
Senescence-Associated β-Galactosidase Detection in Pathology
 ,
,  ,
,
Abstract
:1. Introduction
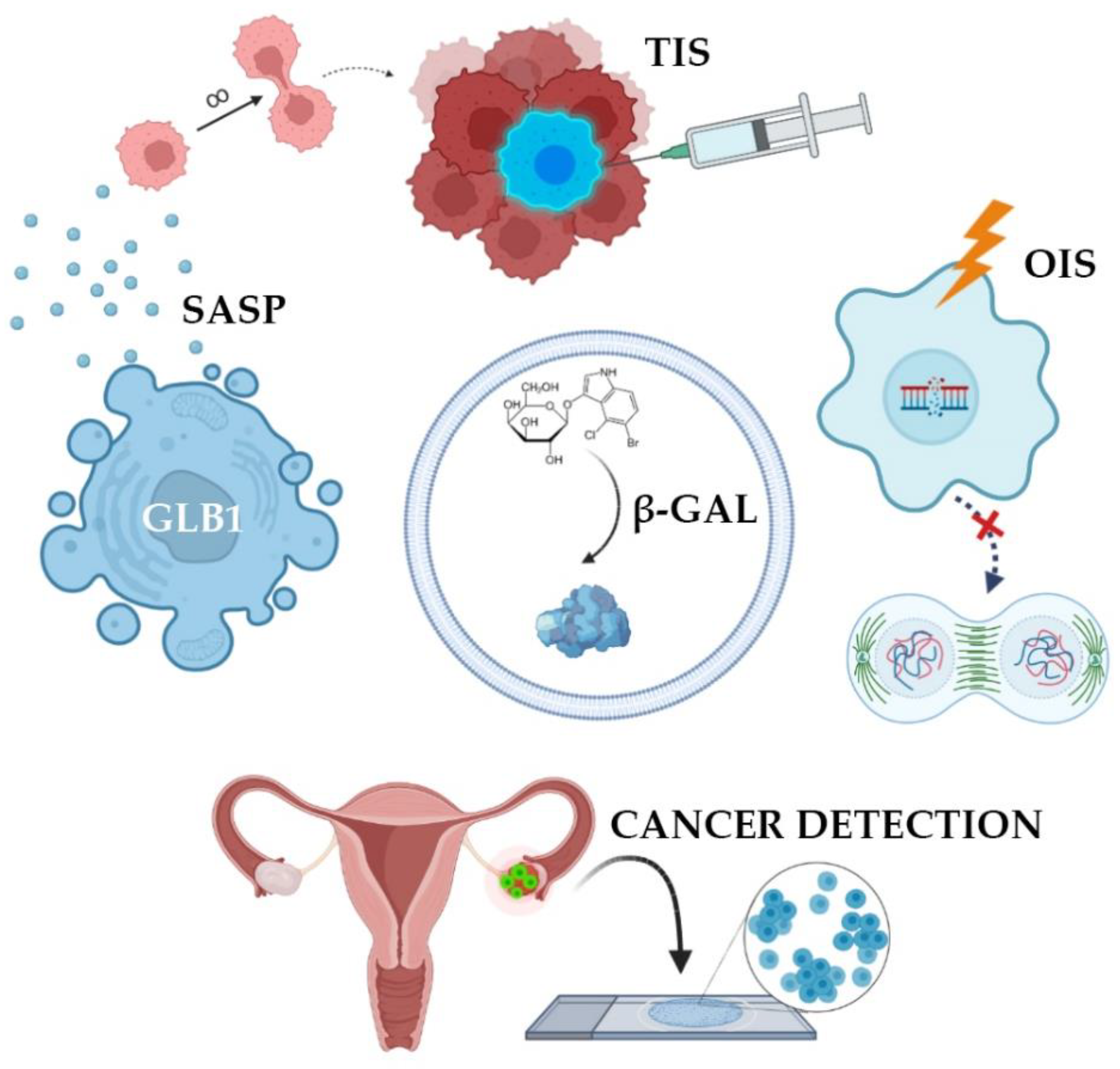
2. Medical Applications of SA-β-Gal Assay
2.1. Mechanism of β-Galactosidase Activation
2.2. Senescence-Associated Secretory Phenotype
2.3. β-Galactosidase Dependent Therapy
2.4. SA-β-Gal Analysis in Oncopathology
2.5. SA-β-Gal Detection
2.6. SA-β-Gal as a Prognostic Marker
2.7. Chemotherapy and Cellular Senescence
2.8. SA-β-Gal in Non-Tumor Lesions
2.9. SA-β-Gal in Fibrosis
3. Future Prospects
- (1)
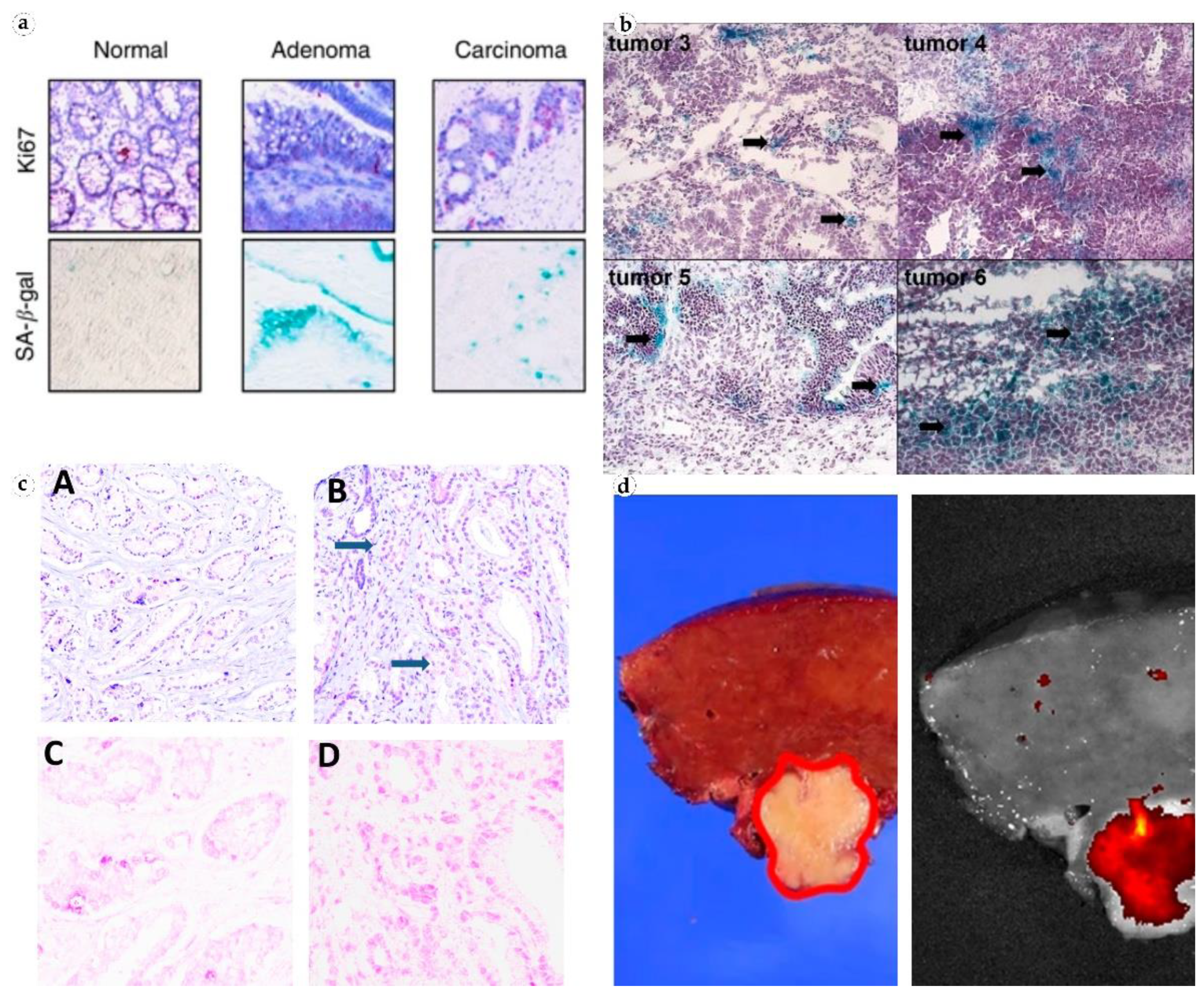
- SA-β-gal accumulation in tumors is non-linear, which can be used to evaluate the progression of precancerous and cancerous lesions. Some studies reported that the expression of SA-β-gal strongly correlated with an increase in the malignant potential of a cell: The highest level of SA-β-gal was detected in carcinomas, the average level was in precancerous conditions and the lowest level was in benign tumors [26,31,45]. However, it is likely that high expression of β-galactosidase is temporary and reaches its peak in obligate precancers or in recent carcinomas [31,43]. Prolonged clinical studies are needed to shed light on the dynamics and possible impact of cellular senescence on tumor growth.
