Abstract
S100 proteins are a family of low-molecular-weight proteins characterized by two calcium-binding sites with a helix-loop-helix (“EF-hand-type”) domain. The S100 family of proteins is distributed across various organs and can interact with diverse molecules. Among the proteins of the S100 family, S100 calcium-binding protein A2 (S100A2) has been identified in mammary epithelial cells, glands, lungs, kidneys, and prostate gland, exhibiting various physiological and pathological actions in human disorders, such as inflammatory diseases and malignant tumors. In this review, we introduce basic knowledge regarding S100A2 regulatory mechanisms. Although S100A2 is a tumor suppressor, we describe the various influences of S100A2 on cancer and inflammatory diseases.
1. Introduction
S100 proteins are a family of low-molecular-weight proteins characterized by having two calcium-binding sites with a helix-loop-helix (“EF-hand-type”) domain [1]. Because S100 family proteins are distributed across various organs and can interact with a diverse range of molecules [2,3,4,5], they play vital roles in numerous aspects of physiological and pathological events. They are currently highlighted for investigators to elucidate their role in human diseases.
Among the S100 family proteins, S100 calcium-binding protein A2 (S100A2) is encoded by a gene located on chromosome 1q21 [6]. Although S100A2 interacts with calcium, S100A2 also binds to zinc with a high affinity [7]. Zinc inactivates S100A2 function by inhibiting response to intracellular calcium signals [7]. S100A2 has been identified in mammary epithelial cells, glands, lungs, kidneys, and prostate gland [8]. Therefore, S100A2 is expected to have various physiological and pathological effects on human diseases.
Although S100A2 has a tumor suppressor function [9], the recently updated research focusing on the prognostic impact of S100A2 revealed the various influences of S100A2 in human malignancies [10,11], in addition to the inflammatory diseases [12]. In this review, we introduce basic knowledge surrounding S100A2 and its regulatory mechanisms. Furthermore, we describe in detail the influence of S100A2 on malignant tumors and inflammatory diseases.
2. Regulatory Action of S100A2 and Interaction with Other Molecules
Human S100A2 is an EF-hand calcium-binding S100 protein predominantly located in epithelial cells [13]. In particular, S100A2 is expressed in keratinocytes in a dimeric form in normal physical state [14]. S100A2 is predominantly expressed in the nucleus and moderately so in the cytoplasm [15]. However, this distribution pattern of S100A2 is altered under stress stimuli, such as oxidative stress, which enhances the cytoplasmic translocation of S100A2 from the nucleus [15]. This phenomenon can be observed under conditions of increased intracellular Ca2+ levels [15]. This cytoplasmic S100A2 is released by the leakage from the S100A2 containing cells due to cell death events.
Various other stimuli, including transforming growth factor (TGF)-β [16] and interferon (IFN)-α [17], can also trigger modulation of S100A2 expression (Figure 1). Therefore, various inflammatory reactions appear to be associated with S100A2-mediated physiological and pathological reactions in the human body.
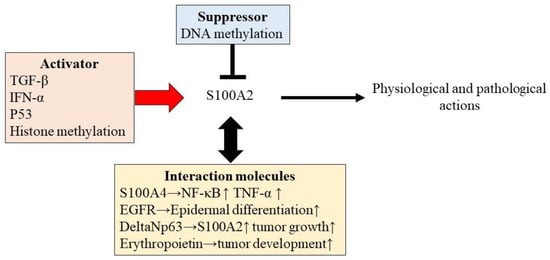
Figure 1.
The regulation of S100A2 expression.
3. Interactions of S100A with Other Molecules
A recent study revealed that S100A2 interacts with various physiological regulatory factors to enhance its regulatory potential. S100A2 interacts with other S100 family proteins, such as S100A4 [18]. S100A2 interacts with S100A4 in a cross-linked manner, mediated by the copper-mediated oxidation of cysteine residues, which increases the induction of NF-κB and TNF-α production [18] (Figure 1).
S100A2 is localized to an intermediate filament cytoplasmic protein, cytokeratin K14, which is predominantly present in basal proliferative keratinocytes [19]. S100A2 is also an enhancer of epidermal differentiation in the regenerative hyperplasia pathway, which is mediated by the epidermal growth factor (EGF) receptor and/or related receptors of ErbB family-mediated pathway activation [20]. These findings suggest an important role for S100A2 in the epidermal physiology and oncogenesis of tumors derived from the epidermis.
S100A2 also plays an important role in the pathogenesis of tumors. Indeed, S100A2 is also induced by the p53-stimulator etoposide [21]. A possible binding site for p53 is located at the promoter site of S100A2 [21]. Therefore, p53 can positively regulate S100A2 expression [21,22,23,24]. S100A2 is also a direct transcriptional target of ΔNp63alpha and is essential for keratinocyte differentiation [25]. Further, BRCA1 can interact with ΔNp63 proteins, enhancing S100A2 expression and tumor growth [26].
In contrast, erythropoietin can interact with S100A2 to enhance tumor development [27]. Since S100A2 plays a role in regulating tumor development in almost all tumors, the interaction of S100A2 with erythropoietin might be important in certain conditions, such as erythropoietin-dominant tumor development.
4. Epigenetic Regulation of S100A2
Epigenetic regulation has been postulated as one mechanism underlying S100A2 regulation, especially in malignancies. Epigenetic chemical modification of DNA itself or DNA-binding proteins, such as histones, changes gene expression without altering the sequence [28,29,30] (Figure 1).
Among epigenetic chemical modifications, deoxyribonucleic acid (DNA) methylation is a representative epigenetic modification [31,32,33]. CpG islands are often modified by DNA methylation, and typically influence CpG islands on the promoter sites of target genes [34]. Such sites comprise a cytosine nucleotide followed by a guanine nucleotide in a linear sequence from the 5′ to the 3′ direction of the DNA region. DNA methylation essentially acts to silence targeted gene expression [35]. As CpG islands are often observed in gene promoter sites, DNA methylation is a powerful transcriptional modulation mechanism in various diseases [36]. Indeed, the hypermethylation of targeted gene promoter CpG sites is often observed in cancer cells [9,37]. In addition, targeted gene-specific DNA methylation leads to the reduced expression of S100A2 in normal cells [9], while treatment with demethylation agent 5-aza-2′-deoxycytidine can recover the expression of S100A2 [38]. These findings indicate that DNA methylation is a mechanism for regulating S100A2 gene expression.
Histone methylation primarily targets histone H3 lysine residues, which can drive both the activation and suppression of gene transcription [28]. Histone methyltransferases enhance histone methylation, whereas histone demethylases reverse histone methylation. Lysine-specific demethylase 5C (KDM5C) is a lysine-specific histone demethylase that belongs to the Jumonji family of demethylases, and is specific for di- and tri-demethylation of lysine 4 residues on histone 3 (H3K4 me2/3) [39]. KDM5C recovers S100A2 expression through histone demethylation to impair the proliferation and migration of cancer cells, suggesting that histone methylation leads to the impairment of S100A2 expression in tumors [40].
