lithic colonization at the boundaries of life in Northern Victoria Land, Antarctica. Polar Biol. 2016, 39, 91–102. [Google Scholar] [CrossRef]
- Selbmann, L.; de Hoog, G.S.; Mazzaglia, A.; Friedmann, E.I.; Onofri, S. Fungi at the edge of life: Cryptoendolithic black fungi from Antarctic deserts. Stud. Mycol. 2005, 51, 1–32. [Google Scholar]
- Selbmann, L.; de Hoog, G.S.; Zucconi, L.; Isola, D.; Ruisi, S.; Gerrits van den Ende, A.H.G.; Ruibal, C.; De Leo, F.; Urzi, C.; Onofri, O. Drought meets acid: Three new genera in a dothidealean clade of extremotolerant fungi. Stud. Mycol. 2008, 61, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; Onofri, S.; Coleine, C.; Buzzini, P.; Canini, F.; Zucconi, L. Effect of environmental parameters on biodiversity of the fungal component in the lithic Antarctic communities. Extremophiles 2017, 21, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Coleine, C.; Stajich, J.E.; Zucconi, L.; Onofri, S.; Pombubpa, N.; Egidi, E.; Franks, A.E.; Buzzini, P.; Selbmann, L. Antarctic cryptoendolithic fungal communities are highly adapted and dominated by Lecanoromycetes and Dothideomycetes. Front. Microbiol. 2018. [Google Scholar]
- Selbmann, L.; Isola, D.; Fenice, F.; Zucconi, L.; Sterflinger, K.; Onofri, S. Potential extinction of Antarctic endemic fungal species as a consequence of global warming. Sci. Total Environ. 2012, 438, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change. In Proceedings of the 22nd Session of the IPCC, New Delhi, India, 9–11 November 2004.
- Steig, E.J.; Schneider, D.P.; Rutherford, S.D.; Mann, M.E.; Comiso, J.C.; Shindell, D.T. Warming of the Antarctic ice-sheet surface since the 1957 international geophysical year. Nature 2009, 457, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Hogg, I.D.; Wall, D.H. Global change and Antarctic terrestrial biodiversity. Polar Biol. 2011, 34, 1625–1627. [Google Scholar] [CrossRef]
- Grau, O.; Geml, J.; Pérez-Haase, A.; Ninot, J.M.; Semenova-Nelsen, T.A.; Peñuelas, J. Abrupt changes in the composition and function of fungal communities along an environmental gradient in the High Arctic. Mol. Ecol. 2017, 26, 4798–4810. [Google Scholar] [CrossRef] [PubMed]
- Timling, I.; Walker, D.A.; Nusbaum, C.; Lennon, N.J.; Taylor, D.L. Rich and cold: Diversity, distribution and drivers of fungal communities in patterned-ground ecosystems of the North American Arctic. Mol. Ecol. 2014, 23, 3258–3272. [Google Scholar] [CrossRef] [PubMed]
- Geml, J.; Morgado, L.N.; Semenova, T.A.; Welker, J.M.; Walker, M.D.; Smets, E. Long-term warming alters richness and composition of taxonomic and functional groups of arctic fungi. FEMS Microbiol. Ecol. 2015, 91, fiv095. [Google Scholar] [CrossRef] [PubMed]
- Geml, J.; Semenova, T.A.; Morgado, L.N.; Welker, J.M. Changes in composition and abundance of functional groups of arctic fungi in response to long-term summer warming. Biol. Lett. 2016, 12, 20160503. [Google Scholar] [CrossRef] [PubMed]
- Semenova, T.A.; Morgado, L.N.; Welker, J.M.; Walker, M.D.; Smets, E.; Geml, J. Compositional and functional shifts in arctic fungal communities in response to experimentally increased snow depth. Soil Biol. Biochem. 2016, 100, 201–209. [Google Scholar] [CrossRef]
