
Differential Vaginal Microbiota Profiling in Lactic-Acid-Producing Bacteria between Infertile Women with and without Chronic Endometritis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects, Sample Collection, and Examinations
2.2. Immunohistochemistry for CD138 and Diagnosis of CE
2.3. DNA Extraction and Sequencing
2.4. Statistics
3. Results
3.1. Characteristics of Patients
3.2. Sequencing Results of VS and EF Samples
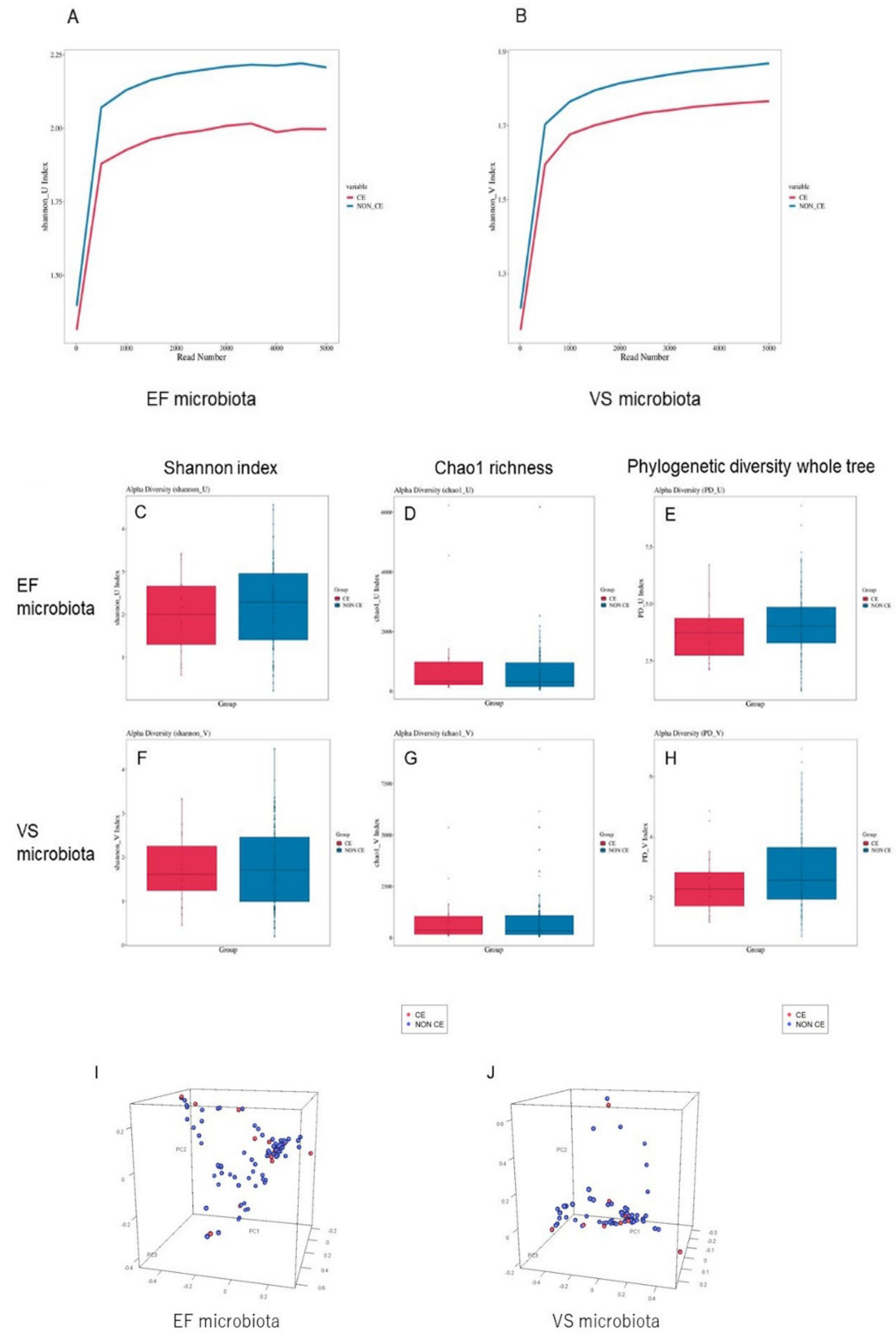
3.3. Diversity Comparison of VS and EF Microbiota between Infertile Women with and without CE
3.4. Comparison of Bacterial Genera in EF Microbiota between Infertile Women with and without CE
3.5. Comparison of Bacterial Genera in VS Microbiota between Infertile Patients with and without CE
3.6. Comparison of Lactic-Acid-Producing Bacteria in Microbiota between Infertile Patients with and without CE
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kitaya, K.; Takeuchi, T.; Mizuta, S.; Matsubayashi, H.; Ishikawa, T. Endometritis: New time, new concepts. Fertil. Steril. 2018, 110, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; De Ziegler, D.; Nicoletti, R.; Colafiglio, G.; Saliani, N.; Resta, L.; Rizzi, D.; De Vito, D. Chronic endometritis: Correlation among hysteroscopic, histologic, and bacteriologic findings in a prospective trial with 2190 consecutive office hysteroscopies. Fertil. Steril. 2008, 89, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Bouet, P.-E.; El Hachem, H.; Monceau, E.; Gariépy, G.; Kadoch, I.-J.; Sylvestre, C. Chronic endometritis in women with recurrent pregnancy loss and recurrent implantation failure: Prevalence and role of office hysteroscopy and immunohistochemistry in diagnosis. Fertil. Steril. 2016, 105, 106–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston-MacAnanny, E.B.; Hartnett, J.; Engmann, L.L.; Nulsen, J.C.; Sanders, M.M.; Benadiva, C.A. Chronic endometritis is a fre-quent finding in women with recurrent implantation failure after in vitro fertilization. Fertil. Steril. 2010, 93, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Kitaya, K.; Tada, Y.; Taguchi, S.; Funabiki, M.; Hayashi, T.; Nakamura, Y. Local mononuclear cell infiltrates in infertile patients with endometrial macropolyps versus micropolyps. Hum. Reprod. 2012, 27, 3474–3480. [Google Scholar] [CrossRef] [PubMed]