5. Inflammatory Diseases
Although a limited number of studies have investigated inflammatory diseases, previous studies have speculated about the possible role of S100A2 in the development of inflammatory diseases (Figure 2).
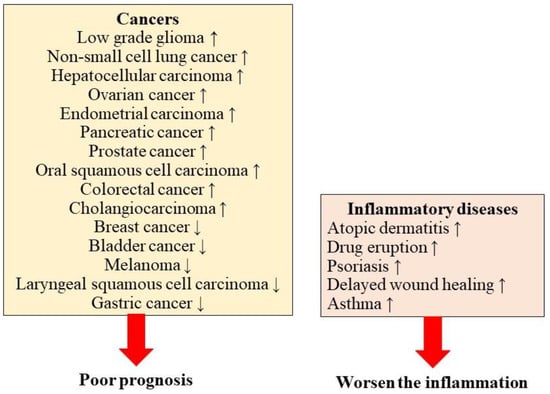
Figure 2.
The influence of S100A2 in human diseases.
S100A2 is highly upregulated in the epidermis under inflammatory conditions and in drug eruptions, in addition to inflammatory skin diseases such as atopic dermatitis and psoriasis [12]. An important finding is that the degree of S100A2 expression depends on the severity of inflammatory skin diseases [12]. Furthermore, epithelial-specific S100A2 transgenic mice showed the activation of the proliferation and migration of S100A2-deficient human keratinocytes [41]. In addition, S100A2-expressing mice exhibit a delayed response to cutaneous wound repair action [41]. The stress-activated p53 tumor suppressor protein plays an important role in cutaneous wound healing and is an S100A2 inducer [41]. Although it remains unclear whether S100A2 is involved in physiological function in hair follicles, S100A2 is highly expressed in the entire outer-root sheath, whereas its expression is lower in the medulla and cuticle in the bulb [42], suggesting that alopecia areata or other hair-related diseases might also be associated with S100A2-mediated pathogenesis.
The exposure of lung epithelial cells to Ni enhances S100A2 expression, suggesting that S100A2 is involved in lung inflammatory diseases [43]. Consistently, S100A2 levels are increased in the saliva of patients with uncontrolled asthma [44], suggesting that lung inflammatory diseases are also associated with S100A2-mediated pathogenesis.
6. Benign Tumors
Several benign tumors are known to be associated with the expression of S100A2. Eccrine poromas and syringomas express S100A2 [45], as do apocrine hidrocystomas. Syringocystadenoma papilliferum express S100A2 [45], and calcifying epitheliomas in basophilic cells exhibit positive staining for S100A2 [46].
Craniopharyngioma is a benign tumor that exhibits positive S100A2 expression [47]. The expression of gamma delta T cells is positively associated with S100A2. In contrast, the presence of CD8+ T cells is negatively correlated with S100A2 expression [47], suggesting a possible role of S100A2 in the regulation of immunological reactions.
7. Role of S100A2 in Malignancies
The importance of S100A2 expression has been identified in various malignancies. In this section, we describe the different roles of S100A2 in different organ-derived malignant tumors to better understand the role of S100A2 in tumor development (Figure 2).
8. Brain Cancer
Low-grade glioma is a progressive malignant brain tumor in which S100A2 is upregulated [48]. High S100A2 expression is significantly associated with poor prognosis in patients with low-grade glioma in vivo [48]. Furthermore, S100A2 is also epigenetically regulated by DNA methylation of CpG islands, and high S100A2 expression is related to glioblastoma tumor cell proliferation, apoptosis, invasion, and migration in vivo [49]. S100A2 expression is also related to high tumor grade and is frequently observed in high-clinical-stage gliomas. CpG methylation has been observed in gliomas, and it negatively regulates S100A2 expression in vivo [49]. These findings suggest that elevated S100A2 expression seems to be associated with an unfavorable prognosis in brain-derived tumors.
9. Thyroid Cancer
Papillary carcinoma is the most common type of well-differentiated thyroid malignancy, and the representative oncogenic basis of this tumor is radiation exposure in vivo [50]. Although 89.5% of thyroid papillary carcinomas show positive expression of S100A2 in vivo [51], the detailed prognostic impact of S100A2 in thyroid papillary carcinoma remains unclear.
10. Lung Cancer
Pulmonary fibrosis is a progressive and sometimes fatal disease with an unfavorable prognosis that has the potential to develop into lung cancer in vivo [52]. S100A2 plays an important role in cancer progression. The downregulation of S100A2 suppresses TGF-β1-mediated epithelial–mesenchymal transition (EMT) in lung adenocarcinoma cells in vitro [53]. Furthermore, epidermal growth factor (EGF)-stimulated EGF receptor (EGFR) phosphorylation induces the expression of S100A2 in nonsmall-cell lung carcinoma (NSCLC) in vitro [54]. Consistently, the overexpression of S100A2 in NSCLC tumor cells enhances transendothelial migration and distant organ metastasis in vivo [55].
These findings show that S100A2 may play a dominant role as a positive driver of tumor development in NSCLC. Positive S100A2 expression is significantly related to a high frequency of lymph node metastasis in lung adenocarcinoma in vivo [56]. In addition, high levels of S100A2 mRNA expression are related to poor clinical survival in patients with NSCLC undergoing surgical resection in vivo [57]. High expression of S100A2 is associated with a significantly unfavorable overall survival and disease-specific survival rate after surgery in patients with NSCLC in vivo [58].
In contrast, S100A2 expression is associated with better prognostic outcomes in patients with p53-negative tumors in lung adenocarcinoma in vivo [59], suggesting that S100A2 and other oncogenic factors might be involved in the pathogenesis of NSCLC.
11. Renal Cell Carcinoma
Renal cell carcinoma is the most common kidney cancer in adults, and S100A2 expression is decreased due to the DNA methylation of the promoter site of S100A2 in vitro [60]. Although the prognostic influence remains unclear, lower S100A2 expression in renal cell carcinoma might be expected to be advantageous for tumor development.
12. Liver Diseases
Hepatocellular carcinoma (HCC) is a representative common hepatic cancer, and S100A2 is highly expressed and associated with advanced clinical features in HCC patients in vivo [10,61]. High S100A2 expression is associated with an unfavorable overall survival in vivo [10,62]. As S100A2 expression is upregulated, the hypomethylation status of the CpG island located at the S100A2 promoter site is correlated with the induction of S100A2 expression by HCC cells in vitro [10].