- Smith, D.P.; Peay, K.G. Sequence depth, not PCR replication, improves ecological inference from next generation DNA sequencing. PLoS ONE 2014, 9, e90234. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Palmer, J.M.; Jusino, M.A.; Banik, M.T.; Lindner, D.L. Non-biological synthetic spike-in controls and the AMPtk software pipeline improve fungal high throughput amplicon sequencing data. BioRxiv 2017, 213470. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers—A user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, H.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.M.; Vilgalys, R. Fungal community analysis by large-scale sequencing of environmental samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef] [PubMed]
- Geml, J.; Laursen, G.A.; Timling, I.; Mcfarland, J.M.; Booth, M.G.; Lennon, N.; Nusbaum, C.; Taylor, D.L. Molecular phylogenetic biodiversity assessment of arctic and boreal Lactarius Pers. (Russulales; Basidiomycota) in Alaska, based on soil and sporocarp DNA. Mol. Ecol. 2009, 18, 2213–2227. [Google Scholar] [CrossRef]
- Geml, J.; Laursen, G.A.; Herriott, I.C.; McFarland, J.M.; Booth, M.G.; Lennon, N.; Nusbaum, C.; Taylor, D.L. Phylogenetic and ecological analyses of soil and sporocarp DNA sequences reveal high diversity and strong habitat partitioning in the boreal ectomycorrhizal genus Russula (Russulales; Basidiomycota). New Phytol. 2010, 187, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Kauserud, H.; Kumar, S.; Brysting, A.K.; Nordén, J.; Carlsen, T. High consistency between replicate 454 pyrosequencing analyses of ectomycorrhizal plant root samples. Mycorrhiza 2012, 22, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.P.; Callaham, M.A.; Oliver, A.K.; Jumpponen, A. Deep Ion Torrent sequencing identifies soil fungal community shifts after frequent prescribed fires in a southeastern US forest ecosystem. FEMS Microbiol. Ecol. 2013, 86, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. Peer J. 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Oliveros, J.C. VENNY. An Interactive Tool for Comparing Lists with Venn Diagrams. 2007. Available online: http://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 20 November 2013).
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Spearman, C. The proof and measurement of association between two things. Am. J. Psychol. 1904, 15, 7–101. [Google Scholar] [CrossRef]
- Amend, A.S.; Seifert, K.A.; Bruns, T.D. Quantifying microbial communities with 454 pyrosequencing: Does read abundance count? Mol. Ecol. 2010, 19, 5555–5565. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Vetrovsky, T.; Cajthaml, T.; Dobiášová, P.; Petránková, M.; Šnajdr, J.; Eichlerová, I. Estimation of fungal biomass in forest litter and soil. Fungal Ecol. 2013, 6, 1–11. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST-Palaeontological Statistics. 2001. Available online: www. uv. es/~pardomv/pe/2001_1/past/pastprog/past.pdf (accessed on 29 May 2018).
- De la Torre, J.R.; Goebel, B.M.; Friedmann, E.; Pace, N.R. Microbial diversity of cryptoendolithic communities from the McMurdo Dry Valleys. Antarctica. Appl. Environ. Microbiol. 2003, 69, 3858–3867. [Google Scholar] [CrossRef] [PubMed]
- Archer, S.D.; de los Ríos, A.; Lee, K.C.; Niederberger, T.S.; Cary, S.C.; Coyne, K.J.; Douglas, S.; Lacap-Bugler, D.C.; Pointing, S.B. Endolithic microbial diversity in sandstone and granite from the McMurdo Dry Valleys, Antarctica. Polar Biol. 2017, 40, 997–1006. [Google Scholar] [CrossRef]
