Sub-Lethal Doses of Clothianidin Inhibit the Conditioning and Biosensory Abilities of the Western Honeybee Apis mellifera

Abstract
:1. Introduction
2. Materials and Methods
2.1. Honeybees
2.2. Treatments
2.3. Movement Assay
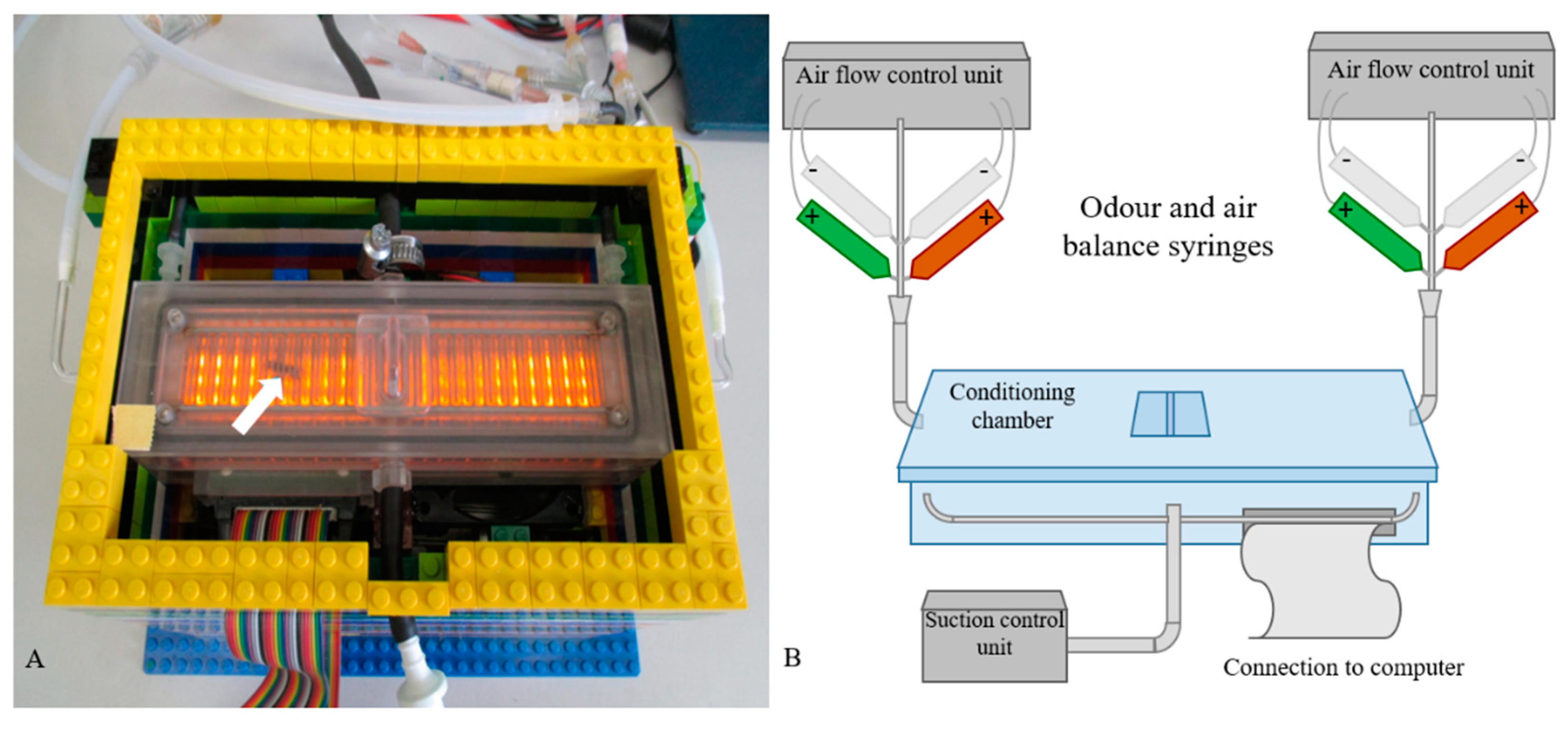
2.4. Conditioning
2.5. Statistical Analysis
3. Results
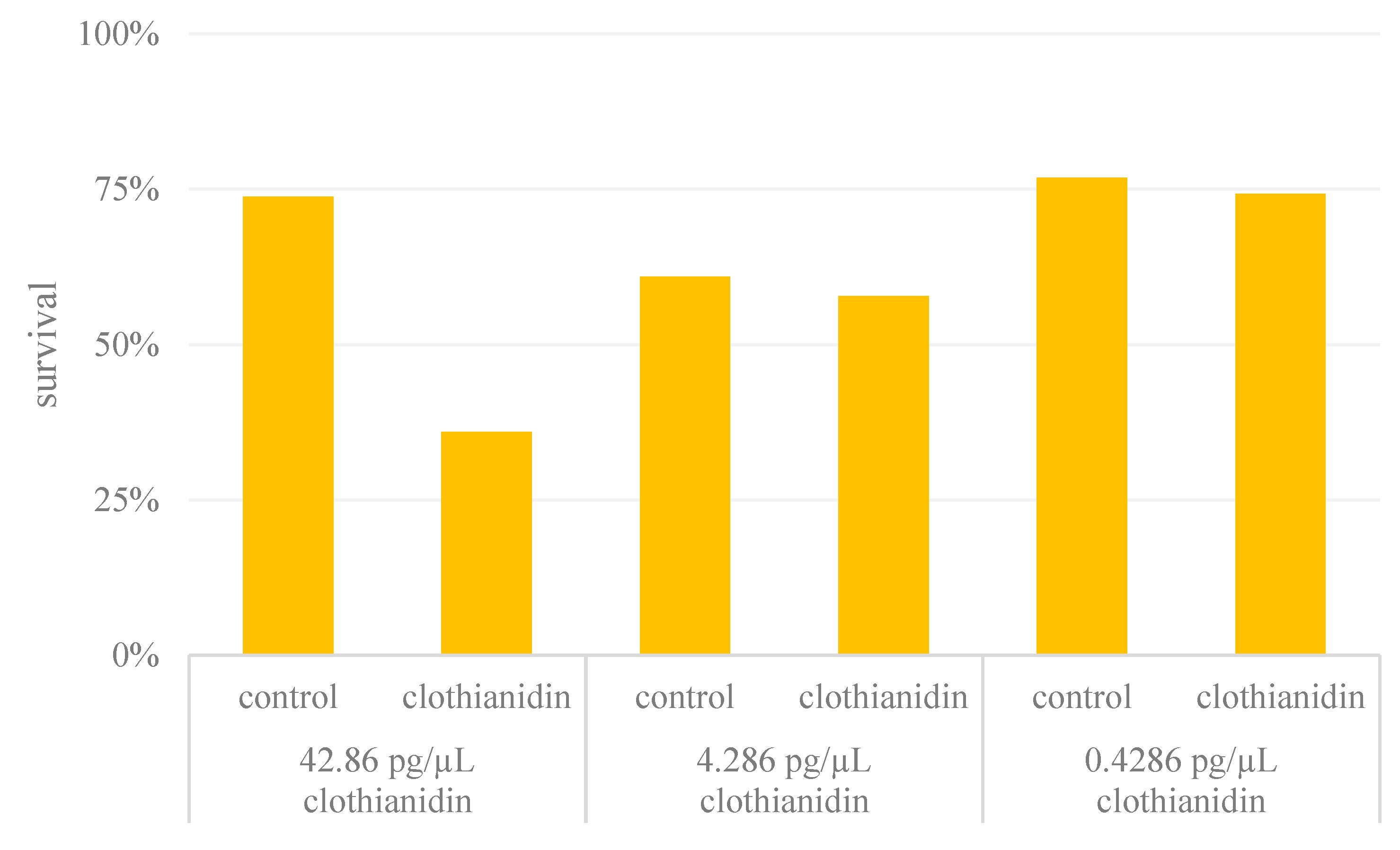
3.1. Mortality Rates in the Treatment and Control Groups
3.2. Movement Impairment in the Treatment and Control Groups
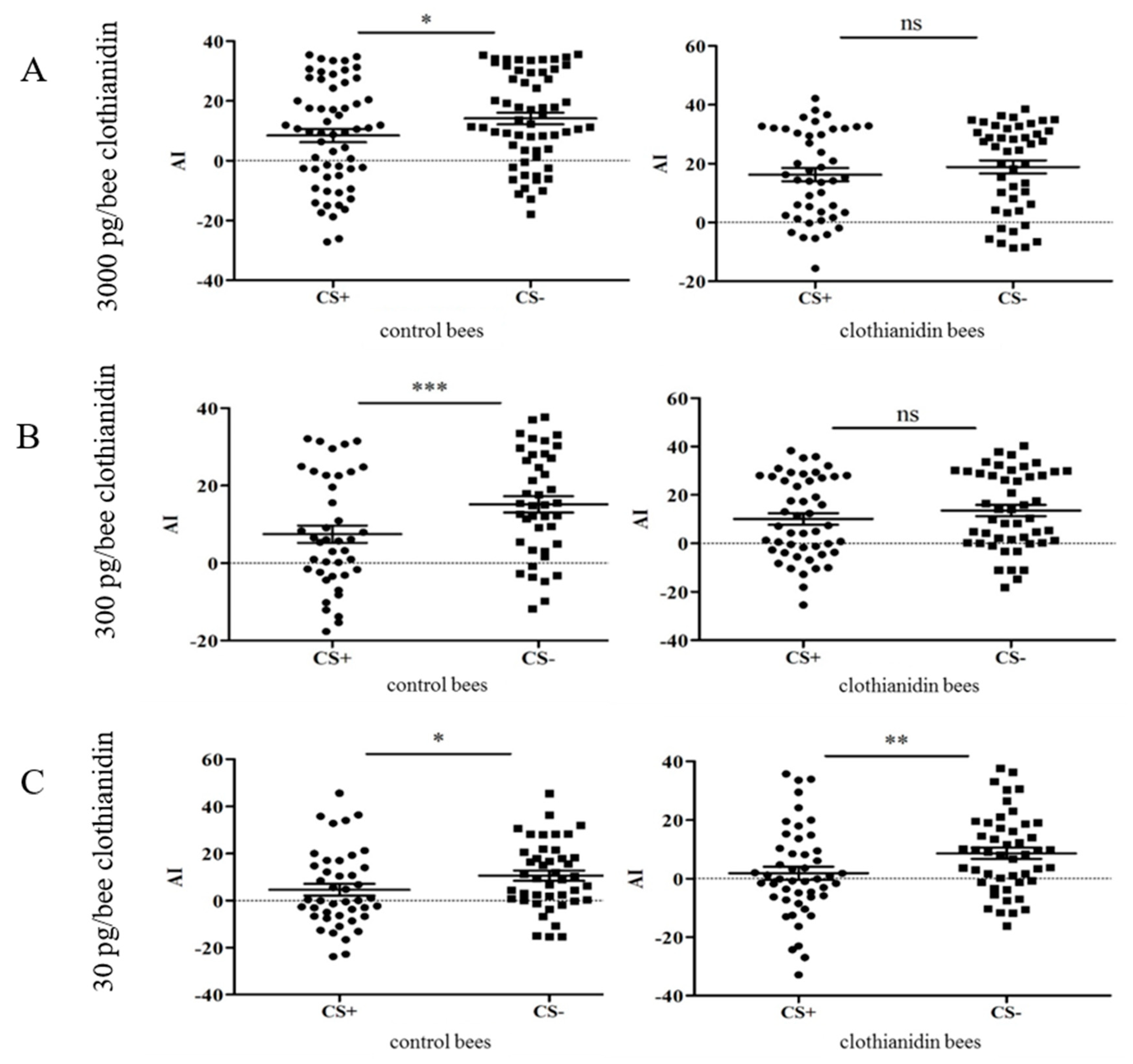
3.3. Conditioning and Biosensory Abilities in the Treatment and Control Groups
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.C. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Meissle, M.; Mouron, P.; Musa, T.; Bigler, F.; Pons, X.; Vasileiadis, V.P.; Otto, S.; Antichi, D.; Kiss, J.; Pálinkás, Z.; et al. Pests, pesticide use and alternative options in European maize production: Current status and future prospects. J. Appl. Entomol. 2010, 134, 357–375. [Google Scholar] [CrossRef]
- Alavanja, M.C. Introduction: Pesticides use and exposure extensive worldwide. Rev. Environ. Health 2009, 24, 303–309. [Google Scholar] [CrossRef]
- Tapparo, A.; Marton, D.; Giorio, C.; Zanella, A.; Soldà, L.; Marzaro, M.; Vivan, L.; Girolami, V. Assessment of the environmental exposure of honeybees to particulate matter containing neonicotinoid insecticides coming from corn coated seeds. Environ. Sci. Technol. 2012, 46, 2592–2599. [Google Scholar] [CrossRef]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. Int. 2015, 22, 35–67. [Google Scholar] [CrossRef]
- Brown, M.J.F.; Paxton, R.J. The conservation of bees: A global perspective. Apidologie 2009, 40, 410. [Google Scholar] [CrossRef]
- Genersch, E.; von der Ohe, W.; Kaatz, H.; Schroeder, A.; Otten, C.; Büchler, R.; Berg, S.; Ritter, W.; Mühlen, W.; Gisder, S.; et al. The German bee monitoring project: A long term studyto understand periodically high winter losses of honey beecolonies. Apidologie 2010, 41, 332–352. [Google Scholar] [CrossRef]