- (2)
- SA-β-gal expression differs among cancer cells. It can be explained by the heterogenous structure of tumors and functional distinctions between primary tumor cells and metastases. Although SA-β-gal was used to visualize ovarian and gastric cancer metastases by bioluminescence imaging [42,50], the enzyme reached its highest concentration in solid non-invasive tumors [21]. This can be used to better evaluate the risk of tumor progression, i.e., after surgical resection.
- (3)
- SA-β-gal content significantly increases after neoadjuvant therapy [30,37]. This can be used to assess patient’s response to the treatment and correct therapeutic strategy. Targeting SA-β-gal for the delivery of senolytic drugs has a chance to become a mainstream direction in the treatment of tumors or inflammatory processes. Cellular senescence may explain the persistence of some tumor cells and may become important in predicting response to therapy. The exploration of SA-β-gal activity in the development of antifibrotic drugs for hepatic fibrosis, hypertrophic scars, and other conditions is also very promising.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krishna, D.R.; Sperker, B.; Fritz, P.; Klotz, U. Does pH 6 β-galactosidase activity indicate cell senescence? Mech. Ageing Dev. 1999, 109, 113–123. [Google Scholar] [CrossRef]
- Morgunova, G.V.; Kolesnikov, A.V.; Klebanov, A.A.; Khokhlov, A.N. Is aging-associated betagalactosidase a biomarker of aging, DNA damage or cell proliferation limitation? Bull. Mosc. Univ. 2015, 16, 15–18. [Google Scholar]
- Li, M.; Yang, M.; Zhu, W.-H. Advances in fluorescent sensors for β-galactosidase. Mater. Chem. Front. 2021, 5, 763–774. [Google Scholar] [CrossRef]
- Okamura-Oho, Y.; Zhang, S.; Callahan, J.W.; Murata, M.; Oshima, A.; Suzuki, Y. Maturation and degradation of beta-galactosidase in the post-Golgi compartment are regulated by cathepsin B and a non-cysteine protease. FEBS Lett. 1997, 419, 231–234. [Google Scholar] [CrossRef]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; LINskENs, M.; Rubelj, I.; Pereira-Smith, O. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef]
- Lee, B.Y.; Han, J.A.; Im, J.S.; Morrone, A.; Johung, K.; Goodwin, E.C.; Kleijer, W.J.; DiMaio, D.; Hwang, E.S. Senescence-associated β-galactosidase is lysosomal β-galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef]
- Kurz, D.J.; Decary, S.; Hong, Y.; Erusalimsky, J.D. Senescence-associated (beta)-galactosidase reflects an increase in lysosomal mass during replicative ageing of human endothelial cells. J. Cell Sci. 2000, 113 Pt 20, 3613–3622. [Google Scholar] [CrossRef]
- Gerland, L.M.; Peyrol, S.; Lallemand, C.; Branche, R.; Magaud, J.P.; Ffrench, M. Association of increased autophagic inclusions labeled for beta-galactosidase with fibroblastic aging. Exp. Gerontol. 2003, 38, 887–895. [Google Scholar] [CrossRef]
- Abe, A.; Shayman, J.A. Sphingolipid Catabolism. In Encyclopedia of Biological Chemistry; Academic Press: Cambridge, MA, USA, 2013; pp. 287–292. [Google Scholar]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to detect senescence-associated beta-galactosidase (SA-betagal) activity, a biomarker of senescent cells in culture and in vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef]
- Burn, S.F. Detection of β-galactosidase activity: X-gal staining. In Kidney Development; Springer: Berlin/Heidelberg, Germany, 2012; pp. 241–250. [Google Scholar]
- Gu, K.; Xu, Y.; Li, H.; Guo, Z.; Zhu, S.; Zhu, S.; Shi, P.; James, T.D.; Tian, H.; Zhu, W.H. Real-Time Tracking and In Vivo Visualization of beta-Galactosidase Activity in Colorectal Tumor with a Ratiometric Near-Infrared Fluorescent Probe. J. Am. Chem. Soc. 2016, 138, 5334–5340. [Google Scholar] [CrossRef]