- Kappen, L. Some aspects of the great success of lichens in Antarctica. Antarct. Sci. 2000, 12, 314–324. [Google Scholar] [CrossRef]
- Cockell, C.S.; McKay, C.P.; Omelon, C. Polar endoliths—An anti-correlation of climatic extremes and microbial biodiversity. Int. J. Astrobiol. 2003, 1, 305–310. [Google Scholar] [CrossRef]
- De los Ríos, A.; Sancho, L.G.; Grube, M.; Wierzchos, J.; Ascaso, C. Endolithic growth of two Lecidea lichens in granite from continental Antarctica detected by molecular and microscopy techniques. New Phytol. 2005, 165, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.C.; Hogg, I.D.; Wilkins, R.J.; Green, T.G.A. Microsatellite analyses of the Antarctic endemic lichen Buellia frigida Darb. (Physciaceae) suggest limited dispersal and the presence of glacial refugia in the Ross Sea region. Polar Biol. 2015, 38, 941–949. [Google Scholar] [CrossRef]
- Liu, X.Z.; Wang, Q.M.; Göker, M.; Groenewald, M.; Kachalkin, A.V.; Lumbsch, H.T.; Millanes, A.M.; Wedin, M.; Yurkov, A.M.; Boekhout, T.; et al. Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef] [PubMed]
- Vishniac, H.S.; Hempfling, W.P. Cryptococcus vishniacii sp. nov., an Antarctic yeast. Int. J. Syst. Evol. Microbiol. 1979, 29, 153–158. [Google Scholar] [CrossRef]
- Vishniac, H.S. Cryptococcus friedmannii, a new species of yeast from the Antarctic. Mycologia 1985, 77, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Vishniac, H.S.; Kurtzman, C.P. Cryptococcus antarcticus sp. nov. and Cryptococcus albidosimilis sp. nov., basidioblastomycetes from Antarctic soils. Int. J. Syst. Evol. Microbiol. 1992, 42, 547–553. [Google Scholar] [CrossRef]
- Selbmann, L.; Zucconi, L.; Onofri, S.; Cecchini, C.; Isola, D.; Turchetti, B.; Buzzini, P. Taxonomic and phenotypic characterization of yeasts isolated from worldwide cold rock-associated habitats. Fungal Biol. 2014, 118, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Turchetti, B.; Selbmann, L.; Blanchette, R.A.; Di Mauro, S.; Marchegiani, E.; Zucconi, L.; Arenz, B.E.; Buzzini, P. Cryptococcus vaughanmartiniae sp. nov. and Cryptococcus onofrii sp. nov.: Two new species isolated from worldwide cold environments. Extremophiles 2015, 19, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; Turchetti, B.; Yurkov, A.; Cecchini, C.; Zucconi, L.; Isola, D.; Buzzini, P.; Onofri, S. Description of Taphrina antarctica fa sp. nov., a new anamorphic ascomycetous yeast species associated with Antarctic endolithic microbial communities and transfer of four Lalaria species in the genus Taphrina. Extremophiles 2014, 18, 707–721. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; Egidi, E.; Isola, D.; Onofri, S.; Zucconi, L.; de Hoog, G.S.; Chinaglia, S.; Testa, L.; Tosi, S.; Balestrazzi, A.; et al. Biodiversity, evolution and adaptation of fungi in extreme environments. Plant Biosyst. 2013, 147, 237–246. [Google Scholar] [CrossRef]
- Selbmann, L.; de Hoog, G.S.; Zucconi, L.; Isola, D.; Onofri, S. Black yeasts in cold habitats. In Cold-Adapted Yeasts; Springer: Berlin/Heidelberg, Germany, 2014; pp. 173–189. [Google Scholar]
