Identification of Regulatory Host Genes Involved in Sigma Virus Replication Using RNAi Knockdown in Drosophila


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Flies, Virus Stock, and CO2 Assay
2.2. dsRNA Preparation
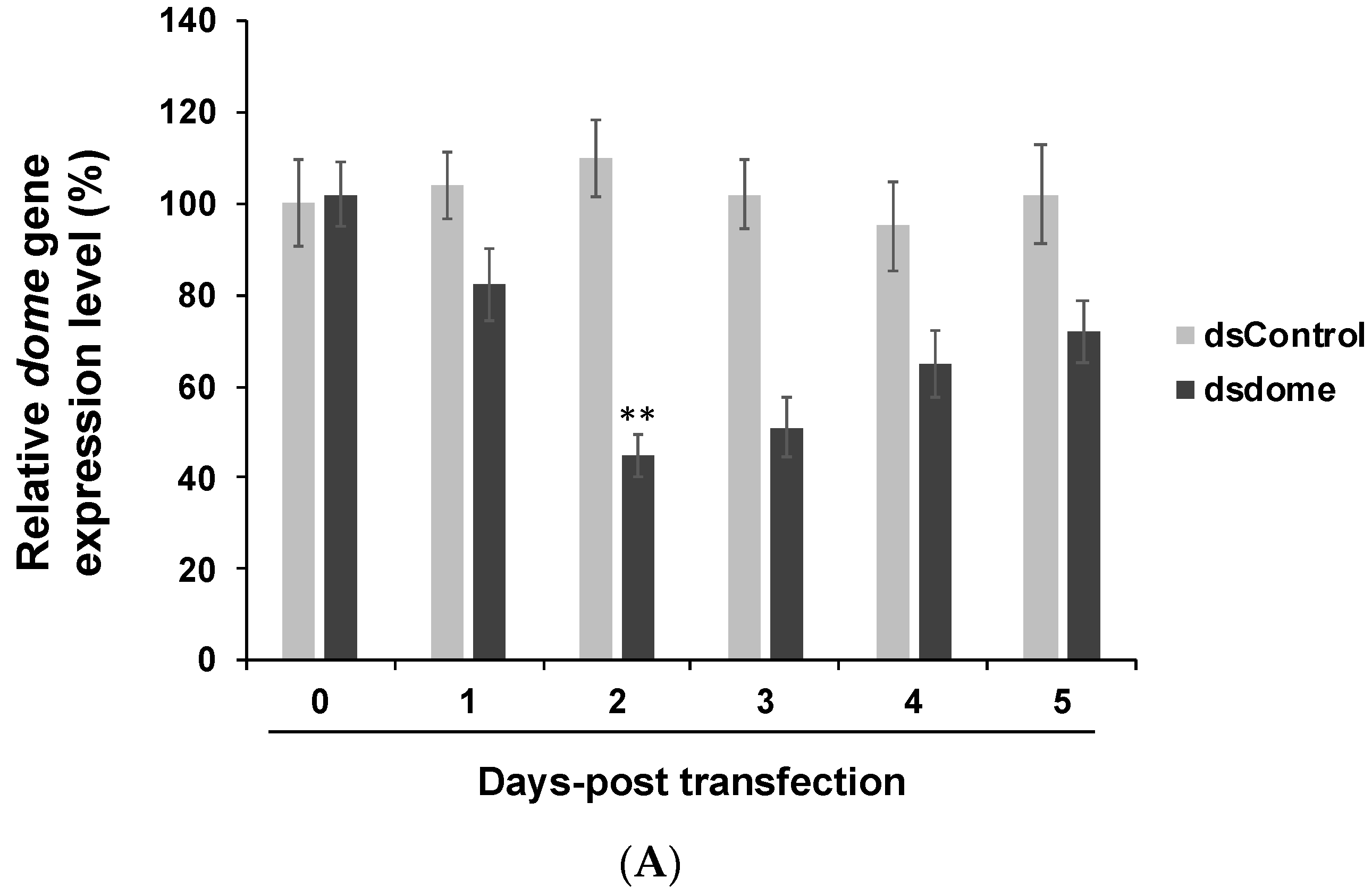
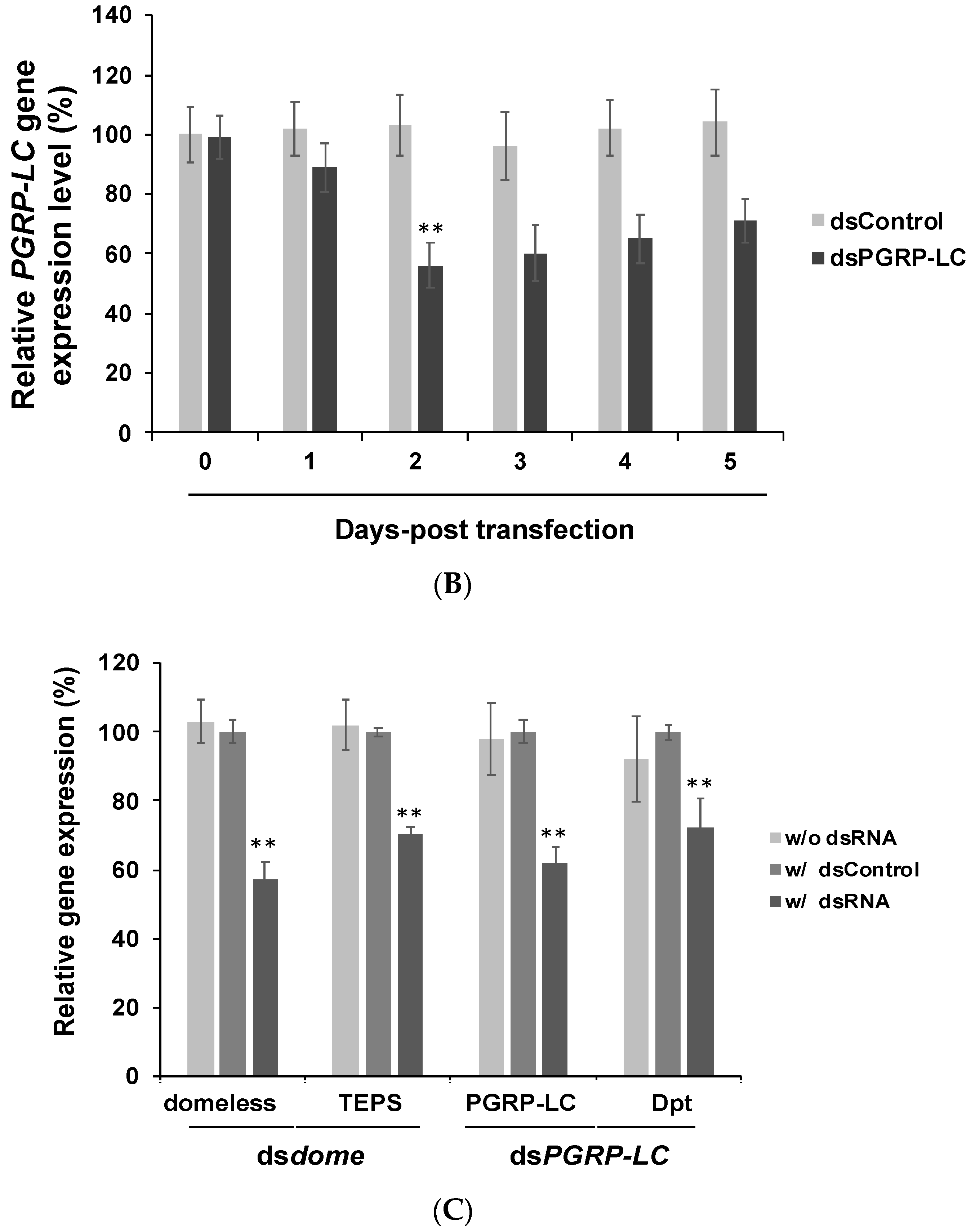
2.3. dsRNA Transfection, Virus Infection, and RNA Extraction
2.4. Gene Expression Assay Using RT-qPCR
2.5. dsRNA Microinjection
2.6. Statistical Analysis
3. Results
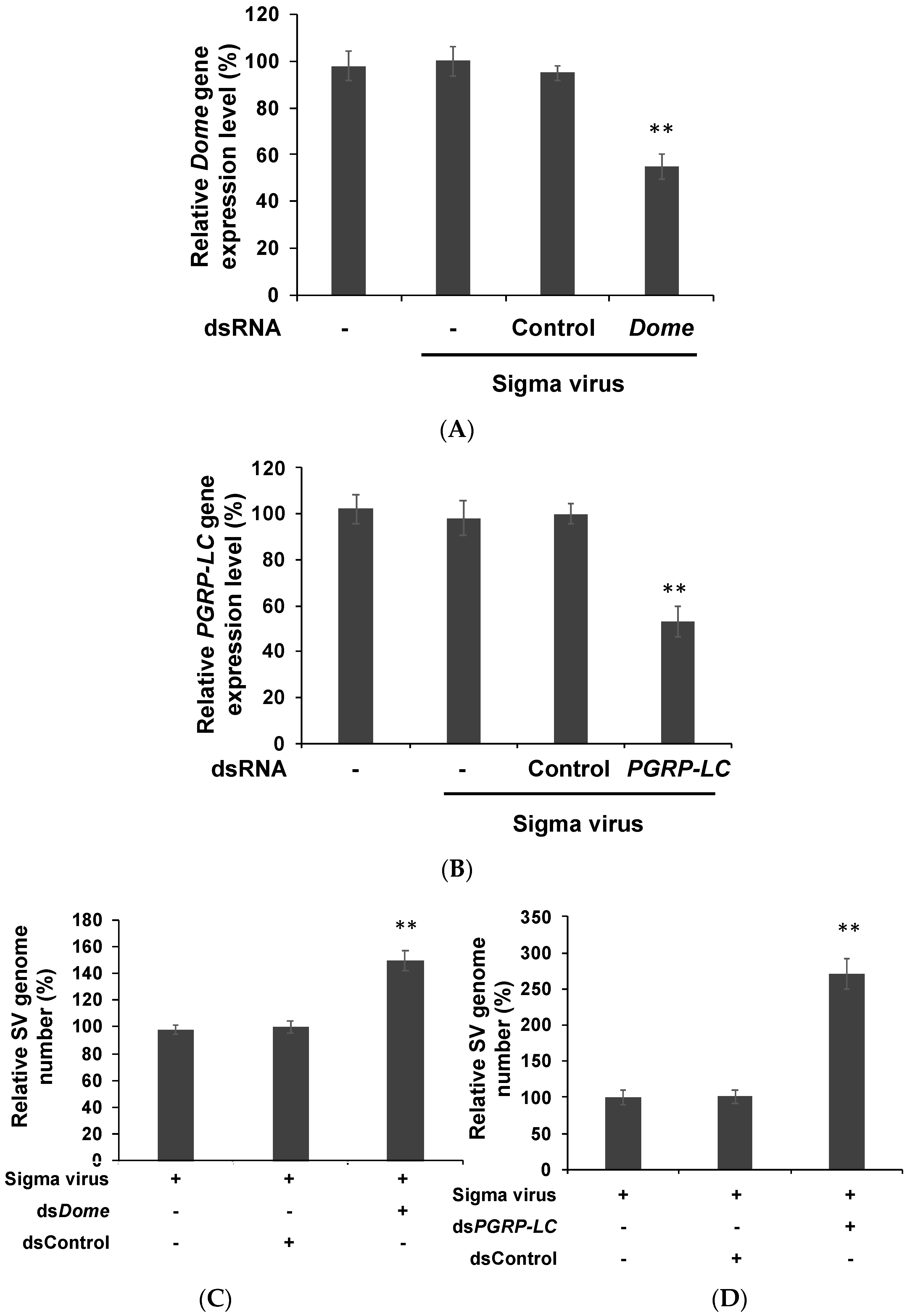
3.1. Viral Genome Replication Level Was Increased after the Knockdown of Immune Pathways in S2 Cells
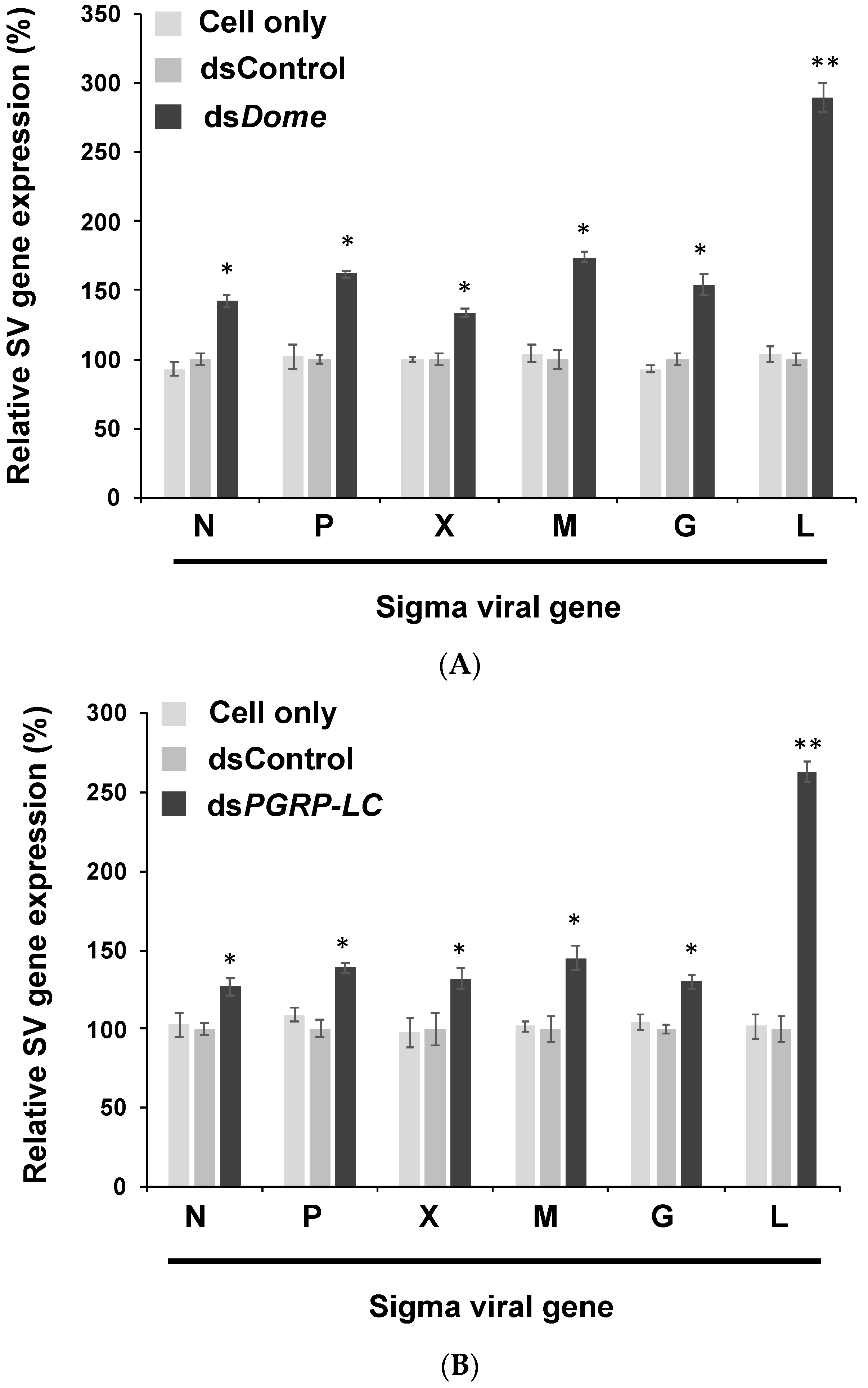
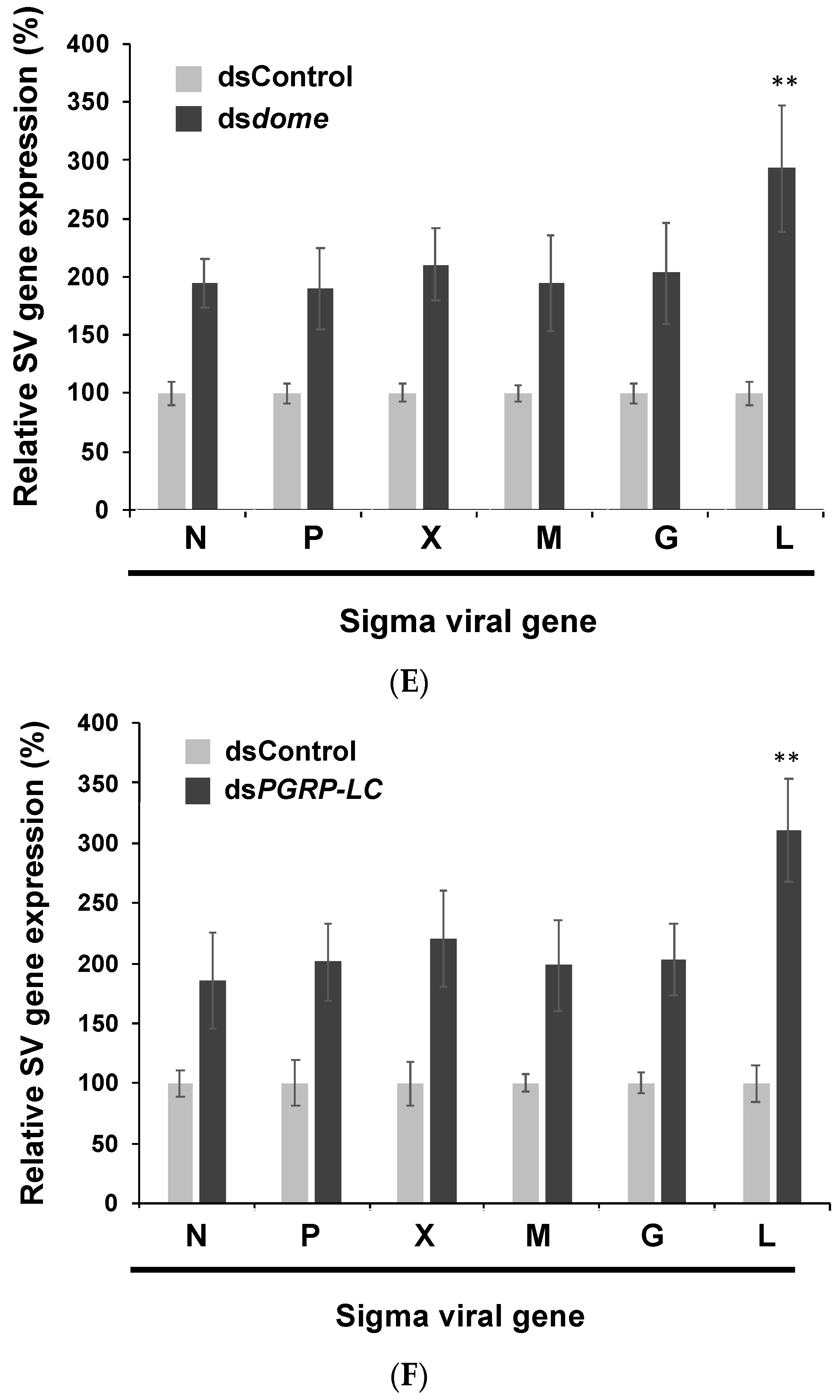
3.2. Viral Gene Expression Increased after the Knockdown of Immune Pathways in S2 Cells
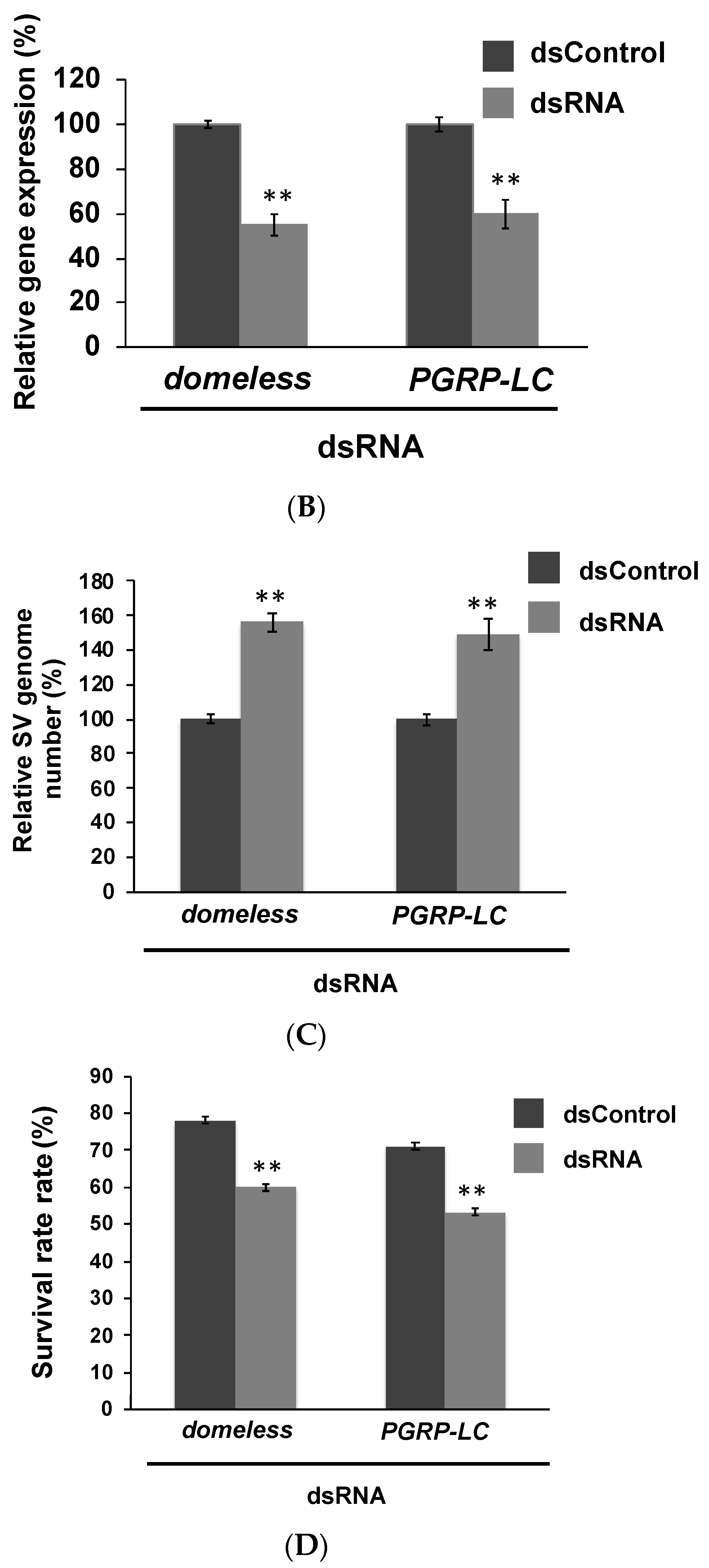
3.3. In Vivo Experiments Confirmed the Findings of in Vitro Experiments
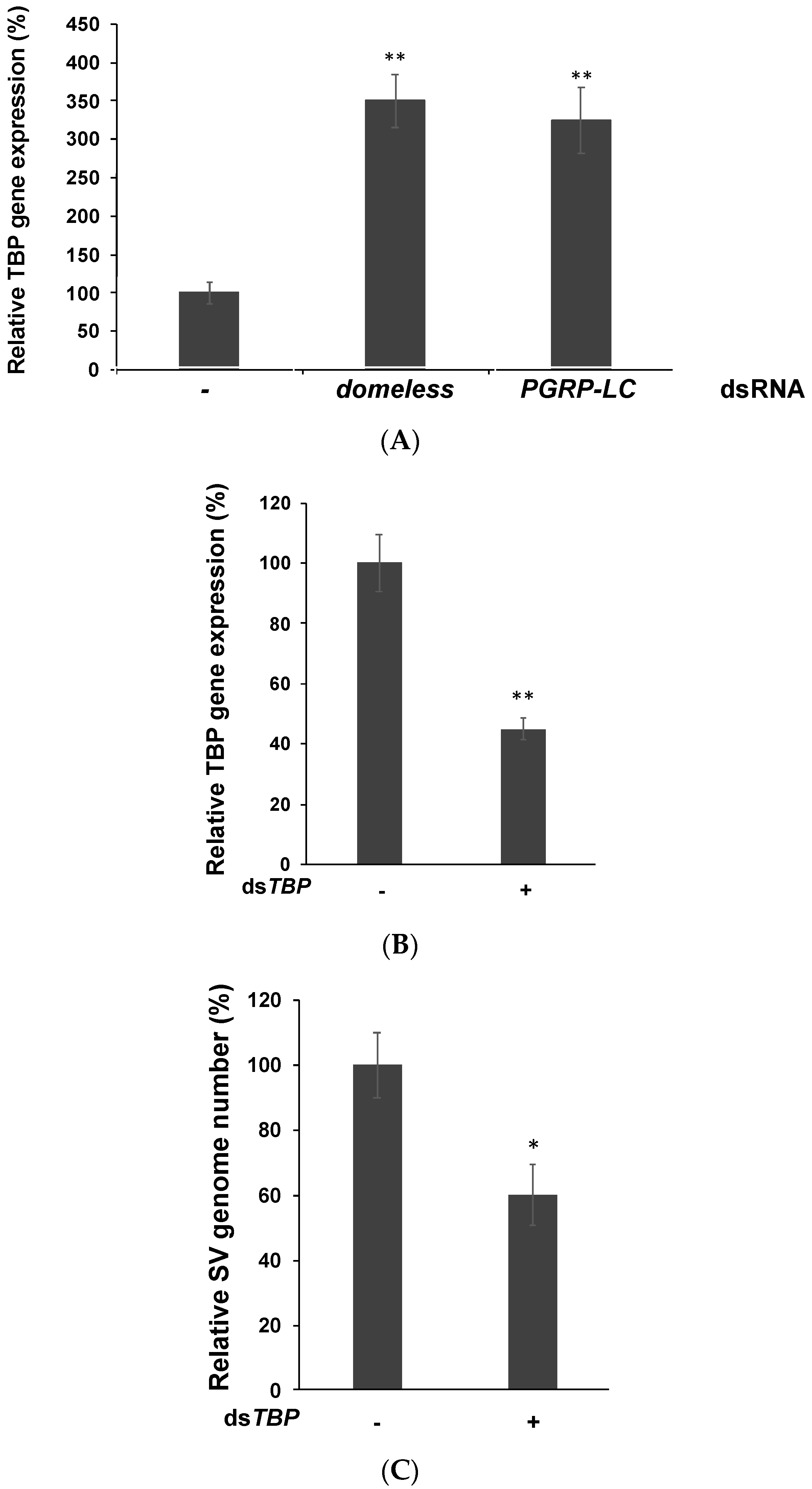
3.4. TATA-Binding Protein May Facilitate Viral Replication by Enhancing the RNA Polymerase Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chtarbanova, S.; Lamiable, O.; Lee, K.Z.; Galiana, D.; Troxler, L.; Meignin, C.; Hetru, C.; Hoffmann, J.A.; Daeffler, L.; Imler, J.L. Drosophila C Virus Systemic Infection Leads to Intestinal Obstruction. J. Virol. 2014, 88, 14057–14069. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.W.; McGraw, E.A.; Ammar, E.D.; Dietzgen, R.G.; Hogenhout, S.A. Drosophila melanogaster mounts a unique immune response to the Rhabdovirus sigma virus. Appl. Environ. Microbiol. 2008, 74, 3251–3256. [Google Scholar] [CrossRef] [PubMed]