- McQueen, D.; Bernardi, L.A.; Stephenson, M.D. Chronic endometritis in women with recurrent early pregnancy loss and/or fetal demise. Fertil. Steril. 2014, 101, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; Matteo, M.; Tinelli, R.; Lepera, A.; Alfonso, R.; Indraccolo, U.; Marrocchella, S.; Greco, P.; Resta, L. Prevalence of chronic endometritis in repeated un-explained implantation failure and the IVF success rate after antibiotic therapy. Hum. Reprod. 2015, 30, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Kitaya, K.; Matsubayashi, H.; Takaya, Y.; Nishiyama, R.; Yamaguchi, K.; Takeuchi, T.; Ishikawa, T. Live birth rate following oral antibiotic treatment for chronic endometritis in infertile women with repeated implantation failure. Am. J. Reprod. Immunol. 2017, 78, e12719. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; Matteo, M.; Trojano, G.; Mitola, P.C.; Tinelli, R.; Vitagliano, A.; Crupano, F.M.; Lepera, A.; Miragliotta, G.; Resta, L. Chronic endometritis in patients with unexplained infertility: Prevalence and effects of antibiotic treatment on spontaneous conception. Am. J. Reprod. Immunol. 2018, 79, e12782. [Google Scholar] [CrossRef] [PubMed]
- Vitagliano, A.; Saccardi, C.; Noventa, M.; Sardo, A.D.S.; Saccone, G.; Cicinelli, E.; Pizzi, S.; Andrisani, A.; Litta, P.S. Effects of chronic endometritis therapy on in vitro fertilization outcome in women with repeated implantation failure: A systematic review and meta-analysis. Fertil. Steril. 2018, 110, 103–112.e1. [Google Scholar] [CrossRef] [PubMed]
- Wiesenfeld, H.C.; Hillier, S.L.; Krohn, M.A.; Amortegui, A.J.; Heine, R.P.; Landers, D.V.; Sweet, R.D. Lower genital tract infection and endome-tritis: Insight into subclinical pelvic inflammatory disease. Obstet. Gynecol. 2002, 100, 456–463. [Google Scholar]
- Mishra, K.; Wadhwa, N.; Guleria, K.; Agarwal, S. ER, PR and Ki-67 expression status in granulomatous and chronic non-specific endometritis. J. Obstet. Gynaecol. Res. 2008, 34, 371–378. [Google Scholar] [CrossRef]
- Di Pietro, C.; Cicinelli, E.; Guglielmino, M.R.; Ragusa, M.; Farina, M.; Palumbo, M.A.; Cianci, A. Altered transcriptional regulation of cyto-kines, growth factors, and apoptotic proteins in the endometrium of infertile women with chronic endometritis. Am. J. Reprod. Immunol. 2013, 69, 509–517. [Google Scholar] [CrossRef]
- Wu, D.; Kimura, F.; Zheng, L.; Ishida, M.; Niwa, Y.; Hirata, K.; Takebayashi, A.; Takashima, A.; Takahashi, K.; Kushima, R. Chronic endometritis modifies decidualization in human endo-metrial stromal cells. Reprod. Biol. Endocrinol. 2017, 15, 16. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Kim, J.-S. Chronic inflammation of the placenta: Definition, classification, pathogenesis, and clinical significance. Am. J. Obstet. Gynecol. 2015, 213, S53–S69. [Google Scholar] [CrossRef] [Green Version]
- Ghidini, A.; Salafia, C.M. Histologic placental lesions in women with recurrent preterm delivery. Acta Obstet. Gynecol. Scand. 2005, 84, 547–550. [Google Scholar] [CrossRef]