- Dadachova, E.; Casadevall, A. Ionizing radiation: How fungi cope, adapt, and exploit with the help of melanin. Curr. Opin. Microbiol. 2008, 11, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; Barreca, D.; Selbmann, L.; Isola, D.; Rabbow, E.; Horneck, G.; de Vera, J.P.; Hatton, J.; Zucconi, L. Resistance of Antarctic black fungi and cryptoendolithic communities to simulated space and Martian conditions. Stud. Mycol. 2008, 61, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; de la Torre, R.; de Vera, J.P.; Ott, S.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, J.K.; Rabbow, E.; Sánchez Iñigo, F.J.; et al. Survival of rock-colonizing organisms after 1.5 Years in outer space. Astrobiology 2012, 12, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; de Vera, J.P.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, K.J.; Elke, R.; de la Torre, R.; Gerda, H. Survival of Antarctic cryptoendolithic fungi in simulated Martian conditions on-board the International Space Station. Astrobiology 2015, 15, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; Isola, D.; Zucconi, L.; Onofri, S. Resistance to UV-B induced DNA damage in extreme-tolerant cryptoendolithic Antarctic fungi: Detection by PCR assays. Fungal Biol. 2011, 115, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; Pacelli, C.; Zucconi, L.; Dadachova, E.; Moeller, R.; de Vera, J.P.; Onofri, S. Resistance of an Antarctic cryptoendolithic black fungus to radiation gives new insights of astrobiological relevance. Fungal Biol. 2017, 122, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Pacelli, C.; Bryan, R.A.; Onofri, S.; Selbmann, L.; Shuryak, I.; Dadachova, E. Melanin is effective in protecting fast and slow growing fungi from various types of ionizing radiation. Environ. Microbiol. 2017, 19, 1612–1624. [Google Scholar] [CrossRef] [PubMed]
- Selbmann, L.; Zucconi, L.; Isola, D.; Onofri, S. Rock black fungi: Excellence in the extremes, from the Antarctic to space. Curr. Genet. 2015, 61, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Friedmann, E.I.; McKay, C.P.; Nienow, J.A. The cryptoendolithic microbial environment in the Ross Desert of Antarctica: Satellite-transmitted continuous nanoclimate data, 1984 to 1986. Polar Biol. 1987, 7, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Sonjak, S.; Zalar, P.; Frisvad, J.C.; Diderichsen, B.; Plemenitaš, A. Extremophilic fungi in arctic ice: A relationship between adaptation to low temperature and water activity. Phys. Chem. Earth Parts A/B/C 2003, 28, 1273–1278. [Google Scholar] [CrossRef]
- Sterflinger, K.; Tesei, D.; Zakharova, K. Fungi in hot and cold deserts with particular reference to microcolonial fungi. Fungal Ecol. 2012, 5, 453–462. [Google Scholar] [CrossRef]
- Rice, A.V.; Currah, R.S. Two new species of Pseudogymnoascus with Geomyces anamorphs and their phylogenetic relationship with Gymnostellatospora. Mycologia 2006, 98, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S.; Bush, T.; Bryce, C.; Direito, S.; Fox-Powell, M.; Harrison, J.P.; Lammer, H.; Landenmark, H.; Martin-Torres, J.; Nicholson, N.; et al. Habitability: A review. Astrobiology 2016, 16, 89–117. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Altitude (m a.s.l.) | Air Temperature (°C) | Humidity (%) | Coordinates |
|---|---|---|---|---|
| Battleship Promontory | 834 | −4.4 | 22.9 | 76°54′04.0′′ S 160°54′36.6′′ E |
| Trio Nunatak | 1388 | −5.1 | 40.9 | 75°28′56.6′′ S 159°35′28.3′′ E |
| Ricker Hills | 1442 | −7.2 | 42.7 | 75°42′14.6′′ S 159°13′39.4′′ E |
| Pudding Butte | 1573 | −8.5 | 32.4 | 75°51′30.2′′ S 159°58′25.7′′ E |