- Jacques, A.; Laurent, M.; EPILOBEE Consortium; Ribière-Chabert, M.; Saussac, M.; Bougeard, S.; Budge, G.E.; Hendrikx, P.; Chauzat, M.-P. A pan-European epidemiological study reveals honey bee colony survival depends on beekeeper education and disease control. PLoS ONE 2017, 12, e0172591. [Google Scholar] [CrossRef] [Green Version]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.-P.; van Engelsdorp, D. Drivers of colony losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Retsching, G.; Williams, G.R.; Odemer, R.; Boltin, J.; Di Poto, C.; Mehmann, M.M.; Retschnig, P.; Winiger, P.; Rosenkranz, P.; Neumann, P. Effects, but no interactions, of ubiquitous pesticideand parasite stressors on honey bee (Apis mellifera) lifespan and behaviour in a colony environment. Environ. Microbiol. 2015, 17, 4322–4331. [Google Scholar] [CrossRef] [PubMed]
- Rolke, D.; Fuchs, S.; Grünewald, B.; Gao, Z.; Blenau, W. Large-scale monitoring of effects of clothianidin-dressedoilseed rape seeds on pollinating insects in Northern Germany:effects on honey bees (Apis mellifera). Ecotoxicology 2016, 25, 1648–1665. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siede, R.; Faust, L.; Meixner, M.D.; Maus, C.; Grünewald, B.; Büchler, R. Performance of honey bee colonies under a long-lasting dietary exposure to sublethal concentrations of the neonicotinoid insecticide thiacloprid. Pest Manag. Sci. 2017, 73, 1334–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odemer, R.; Nilles, L.; Linder, N.; Rosenkranz, P. Sublethal effects of clothianidin and Nosema spp. on the longevity and foraging activity of free flying honey bees. Ecotoxicology 2018, 27, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Odemer, R.; Rosenkranz, P. Chronic exposure to a neonicotinoid pesticide and a 1synthetic pyrethroid in full-sized honey bee colonies. bioRxiv 2018. [Google Scholar] [CrossRef]