- Perni, S.C.; Predanik, M.; Cho, J.E.; Baergen, R.N. Placental pathology and pregnancy outcomes in donor and non-donor oocyte in vitro fertilization pregnancies. J. Périnat. Med. 2005, 33, 27–32. [Google Scholar] [CrossRef]
- Gundogan, F.; Bianchi, D.W.; Scherjon, S.A.; Roberts, D.J. Placental pathology in egg donor pregnancies. Fertil. Steril. 2010, 93, 397–404. [Google Scholar] [CrossRef]
- Romero, R.; Whitten, A.; Korzeniewski, S.J.; Than, N.G.; Chaemsaithong, P.; Miranda, J.; Dong, Z.; Hassan, S.S.; Chaiworapongsa, T. Maternal floor infarction/massive pe-rivillous fibrin deposition: A manifestation of maternal antifetal rejection? Am. J. Reprod. Immunol. 2013, 70, 285–298. [Google Scholar] [CrossRef] [Green Version]
- Raman, K.; Wang, H.; Troncone, M.J.; Khan, W.I.; Pare, G.; Terry, J. Overlap Chronic Placental Inflammation Is Associated with a Unique Gene Expression Pattern. PLoS ONE 2015, 10, e0133738. [Google Scholar] [CrossRef] [Green Version]
- Edmondson, N.; Bocking, A.; Machin, G.; Rizek, R.; Watson, C.; Keating, S. The Prevalence of Chronic Deciduitis in Cases of Preterm Labor without Clinical Chorioamnionitis. Pediatr. Dev. Pathol. 2009, 12, 16–21. [Google Scholar] [CrossRef]
- Maleki, Z.; Bailis, A.J.; Argani, C.H.; Askin, F.B.; Graham, E.M. Periventricular leukomalacia and placental histopathologic abnormalities. Obs. Gynecol. 2009, 114, 1115–1120. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4680–4687. [Google Scholar] [CrossRef] [Green Version]
- Plummer, E.L.; Bradshaw, C.S.; Doyle, M.; Fairley, C.K.; Murray, G.L.; Bateson, D.; Masson, L.; Slifirski, J.; Tachedjian, G.; Vodstrcil, L.A. Lactic acid-containing products for bacterial vaginosis and their impact on the vaginal microbiota: A systematic review. PLoS ONE 2021, 16, e0246953. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Haick, A.; Nkwopara, E.; Garcia, R.; Rendi, M.; Agnew, K.; Fredricks, D.; Eschenbach, D. Colonization of the upper genital tract by vaginal bacterial species in nonpregnant women. Am. J. Obstet. Gynecol. 2015, 212, 611.e1–611.e9. [Google Scholar] [CrossRef] [Green Version]
- Franasiak, J.M.; Werner, M.D.; Juneau, C.R.; Tao, X.; Landis, J.; Zhan, Y.; Treff, N.R.; Scott, R.T. Endometrial microbiome at the time of embryo transfer: Next-generation sequencing of the 16S ribosomal subunit. J. Assist. Reprod. Genet. 2016, 33, 129–136. [Google Scholar] [CrossRef]
- Chen, C.; Song, X.; Chunwei, Z.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef] [Green Version]