- Wang, L.; Lankhorst, L.; Bernards, R. Exploiting senescence for the treatment of cancer. Nat. Rev. Cancer 2022, 22, 340–355. [Google Scholar] [CrossRef]
- Calcinotto, A.; Kohli, J.; Zagato, E.; Pellegrini, L.; Demaria, M.; Alimonti, A. Cellular Senescence: Aging, Cancer, and Injury. Physiol. Rev. 2019, 99, 1047–1078. [Google Scholar] [CrossRef]
- Chen, S.; Wang, L.; Ma, X.; Wu, Y.; Hou, S. Kill two birds with one stone: A near-infrared ratiometric fluorescent probe for simultaneous detection of β-galactosidase in senescent and cancer cells. Sens. Actuators B Chem. 2022, 367, 132061. [Google Scholar] [CrossRef]
- Zieba, J.; Ksiazkiewcz, M.; Janik, K.; Banaszczyk, M.; Peciak, J.; Piaskowski, S.; Lipinski, M.; Olczak, M.; Stoczynska-Fidelus, E.; Rieske, P. Sensitivity of neoplastic cells to senescence unveiled under standard cell culture conditions. Anticancer Res. 2015, 35, 2759–2768. [Google Scholar]
- Gorgoulis, V.G.; Halazonetis, T.D. Oncogene-induced senescence: The bright and dark side of the response. Curr. Opin. Cell Biol. 2010, 22, 816–827. [Google Scholar] [CrossRef]
- Zeng, S.; Shen, W.H.; Liu, L. Senescence and Cancer. Cancer Transl. Med. 2018, 4, 70–74. [Google Scholar] [CrossRef]
- Haigis, K.M.; Sweet-Cordero, A. New insights into oncogenic stress. Nat. Genet. 2011, 43, 177–178. [Google Scholar] [CrossRef]
- Kaplon, J.; Zheng, L.; Meissl, K.; Chaneton, B.; Selivanov, V.A.; Mackay, G.; van der Burg, S.H.; Verdegaal, E.M.; Cascante, M.; Shlomi, T.; et al. A key role for mitochondrial gatekeeper pyruvate dehydrogenase in oncogene-induced senescence. Nature 2013, 498, 109–112. [Google Scholar] [CrossRef]
- Wielgat, P.; Walczuk, U.; Szajda, S.; Bien, M.; Zimnoch, L.; Mariak, Z.; Zwierz, K. Activity of lysosomal exoglycosidases in human gliomas. J. Neurooncol. 2006, 80, 243–249. [Google Scholar] [CrossRef]
- Ewald, J.A.; Desotelle, J.A.; Wilding, G.; Jarrard, D.F. Therapy-induced senescence in cancer. J. Natl. Cancer Inst. 2010, 102, 1536–1546. [Google Scholar] [CrossRef]
- Cai, Y.; Zhou, H.; Zhu, Y.; Sun, Q.; Ji, Y.; Xue, A.; Wang, Y.; Chen, W.; Yu, X.; Wang, L.; et al. Elimination of senescent cells by beta-galactosidase-targeted prodrug attenuates inflammation and restores physical function in aged mice. Cell Res. 2020, 30, 574–589. [Google Scholar] [CrossRef]
- Pakula, M.; Maly, E.; Uruski, P.; Witucka, A.; Bogucka, M.; Jaroszewska, N.; Makowska, N.; Niklas, A.; Moszynski, R.; Sajdak, S.; et al. Deciphering the Molecular Mechanism of Spontaneous Senescence in Primary Epithelial Ovarian Cancer Cells. Cancers 2020, 12, 296. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.K.; Bhattacharya, M.; Barlow, J.J. Glycosyltransferase and glycosidase activities in ovarian cancer patients. Cancer Res. 1979, 39, 1943–1951. [Google Scholar]
- Bosmann, H.B.; Hall, T.C. Enzyme activity in invasive tumors of human breast and colon. Proc. Natl. Acad. Sci. USA 1974, 71, 1833–1837. [Google Scholar] [CrossRef]
- Haugstetter, A.M.; Loddenkemper, C.; Lenze, D.; Grone, J.; Standfuss, C.; Petersen, I.; Dorken, B.; Schmitt, C.A. Cellular senescence predicts treatment outcome in metastasised colorectal cancer. Br. J. Cancer 2010, 103, 505–509. [Google Scholar] [CrossRef]
- Bartkova, J.; Rezaei, N.; Liontos, M.; Karakaidos, P.; Kletsas, D.; Issaeva, N.; Vassiliou, L.V.; Kolettas, E.; Niforou, K.; Zoumpourlis, V.C.; et al. Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints. Nature 2006, 444, 633–637. [Google Scholar] [CrossRef]