13. Breast Cancer
S100A2 expression decreases in advanced breast cancer in vivo [61]. S100A2 is expressed in 37% of specimens of carcinoma in situ, and in <15% of breast cancer samples in vivo [63], suggesting that the loss of S100A2 is related to the development of breast cancer and does not appear to be with an early phase of tumor development. Highly metastatic breast cancers exhibit elevated levels of S100A2 in vivo [64]. miRNAs are involved in the epigenetic regulation of S100A2 expression in breast cancer cells. miR-325-3p overexpression has similar effects to S100A2 silencing in breast cancer tumor cells in vitro [65]. miR-325-3p overexpression in breast cancer tumor cells enhances tumor cell proliferation and invasion, and EMT development by suppressing S100A2 expression in vitro [65].
14. Bladder Cancer
S100A2 is associated with the development of bladder cancer in vivo [66]. Decreased expression of S100A2 is related to tumor progression and unfavorable clinical outcomes in patients with bladder cancer in vivo [66].
15. Gynecologic Cancers
S100A2 expression is upregulated in patients with ovarian cancer, and high S100A2 expression is associated with advanced clinical stage and unfavorable prognosis in vivo [67,68,69].
S100A2 expression is upregulated in endometrial carcinoma tissues in vivo [70]. Patients with endometrial carcinoma with high S100A2 expression exhibit poor overall survival and disease-specific survival in vivo [70].
16. Pancreatic Cancer
Pancreatic cancer is an aggressive malignant tumor with a high mortality rate despite current advances in therapeutic approaches [71]. S100A2 is associated with immunological phenotypes in the tumor microenvironment of pancreatic cancer. S100A2 expression shows a relationship with lower abundance of CD8+ T cells, activated natural killer (NK) cell infiltration, and a higher abundance of M0 macrophage involvement in vivo [11]. S100A2 is an unfavorable prognostic indicator in pancreatic cancer, and S100A2 expression is positively correlated with the expression of programmed cell death 1-ligand 1 (PD-L1) in pancreatic cancer cells in vivo [11]. Although high S100A2 expression represents an independent predictor of survival in patients with pancreatic cancer, patients with S100A2-negative tumors present favorable survival as patients undergoing pancreatectomy, even in the presence of involved positive surgical margins or lymph node metastases in vivo [72]. Pancreatic adenocarcinoma patients with high S100A2 expression exhibit poor prognosis in vivo [73,74,75]. As a mechanism of downregulating S100A2, the promoter methylation of S100A2 is involved in the development of pancreatic cancer in vitro [74].
17. Melanoma
S100A2 is highly expressed in primary melanoma, whereas its expression is low in metastatic melanoma cells in vivo [76]. S100A2 enhances the antitumor action of treatment with retinoid and thiazolidinedione against melanoma in vitro [77]. Although S100A2 is a ligand of the receptor for advanced glycation end products (RAGEs) [14], which participate in melanoma progression by promoting tumor growth, anti-RAGE antibody treatment reduces tumor growth in vitro [78], S100A2 might have other action points in melanoma to exert antitumor action against melanoma.
18. Prostate Cancer
Lower S100A2 expression is associated with favorable clinical outcomes in prostate cancer in vivo [79,80]. Although benign prostate hypertrophy and prostatitis display increased S100A2 expression, low-grade prostate cancer exhibits reduced expression of S100A2 in vivo [79,80]. DNA methylation has been postulated as a mechanism underlying the downregulation of S100A2 in prostate cancer, and S100A2 methylation has also been observed in 75% of nonmalignant tissues and 100% of benign prostate hypertrophy cases. Immunostaining analysis reveals loss of S100A2 expression in prostate cancer in vivo [81].
19. Squamous Cell Carcinoma
S100A2 expression appears to differ from that at the original site of squamous cell carcinoma. Most squamous cell carcinomas are associated with an unfavorable prognosis, depending on the reduced expression of S100A2. S100A2 expression has been observed in various squamous cell carcinomas, including those of conjunctival origin [82].
S100A2 expression in laryngeal squamous cell carcinoma is positively correlated with longer relapse-free and overall survival [83]. These findings indicate that S100A2 acts as a tumor suppressor. A high frequency of metastasis is found in patients with S100A2-negative oral squamous cell carcinoma tumors in vivo [84].
Another study revealed that, in primary squamous cell carcinoma of the larynx, the high expression of S100A2 appeared to be a significant independent predictive factor for favorable survival in vivo [85]. Another study showed that, in esophageal squamous cell carcinomas, S100A2 expression is significantly higher in well-differentiated tumors and lower lymph node metastasis samples in vivo [86], while S100A2-overexpressing tumor cells exhibit a high frequency of lymph node and distant metastases in esophageal squamous cell carcinoma in vivo [87]. Consistently, another study showed that the expression of S100A2 protein is correlated with tumor differentiation and lymph node metastasis in vivo [88]. Studies of patients with oral squamous cell carcinoma also indicate that S100A2 is related to tumor recurrence in vivo [89]. Cisplatin is an important drug for the treatment of head and neck squamous cell carcinoma in vivo [90]. S100A2 was identified as a potential cisplatin-specific chemoresistance factor in vivo [90]. Silencing S100A2 expression in head and neck squamous cell carcinoma reveals cisplatin sensitivity in acquired and naturally cisplatin-resistant tumor cells in vivo [90].
Regarding S100A2 action in squamous cell carcinoma, S100A2 negatively regulates cell motility in squamous cell carcinoma in vivo [91]. Ectopic expression of S100A2 in the human malignant squamous cell carcinoma cell line KB results in significant inhibition of proliferation, migration, and invasion, which can be negatively regulated by cyclooxygenase-2 (COX-2) expression in vitro [92].
Although it is necessary to clarify the detailed molecular mechanisms involving S100A2 in squamous cell carcinoma, the spectrum of action of S100A2 might have antitumor effects on the development of squamous cell carcinoma.
20. Gastroenterological Cancers
Gastric cancer is a common type of gastrointestinal cancer, and S100A2 expression is downregulated in gastric cancer in vivo [93]. S100A2 downregulation is significantly associated with poor differentiation, tumor invasion, and lymph node metastasis in vivo [93,94,95], and unfavorable survival in patients with gastric cancer in vivo [93,94]. S100A2 acts as a tumor suppressor in gastric cancer, and it suppresses the extracellular-signal-regulated kinase (ERK) and mitogen-activated protein kinase (MAPK)/ERK kinase (MEK) signalling pathway, which is essential for the development of gastric cancer. Consistently, the activation of this signaling pathway by S100A2 downregulation increases tumor invasion in gastric cancer cells in vitro [93].
The overexpression of S100A2 in Barrett’s adenocarcinoma was observed showing a tendency for well-differentiated tumors in vivo [96].
Elevated S100A2 expression is associated with unfavorable clinical survival in colorectal cancer in vivo [97,98,99] and tumor recurrence in vivo [100]. S100A2 overexpression enhances glucose metabolism and cell proliferation owing to the advantage of tumor development in vivo [101]. S100A2 activates the phosphoinositide 3-kinase (PI3K)/AKT serine/threonine kinase (AKT) signaling pathway and upregulates GLUT1 expression, which induces glycolytic reprogramming and consequently increases the proliferation of colorectal cancer cells in vitro [101].