- Longdon, B.; Obbard, D.J.; Jiggins, F.M. Sigma viruses from three species of Drosophila form a major new clade in the rhabdovirus phylogeny. Proc. R. Soc. B 2010, 277, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Li, Z.; Jenni, S.; Rahmeh, A.A.; Morin, B.M.; Grant, T.; Grigorieff, N.; Harrison, S.C.; Whelan, S.P.J. Structure of the L Protein of Vesicular Stomatitis Virus from Electron Cryomicroscopy. Cell 2015, 162, 314–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, T.J.; Luo, M. Structure of the vesicular stomatitis virus nucleocapsid in complex with the nucleocapsid-binding domain of the small polymerase cofactor, P. Proc. Natl. Acad. Sci. USA 2009, 106, 11713–11718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleuriet, A. Evolution of the proportions of two sigma viral types in experimental populations of Drosophila melanogaster in the absence of the allele that is restrictive of viral multiplication. Genetics 1999, 153, 1799–1808. [Google Scholar]
- Rosen, L. Carbon dioxide sensitivity in mosquitoes infected with sigma, vesicular stomatitis, and other rhabdoviruses. Science 1980, 207, 989–991. [Google Scholar] [CrossRef]
- Shroyer, D.A.; Rosen, L. Extrachromosomal inheritance of carbon dioxide sensitivity in the mosquito Culex quinquefasciatus. Genetics 1983, 104, 649–659. [Google Scholar]