- Verstraelen, H.; Vilchez-Vargas, R.; Desimpel, F.; Jauregui, R.; Vankeirsbilck, N.; Weyers, S.; Verhelst, R.; De Sutter, P.; Pieper, D.H.; Van De Wiele, T. Characterisation of the human uterine microbiome in non-pregnant women through deep sequencing of the V1-2 region of the 16S rRNA gene. PeerJ 2016, 4, e1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, R.-L.; Chen, L.-X.; Shu, W.-S.; Yao, S.-Z.; Wang, S.-W.; Chen, Y.-Q. Barcoded sequencing reveals diverse intrauterine microbiomes in patients suffering with endometrial polyps. Am. J. Transl. Res. 2016, 8, 1581–1592. [Google Scholar] [PubMed]
- Kyono, K.; Hashimoto, T.; Kikuchi, S.; Nagai, Y.; Sakuraba, Y. A pilot study and case reports on endometrial microbiota and preg-nancy outcome: An analysis using 16S rRNA gene sequencing among IVF patients, and trial therapeutic intervention for dysbiotic endometrium. Reprod. Med. Biol. 2019, 18, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyono, K.; Hashimoto, T.; Nagai, Y.; Sakuraba, Y. Analysis of endometrial microbiota by 16S ribosomal RNA gene sequencing among infertile patients: A single-center pilot study. Reprod. Med. Biol. 2018, 17, 297–306. [Google Scholar] [CrossRef]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef] [Green Version]
- Miles, S.M.; Hardy, B.L.; Merrell, D.S. Investigation of the microbiota of the reproductive tract in women undergoing a total hys-terectomy and bilateral salpingo-oopherectomy. Fertil. Steril. 2017, 107, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Walther-António, M.R.; Chen, J.; Multinu, F.; Hokenstad, A.; Distad, T.J.; Cheek, E.H.; Keeney, G.L.; Creedon, J.L.; Nelson, H.; Mariani, A.; et al. Potential contribution of the uterine mi-crobiome in the development of endometrial cancer. Genome Med. 2016, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Tao, X.; Franasiak, J.M.; Zhan, Y.; Scott, R.T.; Rajchel, J.; Bedard, J.; Newby, R., Jr.; Scott, R.T.; Treff, N.R.; Chu, T. Characterizing the endometrial microbiome by analyzing the ultra-low bacteria from embryo transfer catheter tips in IVF cycles: Next generation sequencing (NGS) analysis of the 16S ri-bosomal gene. Hum. Microbiome J. 2017, 3, 15–21. [Google Scholar] [CrossRef]
- Kitaya, K.; Nagai, Y.; Arai, W.; Sakuraba, Y.; Ishikawa, T. Characterization of microbiota in endometrial fluid and vaginal secre-tions in infertile women with repeated implantation failure. Mediat. Inflamm. 2019, 2019, 4893437. [Google Scholar] [CrossRef] [Green Version]
- Winters, A.D.; Romero, R.; Gervasi, M.T.; Gomez-Lopez, N.; Tran, M.R.; Garcia-Flores, V.; Pacora, P.; Jung, E.; Hassan, S.S.; Hsu, C.-D.; et al. Does the endometrial cavity have a molecular microbial signature? Sci. Rep. 2019, 9, 9905. [Google Scholar] [CrossRef]
- Leoni, C.; Ceci, O.; Manzari, C.; Fosso, B.; Volpicella, M.; Ferrari, A.; Fiorella, P.; Pesole, G.; Cincinelli, E.; Ceci, L. Human endometrial microbiota at term of normal preg-nancies. Genes 2019, 10, 971. [Google Scholar] [CrossRef] [Green Version]