S100A2 expression in cholangiocarcinoma cells is related to a high frequency of lymph node metastasis, advanced clinical stage, and poor patient survival rates in vivo [102].
21. Summary of S100A2 Regulation
The role of S100A2 in inflammatory diseases is largely unknown. However, S100A2 is expected to be a positive driver for the development of inflammatory reactions in atopic dermatitis, psoriasis, drug eruption, and asthma.
High S100A2 expression is associated with an unfavorable clinical course in brain tumors, such as low-grade glioma and glioblastoma, NSCLC, hepatocellular carcinoma, ovarian cancer, endometrial carcinoma, pancreatic cancer, prostate cancer, colorectal cancer, and cholangiocarcinoma. In contrast, lower S100A2 expression is associated with an unfavorable clinical course in patients with renal cell carcinoma, breast cancer, bladder cancer, melanoma, and gastric cancer. These findings indicate that the prognostic impact of S100A2 is completely different in each malignancy. As one of the reasons, S100A2 was a responsible for tumor cell migration in some tumor cells in vitro [91], suggesting that A100S2 might have different unknown action mechanisms in each tumor.
The role of S100A2 in benign tumors is also unknown; however, its expression can also be used as a marker to distinguish it from other benign or malignant tumors.
22. Conclusions
S100A2 expression and its roles are completely different in different diseases. Our review shows that S100A2 functions as a tumor suppressor; however, the influence of its expression is completely different in different types of malignant tumors. In particular, there exists a need to elucidate expression alterations of S100A2 in early and advanced stages of malignant tumors, in addition to determining the actual impact of S100A2 on their prognosis. Despite the analysis of S100A2 in cancers, there are only a few studies focused on inflammatory diseases and benign tumors. In particular, the immunomodulatory action of S100A2 has been speculated upon in previous studies, and further investigation is necessary to clarify the actual role of immunomodulatory functions of S100A2 in inflammatory diseases. In addition, a limited number of studies reported the role of A100A2 as a biomarker in malignancies and inflammatory diseases to estimate the prognosis or disease severity [12,103].
Taken together, we summarized the current trend of the knowledge gained from recent studies that showed a clear role of S100A2 in certain malignancies; however, the unknown mechanisms of S100A2 should be elucidated by further analysis in the future.
This figure shows the regulatory mechanism and intracation factors to enhance the action of S100A2 in physiological and pathological conditions. Various factors are involved in S100A2 regulation for physiological and pathological actions. TGF-β, TNF-α, and p53 can enhance the induction of S100A. S100A2 is also regulated by epigenetic modifications. DNA methylation impairs S100A2 expression, while histone methylation accelerates S100A2 gene expression. S100A2 also involves interaction with other molecules, such as S100A4, EGFR, DeltaNp63, and erythropoietin for the enhancement of the ability of S100A2 function.
The prognostic impact or the severity of influences is summarized in this figure. Although S100A2 has a tumor suppressor function, the recently updated research focusing on the prognostic impact of S100A2 revealed the various influences of S100A2 in human malignancies in addition to inflammatory diseases. High S100A2 expression was associated with a better prognosis in patients with breast cancer, bladder cancer, melanoma, laryngeal squamous cell carcinoma, and gastric cancer. On the other hand, a high expression of S100A2 was associated with a poorer prognosis in patients with nonsmall-cell lung cancer, hepatocellular carcinoma, ovarian cancer, endometrial carcinoma, pancreatic cancer, prostate cancer, oral squamous cell carcinoma, colorectal cancer, cholangiocarcinoma. S100A2 was also involved in the development of inflammatory reactions, atopic dermatitis, drug eruption, psoriasis, delayed wound healing, and asthma.
Author Contributions
H.S. and Y.S. wrote and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Y.S. received a basic research grant from the Japanese Dermatology Association (Shiseido donated).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Marenholz, I.; Heizmann, C.W.; Fritz, G. S100 proteins in mouse and man: From evolution to function and pathology (including an update of the nomenclature). Biochem. Biophys. Res. Commun. 2004, 322, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Marks, A.; Allore, R. S100 protein and Down syndrome. Bioessays 1990, 12, 381–383. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, T.; Samson, J. S-100 protein in normal and pathologic oral tissues. A review. Schweiz Mon. Zahnmed 1993, 103, 413–418. [Google Scholar]
- Zimmer, D.B.; Cornwall, E.H.; Landar, A.; Song, W. The S100 protein family: History, function, and expression. Brain Res. Bull. 1995, 37, 417–429. [Google Scholar] [CrossRef]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef]
- Eckert, R.L.; Broome, A.M.; Ruse, M.; Robinson, N.; Ryan, D.; Lee, K. S100 proteins in the epidermis. J. Investig. Derm. 2004, 123, 23–33. [Google Scholar] [CrossRef]
- Koch, M.; Bhattacharya, S.; Kehl, T.; Gimona, M.; Vasák, M.; Chazin, W.; Heizmann, C.W.; Kroneck, P.M.; Fritz, G. Implications on zinc binding to S100A2. Biochim. Biophys. Acta 2007, 1773, 457–470. [Google Scholar] [CrossRef][Green Version]
- Wolf, S.; Haase-Kohn, C.; Pietzsch, J. S100A2 in cancerogenesis: A friend or a foe? Amino. Acids 2011, 41, 849–861. [Google Scholar] [CrossRef]