- Aliouane, Y.; El Hassani, A.K.; Gary, V.; Armengaud, C.; Lambin, M.; Gauthier, M. Subchronic exposure of honeybees to suble.thal doses of pesticides: Effects on behavior. Environ. Toxicol. Chem. 2009, 28, 113–122. [Google Scholar] [CrossRef]
- Decourtye, A.; Devillers, J.; Cluzeau, S.; Charreton, M.; Pham-Delèque, M.H. Effects of imidacloprid and deltamethrin on associative learning in honeybees under semi-field and laboratory conditions. Ecotoxicol. Environ. Saf. 2004, 57, 410–419. [Google Scholar] [CrossRef]
- Decourtye, A.; Devillers, J.; Genecque, E.; Le Menach, K.; Budzinski, H.; Cluzeau, S.; Pham-Delèque, M.H. Comparative sublethal toxicity of nine pesticides on olfactory learning performances of the honeybee Apis mellifera. Arch. Environ. Contam. Toxicol. 2005, 48, 242–250. [Google Scholar] [CrossRef]
- Menzel, R. Wie Pestizide (Neonicotinoide) die Navigation, die Tanz-Kommunikation und das Lernverhaltenvon Bienen verändern. In Rundgespräche der Kommission für Ökologie; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2014. [Google Scholar]
- Bortolotti, L.; Montanari, R.; Marcelino, J.; Medrzycki, P.; Maini, S.; Porrini, C. Effects of sub-lethal imidacloprid doses on the homing rate and foraging activity of honey bees. Bull. Insectology 2003, 56, 63–67. [Google Scholar]
- Suchail, S.; Guez, D.; Belzunces, L. Toxicity of imidacloprid and its metobolites in Apis mellifera. In Hazards of Pesticides to Bees; INRA: Paris, France, 2001. [Google Scholar]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A common pesticide decreases foraging success and survival in honey bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef] [PubMed]