- Charalambous, C.; Virrey, J.; Kardosh, A.; Jabbour, M.N.; Qazi-Abdullah, L.; Pen, L.; Zidovetzki, R.; Schönthal, A.H.; Chen, T.C.; Hofman, F.M. Glioma-associated endothelial cells show evidence of replicative senescence. Exp. Cell Res. 2007, 313, 1192–1202. [Google Scholar] [CrossRef]
- te Poele, R.H.; Okorokov, A.L.; Jardine, L.; Cummings, J.; Joel, S.P. DNA damage is able to induce senescence in tumor cells in vitro and in vivo. Cancer Res. 2002, 62, 1876–1883. [Google Scholar]
- Wagner, J.; Damaschke, N.; Yang, B.; Truong, M.; Guenther, C.; McCormick, J.; Huang, W.; Jarrard, D. Overexpression of the novel senescence marker beta-galactosidase (GLB1) in prostate cancer predicts reduced PSA recurrence. PLoS ONE 2015, 10, e0124366. [Google Scholar] [CrossRef]
- Choi, J.; Shendrik, I.; Peacocke, M.; Peehl, D.; Buttyan, R.; Ikeguchi, E.F.; Katz, A.E.; Benson, M.C. Expression of senescence-associated beta-galactosidase in enlarged prostates from men with benign prostatic hyperplasia. Urology 2000, 56, 160–166. [Google Scholar] [CrossRef]
- Blute, M.L., Jr.; Damaschke, N.; Wagner, J.; Yang, B.; Gleave, M.; Fazli, L.; Shi, F.; Abel, E.J.; Downs, T.M.; Huang, W.; et al. Persistence of senescent prostate cancer cells following prolonged neoadjuvant androgen deprivation therapy. PLoS ONE 2017, 12, e0172048. [Google Scholar] [CrossRef]
- Paradis, V.; Youssef, N.; Dargere, D.; Ba, N.; Bonvoust, F.; Deschatrette, J.; Bedossa, P. Replicative senescence in normal liver, chronic hepatitis C, and hepatocellular carcinomas. Hum. Pathol. 2001, 32, 327–332. [Google Scholar] [CrossRef]
- Ikeda, H.; Sasaki, M.; Sato, Y.; Harada, K.; Zen, Y.; Mitsui, T.; Nakanuma, Y. Bile ductular cell reaction with senescent hepatocytes in chronic viral hepatitis is lost during hepatocarcinogenesis. Pathol. Int. 2009, 59, 471–478. [Google Scholar] [CrossRef]
- Ogawa, S.; Kubo, H.; Murayama, Y.; Kubota, T.; Yubakami, M.; Matsumoto, T.; Yamamoto, Y.; Morimura, R.; Ikoma, H.; Okamoto, K.; et al. Rapid fluorescence imaging of human hepatocellular carcinoma using the beta-galactosidase-activatable fluorescence probe SPiDER-betaGal. Sci. Rep. 2021, 11, 17946. [Google Scholar] [CrossRef]
- Roberson, R.S.; Kussick, S.J.; Vallieres, E.; Chen, S.Y.; Wu, D.Y. Escape from therapy-induced accelerated cellular senescence in p53-null lung cancer cells and in human lung cancers. Cancer Res. 2005, 65, 2795–2803. [Google Scholar] [CrossRef]
- Michaloglou, C.; Vredeveld, L.C.; Soengas, M.S.; Denoyelle, C.; Kuilman, T.; van der Horst, C.M.; Majoor, D.M.; Shay, J.W.; Mooi, W.J.; Peeper, D.S. BRAFE600-associated senescence-like cell cycle arrest of human naevi. Nature 2005, 436, 720–724. [Google Scholar] [CrossRef]
- Cotter, M.A.; Florell, S.R.; Leachman, S.A.; Grossman, D. Absence of senescence-associated beta-galactosidase activity in human melanocytic nevi in vivo. J. Investig. Dermatol. 2007, 127, 2469–2471. [Google Scholar] [CrossRef]
- Gao, S.G.; Zeng, C.; Li, L.J.; Luo, W.; Zhang, F.J.; Tian, J.; Cheng, C.; Tu, M.; Xiong, Y.L.; Jiang, W.; et al. Correlation between senescence-associated beta-galactosidase expression in articular cartilage and disease severity of patients with knee osteoarthritis. Int. J. Rheum. Dis. 2016, 19, 226–232. [Google Scholar] [CrossRef]
- Vasile, E.; Tomita, Y.; Brown, L.F.; Kocher, O.; Dvorak, H.F. Differential expression of thymosin β-10 by early passage and senescent vascular endothelium is modulated by VPF/VEGF: Evidence for senescent endothelial cells in vivo at sites of atherosclerosis. FASEB J. 2001, 15, 458–466. [Google Scholar] [CrossRef]
- Kubo, H.; Murayama, Y.; Ogawa, S.; Matsumoto, T.; Yubakami, M.; Ohashi, T.; Kubota, T.; Okamoto, K.; Kamiya, M.; Urano, Y.; et al. beta-Galactosidase is a target enzyme for detecting peritoneal metastasis of gastric cancer. Sci. Rep. 2021, 11, 10664. [Google Scholar] [CrossRef]