- Wicki, R.; Franz, C.; Scholl, F.A.; Heizmann, C.W.; Schäfer, B.W. Repression of the candidate tumor suppressor gene S100A2 in breast cancer is mediated by site-specific hypermethylation. Cell Calcium. 1997, 22, 243–254. [Google Scholar] [CrossRef]
- Yan, J.; Huang, Y.J.; Huang, Q.Y.; Liu, P.X.; Wang, C.S. Transcriptional activation of S100A2 expression by HIF-1α via binding to the hypomethylated hypoxia response elements in HCC cells. Mol. Carcinog. 2022, 61, 494–507. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, C.; Song, J.; Xu, R.; Ruze, R.; Zhao, Y. S100A2 Is a Prognostic Biomarker Involved in Immune Infiltration and Predict Immunotherapy Response in Pancreatic Cancer. Front. Immunol. 2021, 12, 758004. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Sawada, Y.; Saito-Sasaki, N.; Yoshioka, H.; Hama, K.; Omoto, D.; Ohmori, S.; Okada, E.; Nakamura, M. High S100A2 expression in keratinocytes in patients with drug eruption. Sci. Rep. 2021, 11, 5493. [Google Scholar] [CrossRef] [PubMed]
- Hountis, P.; Matthaios, D.; Froudarakis, M.; Bouros, D.; Kakolyris, S. S100A2 protein and non-small cell lung cancer. The dual role concept. Tumour. Biol. 2014, 35, 7327–7333. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, R.; Woods, T.L.; Fu, J.; Zhang, T.; Stoll, S.W.; Elder, J.T. Biochemical characterization of S100A2 in human keratinocytes: Subcellular localization, dimerization, and oxidative cross-linking. J. Investig. Derm. 2000, 115, 477–485. [Google Scholar] [CrossRef]
- Zhang, T.; Woods, T.L.; Elder, J.T. Differential responses of S100A2 to oxidative stress and increased intracellular calcium in normal, immortalized, and malignant human keratinocytes. J. Investig. Derm. 2002, 119, 1196–1201. [Google Scholar] [CrossRef][Green Version]
- Naz, S.; Ranganathan, P.; Bodapati, P.; Shastry, A.H.; Mishra, L.N.; Kondaiah, P. Regulation of S100A2 expression by TGF-β-induced MEK/ERK signalling and its role in cell migration/invasion. Biochem. J. 2012, 447, 81–91. [Google Scholar] [CrossRef]
- Foser, S.; Redwanz, I.; Ebeling, M.; Heizmann, C.W.; Certa, U. Interferon-alpha and transforming growth factor-beta co-induce growth inhibition of human tumor cells. Cell Mol. Life Sci. 2006, 63, 2387–2396. [Google Scholar] [CrossRef]
- Haase-Kohn, C.; Wolf, S.; Lenk, J.; Pietzsch, J. Copper-mediated cross-linking of S100A4, but not of S100A2, results in proinflammatory effects in melanoma cells. Biochem. Biophys. Res. Commun. 2011, 413, 494–498. [Google Scholar] [CrossRef]
- Nagy, N.; Hoyaux, D.; Gielen, I.; Schäfer, B.W.; Pochet, R.; Heizmann, C.W.; Kiss, R.; Salmon, I.; Decaestecker, C. The Ca2+-binding S100A2 protein is differentially expressed in epithelial tissue of glandular or squamous origin. Histol. Histopathol. 2002, 17, 123–130. [Google Scholar]
- Stoll, S.W.; Zhao, X.; Elder, J.T. EGF stimulates transcription of CaN19 (S100A2) in HaCaT keratinocytes. J. Investig. Derm. 1998, 111, 1092–1097. [Google Scholar] [CrossRef]
- Tan, M.; Heizmann, C.W.; Guan, K.; Schafer, B.W.; Sun, Y. Transcriptional activation of the human S100A2 promoter by wild-type p53. FEBS Lett. 1999, 445, 265–268. [Google Scholar] [CrossRef]
- Koch, M.; Diez, J.; Wagner, A.; Fritz, G. Crystallization and calcium/sulfur SAD phasing of the human EF-hand protein S100A2. Acta Cryst. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 1032–1036. [Google Scholar] [CrossRef]
- van Dieck, J.; Fernandez-Fernandez, M.R.; Veprintsev, D.B.; Fersht, A.R. Modulation of the oligomerization state of p53 by differential binding of proteins of the S100 family to p53 monomers and tetramers. J. Biol. Chem. 2009, 284, 13804–13811. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, R.D.; Sänger, K.; Müller, G.A.; Engeland, K. Transcriptional activation of the tumor suppressor and differentiation gene S100A2 by a novel p63-binding site. Nucleic. Acids Res. 2008, 36, 2969–2980. [Google Scholar] [CrossRef] [PubMed]
- Lapi, E.; Iovino, A.; Fontemaggi, G.; Soliera, A.R.; Iacovelli, S.; Sacchi, A.; Rechavi, G.; Givol, D.; Blandino, G.; Strano, S. S100A2 gene is a direct transcriptional target of p53 homologues during keratinocyte differentiation. Oncogene 2006, 25, 3628–3637. [Google Scholar] [CrossRef][Green Version]
- Buckley, N.E.; D’Costa, Z.; Kaminska, M.; Mullan, P.B. S100A2 is a BRCA1/p63 coregulated tumour suppressor gene with roles in the regulation of mutant p53 stability. Cell Death Dis. 2014, 5, e1070. [Google Scholar] [CrossRef]
- Kazakov, A.S.; Deryusheva, E.I.; Sokolov, A.S.; Permyakova, M.E.; Litus, E.A.; Rastrygina, V.A.; Uversky, V.N.; Permyakov, E.A.; Permyakov, S.E. Erythropoietin Interacts with Specific S100 Proteins. Biomolecules 2022, 12, 120. [Google Scholar] [CrossRef]
- Sawada, Y.; Gallo, R.L. Role of Epigenetics in the Regulation of Immune Functions of the Skin. J. Investig. Derm. 2021, 141, 1157–1166. [Google Scholar] [CrossRef]
- Seale, K.; Horvath, S.; Teschendorff, A.; Eynon, N.; Voisin, S. Making sense of the ageing methylome. Nat. Rev. Genet. 2022, in press. [Google Scholar] [CrossRef]
- Tsou, P.S.; Varga, J.; O’Reilly, S. Advances in epigenetics in systemic sclerosis: Molecular mechanisms and therapeutic potential. Nat. Rev. Rheumatol. 2021, 17, 596–607. [Google Scholar] [CrossRef]
- Dor, Y.; Cedar, H. Principles of DNA methylation and their implications for biology and medicine. Lancet 2018, 392, 777–786. [Google Scholar] [CrossRef]
- El-Dahr, S.S. DNA methylation links intrauterine stress with abnormal nephrogenesis. Nat. Rev. Nephrol. 2019, 15, 196–197. [Google Scholar] [CrossRef] [PubMed]
- Michalak, E.M.; Burr, M.L.; Bannister, A.J.; Dawson, M.A. The roles of DNA, RNA and histone methylation in ageing and cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 573–589. [Google Scholar] [CrossRef]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef]