- Decourtye, A.; Lacassie, E.; Pham-Delegue, M.H. Learning performances of honeybees (Apis mellifera L.) are differentially affected by imidacloprid according to the season. Pest Manag. Sci. 2003, 59, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Niu, C.-Y.; Lei, C.-L.; Cui, J.-J.; Desneux, N. Use of an innovative T-tube maze assay and the proboscis extension response assay to assess sublethal effects of GM products and pesticides on learning capacity of the honey bee Apis mellifera L. Ecotoxicology 2010, 19, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Romero, R.; Chaufaux, J.; Pham-Delègue, M.-H. Effects of Cry1Ab protoxin, deltamethrin and imidacloprid on the foraging activity and the learning performances of the honeybee Apis mellifera, a comparative approach. Apidologie 2005, 36, 601–611. [Google Scholar] [CrossRef]
- Pistorius, J.; Bischoff, G.; Heimbach, U.; Stähler, M. Bee poisoning incidents in Germany in spring 2008 caused by abrasion of active substance from treated seeds during sowing of maize. Hazards of pesticides to bees. In Proceedings of the 10th International Symposium of the ICP-Bee Protection Group, Bucharest, Romania, 8–10 October 2008. [Google Scholar]
- Page, R.E., Jr.; Peng, C.Y. Aging and development in social insects with emphasis on the honey bee, Apis mellifera L. Exp. Gerontol. 2001, 36, 695–711. [Google Scholar] [CrossRef]
- Rueppell, O.; Bachelier, C.; Fondrk, M.K.; Page, R.E. Regulation of life history determines lifespan of worker honey bees (Apis mellifera L.). Exp. Gerontol. 2007, 42, 1020–1032. [Google Scholar] [CrossRef]
- Fahrbach, S.E.; Robinson, G.E. Juvenile hormone, behavioral maturation, and brain structure in the honey bee. Dev. Neurosci. 1996, 18, 102–114. [Google Scholar] [CrossRef]
- Galizia, C.G. Neuroscience. Brainwashing, honeybee style. Science 2007, 317, 326–327. [Google Scholar] [CrossRef]
- Kirkerud, N.H.; Wehmann, H.N.; Galizia, C.G.; Gustav, D. APIS-a novel approach for conditioning honey bees. Front. Behav. Neurosci. 2013, 7, 29. [Google Scholar] [CrossRef]