- Miyasaka, Y.; Nagai, E.; Ohuchida, K.; Fujita, H.; Nakata, K.; Hayashi, A.; Mizumoto, K.; Tsuneyoshi, M.; Tanaka, M. Senescence in intraductal papillary mucinous neoplasm of the pancreas. Hum. Pathol. 2011, 42, 2010–2017. [Google Scholar] [CrossRef]
- Kobilkova, J.; Lojda, Z.; Dohnalova, A.; Havrankova, E. Cytologic detection of cervical and endometrial carcinoma with other genital tract involvement. Acta Cytol. 2000, 44, 13–17. [Google Scholar] [CrossRef]
- Laser, J.; Lee, P.; Wei, J.J. Cellular senescence in usual type uterine leiomyoma. Fertil. Steril. 2010, 93, 2020–2026. [Google Scholar] [CrossRef]
- Waszkiewicz, N.; Szajda, S.D.; Waszkiewicz, M.; Wojtulewska-Supron, A.; Szulc, A.; Kepka, A.; Chojnowska, S.; Dadan, J.; Ladny, J.R.; Zwierz, K.; et al. The activity of serum beta-galactosidase in colon cancer patients with a history of alcohol and nicotine dependence: Preliminary data. Postep. Hig. Med. Dosw. 2013, 67, 896–900. [Google Scholar] [CrossRef]
- Nakamura, Y.; Mochida, A.; Nagaya, T.; Okuyama, S.; Ogata, F.; Choyke, P.L.; Kobayashi, H. A topically-sprayable, activatable fluorescent and retaining probe, SPiDER-betaGal for detecting cancer: Advantages of anchoring to cellular proteins after activation. Oncotarget 2017, 8, 39512–39521. [Google Scholar] [CrossRef]
- Li, X.; Pan, Y.; Chen, H.; Duan, Y.; Zhou, S.; Wu, W.; Wang, S.; Liu, B. Specific Near-Infrared Probe for Ultrafast Imaging of Lysosomal beta-Galactosidase in Ovarian Cancer Cells. Anal. Chem. 2020, 92, 5772–5779. [Google Scholar] [CrossRef]
- Zhen, X.; Zhang, J.; Huang, J.; Xie, C.; Miao, Q.; Pu, K. Macrotheranostic Probe with Disease-Activated Near-Infrared Fluorescence, Photoacoustic, and Photothermal Signals for Imaging-Guided Therapy. Angew. Chem. Int. Ed. 2018, 57, 7804–7808. [Google Scholar] [CrossRef]
- Asanuma, D.; Sakabe, M.; Kamiya, M.; Yamamoto, K.; Hiratake, J.; Ogawa, M.; Kosaka, N.; Choyke, P.L.; Nagano, T.; Kobayashi, H.; et al. Sensitive beta-galactosidase-targeting fluorescence probe for visualizing small peritoneal metastatic tumours in vivo. Nat. Commun. 2015, 6, 6463. [Google Scholar] [CrossRef]
- Bunn, P.A., Jr. Chemotherapy for advanced non-small-cell lung cancer: Who, what, when, why? J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2002, 20, 23S–33S. [Google Scholar]
- Van Cutsem, E.; Haller, D.; Ohtsu, A. The role of chemotherapy in the current treatment of gastric cancer. Gastric Cancer 2002, 5 (Suppl. S1), 17–22. [Google Scholar] [CrossRef]
- Martinez-Zamudio, R.I.; Dewald, H.K.; Vasilopoulos, T.; Gittens-Williams, L.; Fitzgerald-Bocarsly, P.; Herbig, U. Senescence-associated beta-galactosidase reveals the abundance of senescent CD8+ T cells in aging humans. Aging Cell 2021, 20, e13344. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Fischer, A.; Reagan, J.D.; Yan, L.J.; Ames, B.N. Oxidative DNA damage and senescence of human diploid fibroblast cells. Proc. Natl. Acad. Sci. USA 1995, 92, 4337–4341. [Google Scholar] [CrossRef] [PubMed]
- Dimri, G.P.; Testori, A.; Acosta, M.; Campisi, J. Replicative senescence, aging and growth-regulatory transcription factors. Biol. Signals 1996, 5, 154–162. [Google Scholar] [CrossRef]
- West, M.D.; Pereira-Smith, O.M.; Smith, J.R. Replicative senescence of human skin fibroblasts correlates with a loss of regulation and overexpression of collagenase activity. Exp. Cell Res. 1989, 184, 138–147. [Google Scholar] [CrossRef]