- Nanamori, H.; Sawada, Y. Epigenetic Modification of PD-1/PD-L1-Mediated Cancer Immunotherapy against Melanoma. Int. J. Mol. Sci. 2022, 23, 1119. [Google Scholar] [CrossRef]
- Lorincz, M.C.; Schübeler, D. Evidence for Converging DNA Methylation Pathways in Placenta and Cancer. Dev. Cell 2017, 43, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Xu, X.; Youssef, E.M.; Lotan, R. Diminished expression of S100A2, a putative tumor suppressor, at early stage of human lung carcinogenesis. Cancer Res. 2001, 61, 7999–8004. [Google Scholar]
- Lee, J.; Wysocki, P.T.; Topaloglu, O.; Maldonado, L.; Brait, M.; Begum, S.; Moon, D.; Kim, M.S.; Califano, J.A.; Sidransky, D.; et al. Epigenetic silencing of S100A2 in bladder and head and neck cancers. Oncoscience 2015, 2, 410–418. [Google Scholar] [CrossRef][Green Version]
- Kim, Y.; Jeong, Y.; Kwon, K.; Ismail, T.; Lee, H.K.; Kim, C.; Park, J.W.; Kwon, O.S.; Kang, B.S.; Lee, D.S.; et al. Physiological effects of KDM5C on neural crest migration and eye formation during vertebrate development. Epigenetics Chromatin 2018, 11, 72. [Google Scholar] [CrossRef]
- Wang, Y.; Ye, H.; Yang, Y.; Li, J.; Cen, A.; Zhao, L. microRNA-181a promotes the oncogene S100A2 and enhances papillary thyroid carcinoma growth by mediating the expression of histone demethylase KDM5C. J. Endocrinol. Investig. 2022, 45, 17–28. [Google Scholar] [CrossRef]
- Pan, S.C.; Li, C.Y.; Kuo, C.Y.; Kuo, Y.Z.; Fang, W.Y.; Huang, Y.H.; Hsieh, T.C.; Kao, H.Y.; Kuo, Y.; Kang, Y.R.; et al. The p53-S100A2 Positive Feedback Loop Negatively Regulates Epithelialization in Cutaneous Wound Healing. Sci. Rep. 2018, 8, 5458. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, C.; Kohda, F.; Mizote, Y.; Miake, A.; Ijichi, A.; Kawahara, S.; Kohno, M.; Sonoyama, H.; Mitamura, Y.; Kaku, Y.; et al. Localization of S100A2, S100A4, S100A6, S100A7, and S100P in the human hair follicle. Fukuoka Igaku Zasshi 2014, 105, 148–156. [Google Scholar] [PubMed]
- Gliga, A.R.; Di Bucchianico, S.; Åkerlund, E.; Karlsson, H.L. Transcriptome Profiling and Toxicity Following Long-Term, Low Dose Exposure of Human Lung Cells to Ni and NiO Nanoparticles-Comparison with NiCl(2). Nanomaterials 2020, 10, 649. [Google Scholar] [CrossRef] [PubMed]
- Poachanukoon, O.; Roytrakul, S.; Koontongkaew, S. A shotgun proteomic approach reveals novel potential salivary protein biomarkers for asthma. J. Asthma. 2022, 59, 243–254. [Google Scholar] [CrossRef]
- Zhu, L.; Okano, S.; Takahara, M.; Chiba, T.; Tu, Y.; Oda, Y.; Furue, M. Expression of S100 protein family members in normal skin and sweat gland tumors. J. Derm. Sci. 2013, 70, 211–219. [Google Scholar] [CrossRef]
- Shrestha, P.; Muramatsu, Y.; Kudeken, W.; Mori, M.; Takai, Y.; Ilg, E.C.; Schafer, B.W.; Heizmann, C.W. Localization of Ca(2+)-binding S100 proteins in epithelial tumours of the skin. Virchows Arch. 1998, 432, 53–59. [Google Scholar] [CrossRef]
- Lin, D.; Zhao, W.; Yang, J.; Wang, H.; Zhang, H. Integrative Analysis of Biomarkers and Mechanisms in Adamantinomatous Craniopharyngioma. Front. Genet. 2022, 13, 830793. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, X.; Zhu, X.L.; Bai, H.; Wang, Z.Z.; Zhang, J.J.; Hao, C.Y.; Duan, H.B. S100A gene family: Immune-related prognostic biomarkers and therapeutic targets for low-grade glioma. Aging 2021, 13, 15459–15478. [Google Scholar] [CrossRef]
- Yin, A.; Shang, Z.; Etcheverry, A.; He, Y.; Aubry, M.; Lu, N.; Liu, Y.; Mosser, J.; Lin, W.; Zhang, X.; et al. Integrative analysis identifies an immune-relevant epigenetic signature for prognostication of non-G-CIMP glioblastomas. Oncoimmunology 2021, 10, 1902071. [Google Scholar] [CrossRef]
- Aljohani, A.O.; Merdad, R.H.; Alserif, A.I.; Alhemayed, L.S.; Farsi, N.Z.; Alsufyani, T.A.; Alkaf, H.H.; Alherabi, A.Z.; Abdelmonim, S.K.; Alessa, M.A. The impact of thyroid tumor features on lymph node metastasis in papillary thyroid carcinoma patients in head and neck department at KAMC: A retrospective cross-sectional study. Ann. Med. Surg. 2021, 64, 102217. [Google Scholar] [CrossRef]
- Ito, Y.; Yoshida, H.; Tomoda, C.; Uruno, T.; Miya, A.; Kobayashi, K.; Matsuzuka, F.; Kakudo, K.; Kuma, K.; Miyauchi, A. Expression of S100A2 and S100A6 in thyroid carcinomas. Histopathology 2005, 46, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Lederer, D.J.; Martinez, F.J. Idiopathic Pulmonary Fibrosis. N. Engl. J. Med. 2018, 378, 1811–1823. [Google Scholar] [CrossRef]
- Huang, G.; Zhang, J.; Qing, G.; Liu, D.; Wang, X.; Chen, Y.; Li, Y.; Guo, S. S100A2 Silencing Relieves Epithelial-Mesenchymal Transition in Pulmonary Fibrosis by Inhibiting the Wnt/β-Catenin Signaling Pathway. DNA Cell Biol. 2021, 40, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yun, S.T.; Yun, C.O.; Ahn, B.Y.; Jo, E.C. S100A2 promoter-driven conditionally replicative adenovirus targets non-small-cell lung carcinoma. Gene Ther. 2012, 19, 967–977. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Diederichs, S.; Bulk, E.; Steffen, B.; Ji, P.; Tickenbrock, L.; Lang, K.; Zänker, K.S.; Metzger, R.; Schneider, P.M.; Gerke, V.; et al. S100 family members and trypsinogens are predictors of distant metastasis and survival in early-stage non-small cell lung cancer. Cancer Res. 2004, 64, 5564–5569. [Google Scholar] [CrossRef]
- Wang, T.; Liang, Y.; Thakur, A.; Zhang, S.; Liu, F.; Khan, H.; Shi, P.; Wang, N.; Chen, M.; Ren, H. Expression and clinicopathological significance of S100 calcium binding protein A2 in lung cancer patients of Chinese Han ethnicity. Clin. Chim. Acta 2017, 464, 118–122. [Google Scholar] [CrossRef]