- Carpenter, J.; Hutter, S.; Baines, J.F.; Roller, J.; Saminadin-Peter, S.S.; Parsch, J.; Jiggins, F.M. The transcriptional response of Drosophila melanogaster to infection with the sigma virus (Rhabdoviridae). PLoS ONE 2009, 4, e6838. [Google Scholar] [CrossRef]
- Fleuriet, A.; Sperlich, D. Evolution of the Drosophila melanogaster-sigma virus system in a natural population from Tubingen. Theor. Appl. Genet. 1992, 85, 186–189. [Google Scholar] [CrossRef]
- Kingsolver, M.B.; Huang, Z.; Hardy, R.W. Insect antiviral innate immunity: pathways, effectors, and connections. J. Mol. Biol. 2013, 425, 4921–4936. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Jouanguy, E.; Irving, P.; Troxler, L.; Galiana-Arnoux, D.; Hetru, C.; Hoffmann, J.A.; Imler, J.L. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of drosophila. Nat. Immunol. 2005, 6, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Goldman, W.E.; Mellroth, P.; Steiner, H.; Fukase, K.; Kusumoto, S.; Harley, W.; Fox, A.; Golenbock, D.; Silverman, N. Monomeric and polymeric gram-negative peptidoglycan but not purified LPS stimulate the Drosophila IMD pathway. Immunity 2004, 20, 637–649. [Google Scholar] [CrossRef]
- Mueller, S.; Gausson, V.; Vodovar, N.; Deddouche, S.; Troxler, L.; Perot, J.; Pfeffer, S.; Hoffmann, J.A.; Saleh, M.C.; Imler, J.L. RNAi-mediated immunity provides strong protection against the negative-strand RNA vesicular stomatitis virus in Drosophila. Proc. Natl. Acad. Sci. USA 2010, 107, 19390–19395. [Google Scholar] [CrossRef] [PubMed]