- Kirkerud, N.H.; Schlegel, U.; Galizia, C.G. Aversive learning of colored lights in walking honeybees. Front. Behav. Neurosci. 2017, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Schott, M.; Bischoff, G.; Eichner, G.; Vilcinskas, A.; Büchler, R.; Meixner, M.D.; Brandt, A. Temporal dynamics of whole body residues of the neonicotinoid insecticide imidacloprid in live or dead honeybees. Sci. Rep. 2017, 7, 6288. [Google Scholar] [CrossRef] [PubMed]
- Schott, M.; Klein, B.; Vilcinskas, A. Detection of illicit drugs by trained honeybees (Apis mellifera). PLoS ONE 2015, 10, e0128528. [Google Scholar] [CrossRef] [PubMed]
- Wehmann, H.-N.; Gustav, D.; Kirkerud, N.H.; Galizia, C.G. The sound and the fury—Bees hiss when expecting danger. PLoS ONE 2015, 10, e0118708. [Google Scholar] [CrossRef]
- Trevan, J.W. The error of determination of toxicity. Proc. R. Soc. London. Ser. B Contain. Pap. A Biol. Character 1927, 101, 483–514. [Google Scholar] [CrossRef] [Green Version]
- European_Commission. Chemicals, Contaminants and Pesticides: Clothianidin, D.D.-F.S.P.a.d.c; Health & Consumer Protection Directorate-General: Brussels, Belgium, 2005. [Google Scholar]
- Schmuck, R.; Keppler, J. Clothianidin—Exotocixological profile and risk assessment. Pflanzenschutz Nachr. Bayer 2003, 56, 26–58. [Google Scholar]
- Capaldi, E.A.; Smith, A.D.; Osborne, J.L.; Fahrbach, S.E.; Farris, S.M.; Reynolds, D.R.; Edwards, A.S.; Martin, A.; Robinson, G.E.; Poppy, G.M.; et al. Ontogeny of orientation flight in the honeybee revealed by harmonic radar. Nature 2000, 403, 537–540. [Google Scholar] [CrossRef]
- Dechaume Moncharmont, F.X.; Decourtye, A.; Hennequet-Hantier, C.; Pons, O.; Pham-Delèque, M.H. Statistical analysis of honeybee survival after chronic exposure to insecticides. Environ. Toxicol. Chem. 2003, 22, 3088–3094. [Google Scholar] [CrossRef]
- Dickel, F.; Münch, D.; Amdam, G.V.; Mappes, J.; Freitak, D. Increased survival of honeybees in the laboratory after simultaneous exposure to low doses of pesticides and bacteria. PLoS ONE 2018, 13, e0191256. [Google Scholar] [CrossRef]