- Cicinelli, E.; De Ziegler, D.; Nicoletti, R.; Tinelli, R.; Saliani, N.; Resta, L.; Bellavia, M.; De Vito, D. Poor Reliability of Vaginal and Endocervical Cultures for Evaluating Microbiology of Endometrial Cavity in Women with Chronic Endometritis. Gynecol. Obstet. Investig. 2009, 68, 108–115. [Google Scholar] [CrossRef]
- Cicinelli, E.; Vitagliano, A.; Kumar, A.; Lasmar, R.B.; Bettocchi, S.; Haimovich, S. International Working Group for Standardization of Chronic Endometritis Diagnosis. Unified diagnostic criteria for chronic endometritis at fluid hysteroscopy: Proposal and re-liability evaluation through an international randomized-controlled observer study. Fertil. Steril. 2019, 112, 162–173. [Google Scholar]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.; et al. Improved Bacterial 16S rRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems 2016, 1, e00009-15. [Google Scholar] [CrossRef] [Green Version]
- Aronesty, E. Comparison of Sequencing Utility Programs. Open Bioinform. J. 2013, 7, 1–8. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bac-terial taxonomy. Appl. Env. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurence, M.; Hatzis, C.; Brash, D.E. Common Contaminants in Next-Generation Sequencing That Hinder Discovery of Low-Abundance Microbes. PLoS ONE 2014, 9, e97876. [Google Scholar] [CrossRef] [PubMed]
- Urushiyama, D.; Suda, W.; Ohnishi, E.; Araki, R.; Kiyoshima, C.; Kurakazu, M.; Sanui, A.; Yotsumoto, F.; Murata, M.; Nabeshima, K.; et al. Microbiome profile of the amniotic fluid as a predictive biomarker of perinatal outcome. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ko, E.Y.-L.; Wong, K.K.-W.; Chen, X.; Cheung, W.-C.; Law, T.S.-M.; Chung, J.; Tsui, S.K.-W.; Li, T.C.; Chim, S.S.-C. Endometrial microbiota in infertile women with and without chronic endometritis as diagnosed using a quantitative and reference range-based method. Fertil. Steril. 2019, 112, 707–717.e1. [Google Scholar] [CrossRef]
- Punnonen, R.; Lehtinen, M.; Teisala, K.; Aine, R.; Rantala, I.; Heinonen, P.K.; Miettinen, A.; Laine, S.; Paavonen, J. The relation between serum sex steroid levels and plasma cell infiltrates in endometritis. Arch. Gynecol. Obstet. 1989, 244, 185–191. [Google Scholar] [CrossRef]
- Eckert, L.O.; Hawes, S.E.; Wölner-Hanssen, P.K.; Kiviat, N.B.; Wasserheit, J.N.; Paavonen, J.A.; Eschenbach, D.A.; Holmes, K.K. Endometritis: The clinical-pathologic syndrome. Am. J. Obstet. Gynecol. 2002, 186, 690–695. [Google Scholar] [CrossRef]