- Dekker, P.; Maier, A.B.; van Heemst, D.; de Koning-Treurniet, C.; Blom, J.; Dirks, R.W.; Tanke, H.J.; Westendorp, R.G. Stress-induced responses of human skin fibroblasts in vitro reflect human longevity. Aging Cell 2009, 8, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Zhang, C.L.; Liu, X.C.; Qian, G.; Deng, D.Q. Effects of cigarette smoke extracts on the growth and senescence of skin fibroblasts in vitro. Int. J. Biol. Sci. 2013, 9, 613–623. [Google Scholar] [CrossRef]
- Alvarez, D.; Cardenes, N.; Sellares, J.; Bueno, M.; Corey, C.; Hanumanthu, V.S.; Peng, Y.; D’Cunha, H.; Sembrat, J.; Nouraie, M.; et al. IPF lung fibroblasts have a senescent phenotype. Am. J. Physiol. Cell. Mol. Physiol. 2017, 313, L1164–L1173. [Google Scholar] [CrossRef]
- Cui, H.; Ge, J.; Xie, N.; Banerjee, S.; Zhou, Y.; Antony, V.B.; Thannickal, V.J.; Liu, G. miR-34a Inhibits Lung Fibrosis by Inducing Lung Fibroblast Senescence. Am. J. Respir. Cell Mol. Biol. 2017, 56, 168–178. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Condition | Number of Patients | SA-β-Gal Localization | Results | Reference |
|---|---|---|---|---|
| Primary ovarian cancer | 11 | Cells of ovarian cancer not subject to chemotherapy | SA-β-gal activity was detected in 100% of cases of primary ovarian cancer | [24] |
| Epithelial ovarian adenocarcinoma | 40 | Healthy tissues and epithelial ovarian adenocarcinoma | SA-β-gal activity in tumor tissues is 50% higher than in healthy tissues | [25] |
| Colon adenocarcinoma | 7 | Colon adenocarcinoma tissue | SA-β-gal activity in adenocarcinoma tissues is 2 times higher than normal | [26] |
| Colon adenoma and untreated invasive carcinoma | 23 | Senescent cells within neoplastic epithelial areas of manifest colorectal carcinomas | SA-β-gal activity was found in 8 out of 12 adenomas, 1 of out 6 invasive carcinomas, 0 out of 5 normal crypt mucosa tissues | [27] |
| Colon adenoma | 59 | Cells of colon adenomas | A strong correlation between SA-β-gal staining and senescence immunohistochemical markers | [28] |
| Glioma | 23 | Cells of malignant glial tumors | The greatest increase in the activity of β-galactosidase was in anaplastic oligodendroglioma; in other glial tumors, it was also higher than in meningiomas and metastatic tumors | [21] |
| Glioma | 5 | Tumor-associated brain endothelial cells | SA-β-gal activity was detected in glioma tissue cells, while normal brain tissue was negative | [29] |
| Breast cancer, fibroadenoma, fibrocystic disease | 18 | Carcinomas, fibrocystic, fibroadenomatous tissues | SA-β-gal activity was normal in fibroadenoma cells, increased in fibrocystic cells, and maximum level was observed in breast cancer cells | [26] |
| Breast cancer | 56 | Tissues of treated and untreated breast tumors | SA-β-gal activity was detected in 15 of 36 tumors. Tumor sections of patients who had not received chemotherapy expressed SA-β-gal in 2 of 20 cases | [30] |
| Prostate cancer, benign prostatic hyperplasia, high-grade prostatic intraepithelial neoplasia | 124 | Human prostate epithelial cells | High expression of SA-β-gal was observed in 37% of primary cancer specimens, 72% of high-grade prostatic intraepithelial neoplasia samples, and 13% of benign tissues | [31] |
| Benign prostatic hyperplasia | 43 | Human prostate epithelial cells of hypertrophied tissues | SA-β-gal activity was detected in 17 out of 43 specimens. Prostate epithelial cells expressed SA-β-gal in patients with more pronounced prostate enlargement weighing more than 55 g, while the senescent cells were absent in prostate cells weighing less than 55 g | [32] |
| Prostate cancer | 126 | Senescent prostate cancer cells | GLB1 staining was expressed at higher levels in prostate samples treated with androgen deprivation | [33] |