- Bronkhorst, A.W.; van Cleef, K.W.; Vodovar, N.; Ince, I.A.; Blanc, H.; Vlak, J.M.; Saleh, M.C.; van Rij, R.P. The DNA virus Invertebrate iridescent virus 6 is a target of the Drosophila RNAi machinery. Proc. Natl. Acad. Sci. USA 2012, 109, E3604–E3613. [Google Scholar] [CrossRef] [PubMed]
- Sabin, L.R.; Zheng, Q.; Thekkat, P.; Yang, J.; Hannon, G.J.; Gregory, B.D.; Tudor, M.; Cherry, S. Dicer-2 processes diverse viral RNA species. PLoS ONE 2013, 8, e55458. [Google Scholar] [CrossRef] [PubMed]
- Gaines, P.J.; Olson, K.E.; Higgs, S.; Powers, A.M.; Beaty, B.J.; Blair, C.D. Pathogen-derived resistance to dengue type 2 virus in mosquito cells by expression of the premembrane coding region of the viral genome. J. Virol. 1996, 70, 2132–2137. [Google Scholar] [Green Version]
- Zambon, R.A.; Nandakumar, M.; Vakharia, V.N.; Wu, L.P. The Toll pathway is important for an antiviral response in Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 7257–7262. [Google Scholar] [CrossRef]
- Zambon, R.A.; Vakharia, V.N.; Wu, L.P. RNAi is an antiviral immune response against a dsRNA virus in Drosophila melanogaster. Cell Microbiol. 2006, 8, 880–889. [Google Scholar] [CrossRef]
- Sabin, L.R.; Hanna, S.L.; Cherry, S. Innate antiviral immunity in Drosophila. Curr. Opin. Immunol. 