- Song, D.; Feng, X.; Zhang, Q.; Xia, E.; Xiao, Y.; Xie, W.; Li, T.C. Prevalence and confounders of chronic endometritis in premenopausal women with abnormal bleeding or reproductive failure. Reprod. Biomed. Online 2018, 36, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Green, S.J.; Prakash, O.; Jasrotia, P.; Overholt, W.A.; Cardenas, E.; Hubbard, D.; Tiedje, J.M.; Watson, D.B.; Schadt, C.W.; Brooks, S.C.; et al. Denitrifying bacteria from the genus Rhodano-bacter dominate bacterial communities in the highly contaminated subsurface of a nuclear legacy waste site. Appl. Env. Microbiol. 2012, 78, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Grau, I.; Perez-Villaroya, D.; Bau, D.; Gonzalez-Monfort, M.; Vilella, F.; Moreno, I.; Simon, C. Taxonomical and Functional As-sessment of the Endometrial Microbiota in A Context of Recurrent Reproductive Failure: A Case Report. Pathogens 2019, 8, 205. [Google Scholar] [CrossRef] [Green Version]
- Onderdonk, A.B.; Delaney, M.L.; Fichorova, R.N. The Human Microbiome during Bacterial Vaginosis. Clin. Microbiol. Rev. 2016, 29, 223–238. [Google Scholar] [CrossRef] [Green Version]
- Haggerty, C.L.; Hillier, S.L.; Bass, D.C.; Ness, R.B.; the PID Evaluation and Clinical Health (PEACH) Study Investigators. Bacterial Vaginosis and Anaerobic Bacteria Are Associated with Endometritis. Clin. Infect. Dis. 2004, 39, 990–995. [Google Scholar] [CrossRef] [Green Version]
- Drizi, A.; Djokovic, D.; Laganà, A.S.; van Herendael, B. Impaired inflammatory state of the endometrium: A multifaceted ap-proach to endometrial inflammation. Current insights and future directions. Prz. Menopauzalny 2020, 19, 90–100. [Google Scholar] [CrossRef]
- Puente, E.; Alonso, L.; Laganà, A.S.; Ghezzi, F.; Casarin, J.; Carugno, J. Chronic Endometritis: Old Problem, Novel Insights and Future Challenges. Int. J. Fertil. Steril. 2019, 13, 250–256. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Strawberry aspect | 10% (2/20) |
| Focal hyperemia | 20% (4/20) |
| Hemorrhagic spots | 15% (3/20) |
| Micropolyposis | 25% (5/20) |
| Stromal edema | 30% (6/20) |
| Total | 40% (8/20) a |
| CE Group (n = 20) | Non-CE Group (n = 103) | p Value c | ||
|---|---|---|---|---|
| Age (years), mean ± SD | 38.1 ± 3.7 | 38.4 ± 4.1 | 0.76 | |
| Body mass index (kg/m2), mean ± SD | 22.1 ± 1.8 | 21.8 ± 2.3 | 0.58 | |
| Gravidity, median (range) | 0 (0–3) | 0 (0–4) | 0.65 | |
| Parity, median (range) | 0 (0–1) | 0 (0–1) | 0.84 | |
| Indication of IVF-ET a | ||||
| Male factor | 4 (20.0%) | 29 (28.2%) | 0.59 | |
| Polycystic ovarian syndrome | 1 (5.0%) | 16 (15.5%) | 0.30 | |
| Endometriosis | 3 (15.0%) | 25 (24.3%) | 0.56 | |
| Tubal factor | 4 (20.0%) | 10 (9.7%) | 0.24 | |
| Unexplained | 8 (40.0%) | 19 (18.4%) | 0.042 | |
| Diminished Ovarian Reserve | 3 (15.0%) | 35 (34.0%) | 0.12 | |
| Controlled ovarian stimulation protocol history b | ||||