- Bulk, E.; Sargin, B.; Krug, U.; Hascher, A.; Jun, Y.; Knop, M.; Kerkhoff, C.; Gerke, V.; Liersch, R.; Mesters, R.M.; et al. S100A2 induces metastasis in non-small cell lung cancer. Clin. Cancer Res. 2009, 15, 22–29. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Z.; Li, R.; Ang, K.K.; Zhang, H.; Caraway, N.P.; Katz, R.L.; Jiang, F. Overexpression of S100A2 protein as a prognostic marker for patients with stage I non small cell lung cancer. Int. J. Cancer 2005, 116, 285–290. [Google Scholar] [CrossRef]
- Matsubara, D.; Niki, T.; Ishikawa, S.; Goto, A.; Ohara, E.; Yokomizo, T.; Heizmann, C.W.; Aburatani, H.; Moriyama, S.; Moriyama, H.; et al. Differential expression of S100A2 and S100A4 in lung adenocarcinomas: Clinicopathological significance, relationship to p53 and identification of their target genes. Cancer Sci. 2005, 96, 844–857. [Google Scholar] [CrossRef]
- Sugiyama, T.; Ozono, S.; Miyake, H. Expression Profile of S100A2 and its Clinicopathological Significance in Renal Cell Carcinoma. Anticancer Res. 2020, 40, 6337–6343. [Google Scholar] [CrossRef]
- Zheng, S.; Liu, L.; Xue, T.; Jing, C.; Xu, X.; Wu, Y.; Wang, M.; Xie, X.; Zhang, B. Comprehensive Analysis of the Prognosis and Correlations With Immune Infiltration of S100 Protein Family Members in Hepatocellular Carcinoma. Front. Genet. 2021, 12, 648156. [Google Scholar] [CrossRef]
- Zhang, C.; Yao, R.; Chen, J.; Zou, Q.; Zeng, L. S100 family members: Potential therapeutic target in patients with hepatocellular carcinoma: A STROBE study. Medicine 2021, 100, e24135. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Rudland, P.S.; Sibson, D.R.; Platt-Higgins, A.; Barraclough, R. Expression of calcium-binding protein S100A2 in breast lesions. Br. J. Cancer 2000, 83, 1473–1479. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Lamar, J.M.; Carr, S.A.; Hynes, R.O. Extracellular matrix signatures of human mammary carcinoma identify novel metastasis promoters. eLife 2014, 3, e01308. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, X.; Yang, F.; Xiao, H. miR-325-3p Promotes the Proliferation, Invasion, and EMT of Breast Cancer Cells by Directly Targeting S100A2. Oncol. Res. 2021, 28, 731–744. [Google Scholar] [CrossRef]
- Matsumoto, K.; Irie, A.; Satoh, T.; Ishii, J.; Iwabuchi, K.; Iwamura, M.; Egawa, S.; Baba, S. Expression of S100A2 and S100A4 predicts for disease progression and patient survival in bladder cancer. Urology 2007, 70, 602–607. [Google Scholar] [CrossRef]
- Xu, H.Y.; Song, H.M.; Zhou, Q. Comprehensive analysis of the expression and prognosis for S100 in human ovarian cancer: A STROBE study. Medicine 2020, 99, e22777. [Google Scholar] [CrossRef]
- Lou, W.; Ding, B.; Zhong, G.; Du, C.; Fan, W.; Fu, P. Dysregulation of pseudogene/lncRNA-hsa-miR-363-3p-SPOCK2 pathway fuels stage progression of ovarian cancer. Aging 2019, 11, 11416–11439. [Google Scholar] [CrossRef]
- Bai, Y.; Li, L.D.; Li, J.; Lu, X. Prognostic values of S100 family members in ovarian cancer patients. BMC Cancer 2018, 18, 1256. [Google Scholar] [CrossRef]
- Zhang, Q.; Xia, T.; Qi, C.; Du, J.; Ye, C. High expression of S100A2 predicts poor prognosis in patients with endometrial carcinoma. BMC Cancer 2022, 22, 77. [Google Scholar] [CrossRef]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Biankin, A.V.; Kench, J.G.; Colvin, E.K.; Segara, D.; Scarlett, C.J.; Nguyen, N.Q.; Chang, D.K.; Morey, A.L.; Lee, C.S.; Pinese, M.; et al. Expression of S100A2 calcium-binding protein predicts response to pancreatectomy for pancreatic cancer. Gastroenterology 2009, 137, 558–568.e11. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qiu, N.; Li, Q. Prognostic Values and Clinical Significance of S100 Family Member’s Individualized mRNA Expression in Pancreatic Adenocarcinoma. Front. Genet. 2021, 12, 758725. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Wang, J.L.; Jin, X.D.; Zhao, L.; Ye, H.L.; Kuang, Y.B.; Ma, Y.; Jiang, X.Y.; Yu, Z.Y. Comprehensive analysis of the transcriptional expressions and prognostic value of S100A family in pancreatic ductal adenocarcinoma. BMC Cancer 2021, 21, 1039. [Google Scholar] [CrossRef]
- Bachet, J.B.; Maréchal, R.; Demetter, P.; Bonnetain, F.; Cros, J.; Svrcek, M.; Bardier-Dupas, A.; Hammel, P.; Sauvanet, A.; Louvet, C.; et al. S100A2 is a predictive biomarker of adjuvant therapy benefit in pancreatic adenocarcinoma. Eur. J. Cancer 2013, 49, 2643–2653. [Google Scholar] [CrossRef]
- Xiong, T.F.; Pan, F.Q.; Li, D. Expression and clinical significance of S100 family genes in patients with melanoma. Melanoma Res. 2019, 29, 23–29. [Google Scholar] [CrossRef]
- Klopper, J.P.; Sharma, V.; Bissonnette, R.; Haugen, B.R. Combination PPARgamma and RXR Agonist Treatment in Melanoma Cells: Functional Importance of S100A2. PPAR Res. 2010, 2010, 729876. [Google Scholar] [CrossRef]
- Meghnani, V.; Wagh, A.; Indurthi, V.S.; Koladia, M.; Vetter, S.W.; Law, B.; Leclerc, E. The receptor for advanced glycation end products influences the expression of its S100 protein ligands in melanoma tumors. Int. J. Biochem. Cell Biol. 2014, 57, 54–62. [Google Scholar] [CrossRef]
- Gupta, S.; Hussain, T.; MacLennan, G.T.; Fu, P.; Patel, J.; Mukhtar, H. Differential expression of S100A2 and S100A4 during progression of human prostate adenocarcinoma. J. Clin. Oncol. 2003, 21, 106–112. [Google Scholar] [CrossRef]
- Kwon, Y.W.; Chang, I.H.; Kim, K.D.; Kim, Y.S.; Myung, S.C.; Kim, M.K.; Kim, T.H. Significance of S100A2 and S100A4 Expression in the Progression of Prostate Adenocarcinoma. Korean J. Urol. 2010, 51, 456–462. [Google Scholar] [CrossRef][Green Version]