2010, 22, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Aliyari, R.; Wu, Q.; Li, H.W.; Wang, X.H.; Li, F.; Green, L.D.; Han, C.S.; Li, W.X.; Ding, S.W. Mechanism of induction and suppression of antiviral immunity directed by virus-derived small RNAs in Drosophila. Cell Host Microbe 2008, 4, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Belvin, M.P.; Anderson, K.V. A conserved signaling pathway: the Drosophila toll-dorsal pathway. Annu. Rev. Cell Dev. Biol. 1996, 12, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Choe, K.M.; Lee, H.; Anderson, K.V. Drosophila peptidoglycan recognition protein LC (PGRP-LC) acts as a signal-transducing innate immune receptor. Proc. Natl. Acad. Sci. USA 2005, 102, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Luo, L. Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron 1999, 22, 451–461. [Google Scholar] [CrossRef]
- Teninges, D.; Bras-Herreng, F. Rhabdovirus sigma, the hereditary CO2 sensitivity agent of Drosophila: nucleotide sequence of a cDNA clone encoding the glycoprotein. J. Gen. Virol. 1987, 68 Pt 10, 2625–2638. [Google Scholar] [CrossRef]
- Longdon, B.; Hadfield, J.D.; Webster, C.L.; Obbard, D.J.; Jiggins, F.M. Host phylogeny determines viral persistence and replication in novel hosts. PLoS Pathogens 2011, 7, e1002260. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, Y.W.; Wu, C.P.; Wu, T.C.; Wu, Y.L. Analyses of the transcriptome of Bombyx mori cells infected with either BmNPV or AcMNPV. J. Asia-Pac. Entomol. 2018, 21, 37–45. [Google Scholar] [CrossRef]
- Dexter, F. Wilcoxon-Mann-Whitney Test Used for Data That Are Not Normally Distributed. Anesth. Analg. 2013, 117, 537–538. [Google Scholar] [CrossRef]