| Siegfried Peak | 1620 | −9.3 | 52.8 | 77°34′43.3′′ S 161°47′11.7′′ E |
| Linnaeus Terrace | 1649 | −9.6 | 58.6 | 77°36′01.3′′ S 161°05′00.5′′ E |
| Finger Mt. | 1720 | −6.4 | 35.1 | 77°54′43.6′′ S 161°34′39.3′′ E |
| Mt. Elektra | 2080 | −11.9 | 63 | 77°29′28.0′′ S 160°54′16.4′′ E |
| University Valley | 2090 | −14.3 | 18 | 77°52′28.6′′ S 160°44′22.6′′ E |
| Knobhead | 2150 | −12.5 | 50 | 77°54′37.8′′ S 161°34′48.8′′ E |
| Timber Peak | 2702 | −12.4 | 30.1 | 74°10′10.5′′ S 162°25′38.0′′ E |
| Mt New Zealand | 3100 | −17.2 | 47.6 | 74°10′44.0′′ S 162°30′53.0′′ E |
| N° of OTUs | Dataset 1 | Dataset 2 |
|---|---|---|
| LIC | 36 | 53 |
| SAP | 19 | 20 |
| RIF + BY | 24 | 28 |
| SAP + PP | 10 | 9 |
| Index | Altitude (m) | LIC | SAP | RIF + BY | SAP + PP | Unidentified OTUs |
|---|---|---|---|---|---|---|
| Richness (S) | 834 | 9 | 4 | 4 | 2 | 13 |
| 1388 | 20 | 5 | 4 | 3 | 15 | |
| 1442 | 13 | 8 | 4 | 3 | 11 | |
| 1573 | 18 | 4 | 7 | 2 | 12 | |
| 1620 | 14 | 6 | 8 | 3 | 15 | |
| 1649 | 16 | 8 | 7 | 4 | 13 | |
| 1720 | 25 | 6 | 6 | 4 | 14 | |
| 2080 | 22 | 7 | 6 | 5 | 15 | |
| 2090 | 12 | 4 | 12 | 3 | 11 | |
| 2150 | 18 | 3 | 8 | 4 | 9 | |
| 2702 | 11 | 3 | 12 | 2 | 11 | |
| 3100 | 16 | 6 | 11 | 3 | 8 | |
| Mean Value | 16 | 5 | 7 | 3 | 12 | |
| Shannon’s Diversity (H’) | 834 | 1.95 | 1.23 | 1.31 | 0.68 | |
| 1388 | 2.78 | 1.2 | 1.24 | 0.76 | ||
| 1442 | 2.36 | 1.35 | 1.11 | 0.35 | ||
| 1573 | 2.77 | 0.98 | 2.28 | 0.58 | ||
| 1620 | 2.43 | 1.36 | 1.41 | 0.23 | ||
| 1649 | 2.55 | 1.84 | 1.64 | 1.07 | ||
| 1720 | 3.06 | 1.1 | 1.76 | 1.04 | ||
| 2080 | 2.84 | 1.31 | 2.06 | 1.52 | ||
| 2090 | 2.27 | 1.06 | 1.28 | 1.07 | ||
| 2150 | 2.73 | 0.85 | 2.34 | 1.19 | ||
| 2702 | 2.20 | 0.83 | 1.48 | 0.4 | ||
| 3100 | 2.56 | 1.38 | 2.42 | 0.93 | ||
| Mean Value | 2.54 | 1.21 | 1.69 | 0.81 | ||
| Simpson’s Dominance (1-D) | 834 | 0.86 | 0.98 | 0.83 | 0.57 | |
| 1388 | 0.94 | 0.71 | 0.77 | 0.60 | ||
| 1442 | 0.91 | 0.74 | 0.73 | 0.58 | ||
| 1573 | 0.94 | 0.71 | 0.92 | 0.51 | ||
| 1620 | 0.91 | 0.79 | 0.72 | 0.58 | ||
| 1649 | 0.92 | 0.88 | 0.84 | 0.64 | ||
| 1720 | 0.96 | 0.57 | 0.80 | 0.65 | ||
| 2080 | 0.94 | 0.73 | 0.90 | 0.86 | ||
| 2090 | 0.90 | 0.71 | 0.70 | 0.71 | ||
| 2150 | 0.93 | 0.61 | 0.90 | 0.75 | ||
| 2702 | 0.89 | 0.57 | 0.85 | 0.75 | ||
| 3100 | 0.92 | 0.78 | 0.94 | 0.62 | ||
| mean value | 0.92 | 0.73 | 0.86 | 0.65 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coleine, C.; Zucconi, L.; Onofri, S.; Pombubpa, N.; Stajich, J.E.; Selbmann, L. Sun Exposure Shapes Functional Grouping of Fungi in Cryptoendolithic Antarctic Communities. Life 2018, 8, 19. https://doi.org/10.3390/life8020019
Coleine C, Zucconi L, Onofri S, Pombubpa N, Stajich JE, Selbmann L. Sun Exposure Shapes Functional Grouping of Fungi in Cryptoendolithic Antarctic Communities. Life. 2018; 8(2):19. https://doi.org/10.3390/life8020019
Chicago/Turabian StyleColeine, Claudia, Laura Zucconi, Silvano Onofri, Nuttapon Pombubpa, Jason E. Stajich, and Laura Selbmann. 2018. "Sun Exposure Shapes Functional Grouping of Fungi in Cryptoendolithic Antarctic Communities" Life 8, no. 2: 19. https://doi.org/10.3390/life8020019
APA StyleColeine, C., Zucconi, L., Onofri, S., Pombubpa, N., Stajich, J. E., & Selbmann, L. (2018). Sun Exposure Shapes Functional Grouping of Fungi in Cryptoendolithic Antarctic Communities. Life, 8(2), 19. https://doi.org/10.3390/life8020019