| Short gonadotropin-releasing hormone agonist cycle | 12 (60.0%) | 37 (35.9%) | 0.0503 | |
| Long gonadotropin-releasing hormone agonist cycle | 0 (0.0%) | 1 (0.1%) | 1.00 | |
| Flexible GnRH antagonist cycle | 15 (80.0%) | 82 (79.6%) | 0.76 | |
| Mild stimulation cycle | 4 (20.0%) | 33 (32.0%) | 0.42 | |
| Natural cycle | 0 (0.0%) | 4 (3.9%) | 1.00 | |
| Past embryo transfer history, mean ± SD | ||||
| Number of cycles | 4.7 ± 3.6 | 4.4 ± 2.9 | 0.69 | |
| Number of embryos transferred | 5.9 ± 4.5 | 5.3 ± 3.8 | 0.53 | |
| CE Group (n = 20) | Non-CE Group (n = 103) | Fisher’s Exact Test p Value | Wilcoxon Rank-Sum Test p Value | Adjusted Welch’s t Test p Value | |
|---|---|---|---|---|---|
| EF microbiota | |||||
| Rhodanobacter | 3 (15.0%) | 2 (1.94%) | 0.030 | 0.0062 | 0.12 |
| Atopobium | 2 (10.0%) | 24 (23.3%) | 0.24 | 0.13 | 0.0022 |
| Bifidobacterium | 2 (10.0%) | 19 (18.5%) | 0.52 | 0.30 | 0.0054 |
| Aeromonadaceae | 0 (0.0%) | 11 (10.7%) | 0.21 | 0.13 | 0.014 |
| Vibrio | 2 (10.0%) | 19 (18.5%) | 0.52 | 0.33 | 0.037 |
| Clostridiales | 0 (0.0%) | 11 (10.7%) | 0.69 | 0.41 | 0.048 |
| VS microbiota | |||||
| Streptococcus | 1 (5.0%) | 40 (38.8%) | 0.0033 | 0.0072 | 0.054 |
| Enterococcus | 0 (0.0%) | 18 (17.5%) | 0.042 | 0.045 | 0.67 |
| Atopobium | 0 (0.0%) | 16 (15.5%) | 0.072 | 0.061 | 0.0029 |
| Bifidobacterium | 1 (5.0%) | 18 (17.5%) | 0.31 | 0.15 | 0.0044 |
| Nesterenkonia | 0 (0.0%) | 6 (5.8%) | 0.59 | 0.27 | 0.019 |
| Anaerococcus | 0 (0.0%) | 9 (8.7%) | 0.35 | 0.17 | 0.024 |
| Staphylococcus | 0 (0.0%) | 5 (4.9%) | 0.59 | 0.32 | 0.033 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, S.E.; Sakuraba, Y.; Kitaya, K.; Ishikawa, T. Differential Vaginal Microbiota Profiling in Lactic-Acid-Producing Bacteria between Infertile Women with and without Chronic Endometritis. Diagnostics 2022, 12, 878. https://doi.org/10.3390/diagnostics12040878
Tanaka SE, Sakuraba Y, Kitaya K, Ishikawa T. Differential Vaginal Microbiota Profiling in Lactic-Acid-Producing Bacteria between Infertile Women with and without Chronic Endometritis. Diagnostics. 2022; 12(4):878. https://doi.org/10.3390/diagnostics12040878
Chicago/Turabian StyleTanaka, Suguru E., Yoshiyuki Sakuraba, Kotaro Kitaya, and Tomomoto Ishikawa. 2022. "Differential Vaginal Microbiota Profiling in Lactic-Acid-Producing Bacteria between Infertile Women with and without Chronic Endometritis" Diagnostics 12, no. 4: 878. https://doi.org/10.3390/diagnostics12040878
APA StyleTanaka, S. E., Sakuraba, Y., Kitaya, K., & Ishikawa, T. (2022). Differential Vaginal Microbiota Profiling in Lactic-Acid-Producing Bacteria between Infertile Women with and without Chronic Endometritis. Diagnostics, 12(4), 878. https://doi.org/10.3390/diagnostics12040878