- Rehman, I.; Cross, S.S.; Catto, J.W.; Leiblich, A.; Mukherjee, A.; Azzouzi, A.R.; Leung, H.Y.; Hamdy, F.C. Promoter hyper-methylation of calcium binding proteins S100A6 and S100A2 in human prostate cancer. Prostate 2005, 65, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Mahale, A.; Alkatan, H.; Alwadani, S.; Othman, M.; Suarez, M.J.; Price, A.; Al-Hussain, H.; Jastaneiah, S.; Yu, W.; Maktabi, A.; et al. Altered gene expression in conjunctival squamous cell carcinoma. Mod. Pathol. 2016, 29, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Lauriola, L.; Michetti, F.; Maggiano, N.; Galli, J.; Cadoni, G.; Schäfer, B.W.; Heizmann, C.W.; Ranelletti, F.O. Prognostic significance of the Ca(2+) binding protein S100A2 in laryngeal squamous-cell carcinoma. Int. J. Cancer 2000, 89, 345–349. [Google Scholar] [CrossRef]
- Suzuki, F.; Oridate, N.; Homma, A.; Nakamaru, Y.; Nagahashi, T.; Yagi, K.; Yamaguchi, S.; Furuta, Y.; Fukuda, S. S100A2 expression as a predictive marker for late cervical metastasis in stage I and II invasive squamous cell carcinoma of the oral cavity. Oncol. Rep. 2005, 14, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Almadori, G.; Bussu, F.; Galli, J.; Rigante, M.; Lauriola, L.; Michetti, F.; Maggiano, N.; Schafer, B.W.; Heizmann, C.W.; Ranelletti, F.O.; et al. Diminished expression of S100A2, a putative tumour suppressor, is an independent predictive factor of neck node relapse in laryngeal squamous cell carcinoma. J. Otolaryngol. Head Neck Surg. 2009, 38, 16–22. [Google Scholar] [PubMed]
- Kyriazanos, I.D.; Tachibana, M.; Dhar, D.K.; Shibakita, M.; Ono, T.; Kohno, H.; Nagasue, N. Expression and prognostic significance of S100A2 protein in squamous cell carcinoma of the esophagus. Oncol. Rep. 2002, 9, 503–510. [Google Scholar] [CrossRef]
- Imazawa, M.; Hibi, K.; Fujitake, S.; Kodera, Y.; Ito, K.; Akiyama, S.; Nakao, A. S100A2 overexpression is frequently observed in esophageal squamous cell carcinoma. Anticancer Res. 2005, 25, 1247–1250. [Google Scholar]
- Cao, L.Y.; Yin, Y.; Li, H.; Jiang, Y.; Zhang, H.F. Expression and clinical significance of S100A2 and p63 in esophageal carcinoma. World J. Gastroenterol. 2009, 15, 4183–4188. [Google Scholar] [CrossRef]
- Kumar, M.; Srivastava, G.; Kaur, J.; Assi, J.; Alyass, A.; Leong, I.; MacMillan, C.; Witterick, I.; Shukla, N.K.; Thakar, A.; et al. Prognostic significance of cytoplasmic S100A2 overexpression in oral cancer patients. J. Transl. Med. 2015, 13, 8. [Google Scholar] [CrossRef]
- Inukai, D.; Nishimura, K.; Okamoto, H.; Sano, R.; Ueda, H.; Ota, A.; Karnan, S.; Hosokawa, Y.; Yoshikawa, K.; Suzuki, S.; et al. Identification of cisplatin-resistant factor by integration of transcriptomic and proteomic data using head and neck carcinoma cell lines. Nagoya J. Med. Sci. 2020, 82, 519–531. [Google Scholar]
- Nagy, N.; Brenner, C.; Markadieu, N.; Chaboteaux, C.; Camby, I.; Schäfer, B.W.; Pochet, R.; Heizmann, C.W.; Salmon, I.; Kiss, R.; et al. S100A2, a putative tumor suppressor gene, regulates in vitro squamous cell carcinoma migration. Lab. Investig. 2001, 81, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Tsai, S.T.; Jin, Y.T.; Wu, L.W. Cyclooxygenase-2 is involved in S100A2-mediated tumor suppression in squamous cell carcinoma. Mol. Cancer Res. 2006, 4, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Liu, Q.Q.; Wang, X.; Luo, C.H. Clinical significance of S100A2 expression in gastric cancer. Tumour Biol. 2014, 35, 3731–3741. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, T.B.; Wang, Q. Clinical significance of altered S100A2 expression in gastric cancer. Oncol. Rep. 2013, 29, 1556–1562. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Zhu, Y.; Yang, G.; Gong, L.; Wang, B.; Liu, H. Loss of Reprimo and S100A2 expression in human gastric adenocarcinoma. Diagn. Cytopathol. 2011, 39, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Lee, O.J.; Hong, S.M.; Razvi, M.H.; Peng, D.; Powell, S.M.; Smoklin, M.; Moskaluk, C.A.; El-Rifai, W. Expression of calcium-binding proteins S100A2 and S100A4 in Barrett’s adenocarcinomas. Neoplasia 2006, 8, 843–850. [Google Scholar] [CrossRef][Green Version]
- Masuda, T.; Ishikawa, T.; Mogushi, K.; Okazaki, S.; Ishiguro, M.; Iida, S.; Mizushima, H.; Tanaka, H.; Uetake, H.; Sugihara, K. Overexpression of the S100A2 protein as a prognostic marker for patients with stage II and III colorectal cancer. Int. J. Oncol. 2016, 48, 975–982. [Google Scholar] [CrossRef]
- Long, N.P.; Park, S.; Anh, N.H.; Nghi, T.D.; Yoon, S.J.; Park, J.H.; Lim, J.; Kwon, S.W. High-Throughput Omics and Statistical Learning Integration for the Discovery and Validation of Novel Diagnostic Signatures in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 296. [Google Scholar] [CrossRef]
- Alajez, N.M. Large-Scale Analysis of Gene Expression Data Reveals a Novel Gene Expression Signature Associated with Colorectal Cancer Distant Recurrence. PLoS ONE 2016, 11, e0167455. [Google Scholar] [CrossRef]
- Giráldez, M.D.; Lozano, J.J.; Cuatrecasas, M.; Alonso-Espinaco, V.; Maurel, J.; Mármol, M.; Hörndler, C.; Ortego, J.; Alonso, V.; Escudero, P.; et al. Gene-expression signature of tumor recurrence in patients with stage II and III colon cancer treated with 5’fluoruracil-based adjuvant chemotherapy. Int. J. Cancer 2013, 132, 1090–1097. [Google Scholar] [CrossRef]
- Li, C.; Chen, Q.; Zhou, Y.; Niu, Y.; Wang, X.; Li, X.; Zheng, H.; Wei, T.; Zhao, L.; Gao, H. S100A2 promotes glycolysis and proliferation via GLUT1 regulation in colorectal cancer. FASEB J. 2020, 34, 13333–13344. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Harada, K.; Sasaki, M.; Nakanuma, Y. Clinicopathological significance of S100 protein expression in cholangiocarcinoma. J. Gastroenterol. Hepatol. 2013, 28, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.T.; Jin, Y.T.; Tsai, W.C.; Wang, S.T.; Lin, Y.C.; Chang, M.T.; Wu, L.W. S100A2, a potential marker for early recurrence in early-stage oral cancer. Oral. Oncol. 2005, 41, 349–357. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).