| Chronic hepatitis C, hepatocellular carcinoma | 57 | Replicating cells in normal liver, liver with chronic hepatitis C and hepatocellular carcinomas | In normal liver tissue, SA-β-gal activity can be detected in 20% of cases. SA-β-gal was expressed in 60% of hepatocellular carcinoma samples and in 50% of samples with hepatitis C | [34] |
| Chronic viral hepatitis | 20 | Tissues of liver with chronic viral hepatitis B, chronic viral hepatitis C, and normal liver | The SA-β-gal activity was frequently detected in periportal or periseptal hepatocytes of liver cirrhosis and focally in chronic hepatitis irrespective of type B or type C infection, while the enzyme activity was extremely weak in normal livers | [35] |
| Hepatocellular carcinoma | 95 | Cancer cells | High SA-β-gal activity in tumor and low SA-β-gal activity in non-tumor tissues | [36] |
| Lung Cancer | 6 | Tumor cells treated with chemotherapy and radiation | The enzyme was not detected in samples from patients who did not receive chemotherapy. SA-β-gal expression elevated in tissues after neoadjuvant chemotherapy | [37] |
| Melanocytic naevi | 23 | Tissues of congenital naevi | Human naevi, largely growth-arrested neoplastic lesions, are positive for the senescence marker SA-β-gal | [38] |
| Melanocytic naevi | 17 | Adult nevi cells | Every specimen evaluated showed varying degrees of positivity at the optimal pH 4, none of the specimens showed staining at pH 6 | [39] |
| Knee osteoarthritis | 50 | Chondrocytes in articular cartilage | SA-β-gal staining was found in a subset of chondrocytes close to the lesion site of mild, moderate, and severely altered knee cartilage with osteoarthritis. No SA-β-gal staining was observed in normal articular cartilage samples | [40] |
| Atherosclerosis | 3 | Atherosclerotic aorta | The aortic endothelium cells overlying atherosclerotic plaques were SA-β-gal-positive. The endothelium covering nearby regions of relatively normal aorta was SA-β-gal-negative | [41] |
| Gastric cancer | 13 | Gastric cancer cells, peritoneum metastatic cells | The SA-β-gal content in the tumor was high. SA-β-gal allowed locating peritoneal metastases | [42] |
| Intraductal papillary mucinous neoplasm of the pancreas | 39 | Neoplasms with low-, intermediate- and high-grade dysplasia, associated invasive carcinoma | Senescence is induced in the early stage of intraductal papillary mucinous neoplasm and gradually attenuated according to the progression | [43] |
| Cervical and endometrial carcinoma | 77 | Tissues of invasive cervical cancer and endometrial cancer | Squamous cell carcinoma (negative reaction) and cylindrocellular carcinoma (positive reaction in cancer cells) | [44] |
| Usual type uterine leiomyoma | 14 (86 samples) | Fibroid tissue sections | 48 out of 82 tumors were SA-β-gal positive in >10% of the tumor volume. The more senescent cells, the higher the stage (1–4) and the lower the growth potential of the tumor | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valieva, Y.; Ivanova, E.; Fayzullin, A.; Kurkov, A.; Igrunkova, A. Senescence-Associated β-Galactosidase Detection in Pathology. Diagnostics 2022, 12, 2309. https://doi.org/10.3390/diagnostics12102309
Valieva Y, Ivanova E, Fayzullin A, Kurkov A, Igrunkova A. Senescence-Associated β-Galactosidase Detection in Pathology. Diagnostics. 2022; 12(10):2309. https://doi.org/10.3390/diagnostics12102309
Chicago/Turabian StyleValieva, Yana, Elena Ivanova, Alexey Fayzullin, Alexander Kurkov, and Alexandra Igrunkova. 2022. "Senescence-Associated β-Galactosidase Detection in Pathology" Diagnostics 12, no. 10: 2309. https://doi.org/10.3390/diagnostics12102309