- Hu, Y.T.; Wu, T.C.; Yang, E.C.; Wu, P.C.; Lin, P.T.; Wu, Y.L. Regulation of genes related to immune signaling and detoxification in Apis mellifera by an inhibitor of histone deacetylation. Sci. Rep-Uk 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Merkling, S.H.; van Rij, R.P. Analysis of resistance and tolerance to virus infection in Drosophila. Nat. Protocols 2015, 10, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Quadt, I.; Mainz, D.; Mans, R.; Kremer, A.; Knebel-Morsdorf, D. Baculovirus infection raises the level of TATA-binding protein that colocalizes with viral DNA replication sites. J. Virol. 2002, 76, 11123–11127. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Yano, T.; Aggarwal, K.; Lim, J.H.; Ueda, K.; Oshima, Y.; Peach, C.; Erturk-Hasdemir, D.; Goldman, W.E.; Oh, B.H.; et al. PGRP-LC and PGRP-LE have essential yet distinct functions in the drosophila immune response to monomeric DAP-type peptidoglycan. Nat. Immunol. 2006, 7, 715–723. [Google Scholar] [CrossRef]
- Hsu, S.F.; Su, W.C.; Jeng, K.S.; Lai, M.M. A host susceptibility gene, DR1, facilitates influenza A virus replication by suppressing host innate immunity and enhancing viral RNA replication. J. Virol. 2015, 89, 3671–3682. [Google Scholar] [CrossRef]
- Wang, H.D.; Trivedi, A.; Johnson, D.L. Regulation of RNA polymerase I-dependent promoters by the hepatitis B virus X protein via activated Ras and TATA-binding protein. Mol. Cell. Biol. 1998, 18, 7086–7094. [Google Scholar] [CrossRef]
- Ward, D.M.; Vaughn, M.B.; Shiflett, S.L.; White, P.L.; Pollock, A.L.; Hill, J.; Schnegelberger, R.; Sundquist, W.I.; Kaplan, J. The role of LIP5 and CHMP5 in multivesicular body formation and HIV-1 budding in mammalian cells. J. Biol. Chem. 2005, 280, 10548–10555. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | dsRNA Synthesis Primers |
|---|---|
| domeless (dome) | F: TAATACGACTCACTATAGGG TAACGGCAAGAGCGC |
| R: TAATACGACTCACTATAGGG AGGTTCTGGCCAGGT | |
| PGRP-LC | F: TAATACGACTCACTATAGGGG GCGGTT TCCATACGG |
| R: TAATACGACTCACTATAGGGG CCATTGCTGACGCTC | |
| GFP | F: GCTCGGGAGATCTCCTGCCTTTGGGTGTGTCTGGG |
| R: CTAGACTCGAGCGGCCAACGGATCCTTCGTAGCCC | |
| TBP | F: AATTAACCCTCACTAAAGGGAT GGACCAAATGCTAAGCCC |
| R: AATTAACCCTCACTAAAGGGTACTTTCTCGCTGCCAGTCT |
| Gene | qPCR Check Primers | Gene | qPCR Check Primers |
|---|---|---|---|
| domeless (dome) | F: ACAACAGGCGTCTTCGGATT | SV-NP | F: TAACTCGGGTGTGACAGCTC |
| R: ACCCTTCAGTTTTGCCATGGT | R: CTTCGTTCATCTTCCTGGGT | ||
| PGRP-LC | F: CGCAAGGCCGTCACAGTTAC | SV-N | F: CACATGAGAAAATGCAAACAGCTT |
| R: GGTTCAACGTCTTTCCGAAGAG | R: GAAAATGGAGCGAGGATCGA | ||
| Diptericin (Dpt) | F: CTATTCATTGGACTGGCTTGTGCC | SV-P | F: TCAAACCCAGAGCCAGAGATAGTAT |
| R: TGGAACTGGCGACGCACTCT | R: CGCTTTTATCTGACGCTCAGGTA | ||
| TEPS | F: AACTCCGCAAACACCAAGTTGG | SV-X | F: TGGCCCCAATATTTCCTGAA |
| R: CTTCAACGCTTCGTGTAACACCAC | R: GCGTCACTCCATCAGGGTTT | ||
| Actin | F: CAAAGCGCAAAAAGAACACA | SV-M | F: ACACACTCCACAGTTTACCACCAT |
| R: AGAGGAGAGGGCGAGGTTAG | R: CGCCCTCCTGTCAATGAATAG | ||
| GFP | F: GTGTTCAATGCTTTGCGAGA | SV-G | F: CCATGTTTCGTTGAGCTTTCC |
| R: AAAGGGCAGATTGTGTGGAC | R: CGCCTTCGTGTTCACTGAGTT | ||
| TBP | F: TAGTGGCCAATCCTGTGTACCA | SV-L | F: TTCCCTGAAGACGCCCATTA |
| R: TCAGCGGAACCTGGTGTGGC | R: TGCCGCCCTCATCCAA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, J.-F.; Wu, C.-P.; Tang, C.-K.; Tsai, C.-W.; Rouhová, L.; Wu, Y.-L. Identification of Regulatory Host Genes Involved in Sigma Virus Replication Using RNAi Knockdown in Drosophila. Insects 2019, 10, 339. https://doi.org/10.3390/insects10100339
Liao J-F, Wu C-P, Tang C-K, Tsai C-W, Rouhová L, Wu Y-L. Identification of Regulatory Host Genes Involved in Sigma Virus Replication Using RNAi Knockdown in Drosophila. Insects. 2019; 10(10):339. https://doi.org/10.3390/insects10100339
Chicago/Turabian StyleLiao, Jen-Fu, Carol-P Wu, Cheng-Kang Tang, Chi-Wei Tsai, Lenka Rouhová, and Yueh-Lung Wu. 2019. "Identification of Regulatory Host Genes Involved in Sigma Virus Replication Using RNAi Knockdown in Drosophila" Insects 10, no. 10: 339. https://doi.org/10.3390/insects10100339
APA StyleLiao, J.-F., Wu, C.-P., Tang, C.-K., Tsai, C.-W., Rouhová, L., & Wu, Y.-L. (2019). Identification of Regulatory Host Genes Involved in Sigma Virus Replication Using RNAi Knockdown in Drosophila. Insects, 10(10), 339. https://doi.org/10.3390/insects10100339