- Williams, G.R.; Alaux, C.; Costa, C.; Csáki, T.; Doublet, V.; Eisenhardt, D.; Fries, I.; Kuhn, R.; McMahon, D.; Mderzycki, P.; et al. Standard methods for maintaining adult Apis mellifera in cages under in vitro laboratory conditions. J. Apic. Res. 2013, 52, 1–36. [Google Scholar] [CrossRef]
- Perry, C.J.; Sovik, E.; Myerscough, M.R.; Barron, A.B. Rapid behavioral maturation accelerates failure of stressed honey bee colonies. Proc. Natl. Acad. Sci. USA 2015, 112, 3427–3432. [Google Scholar] [CrossRef] [Green Version]
- Carreck, N.L.; Ratnieks, F.L.W. The dose makes the poison: Have “field realistic”rates of exposure of bees to neonicotinoidinsecticides been overestimated in laboratorystudies? J. Apic. Res. 2015, 53, 607–614. [Google Scholar] [CrossRef]
- Sponsler, D.B.; Johnson, R.M. Mechanistic modeling of pesticide exposure: The missing keystone of honey bee toxicology. Environ. Toxicol. Chem. 2017, 36, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Heuvel, B. Neonicotinoids/Systemic Insecticides and Their Effects on Insects in the Environment: An Overview in Context of the Honeybee Deaths in Germany in 2008; Mellifera e.V.: Rosenfeld, Germany, 2008. [Google Scholar]
- Yang, E.C.; Chang, H.C.; Wu, W.Y.; Chen, Y.W. Impaired olfactory associative behavior of honeybee workers due to contamination of imidacloprid in the larval stage. PLoS ONE 2012, 7, e49472. [Google Scholar] [CrossRef] [PubMed]
- Kropf, J.; Kelber, C.; Bieringer, K.; Rössler, W. Olfactory subsystems in the honeybee: Sensory supply and sex specificity. Cell Tissue Res. 2014, 357, 583–595. [Google Scholar] [CrossRef]
- Brill, M.F.; Rosenbaum, T.; Reus, I.; Kleineidam, C.J.; Nawrot, M.P.; Rössler, W. Parallel processing via a dual olfactory pathway in the honeybee. J. Neurosci. 2013, 33, 2443–2456. [Google Scholar] [CrossRef]
- Eiri, D.M.; Nieh, J.C. A nicotinic acetylcholine receptor agonist affects honey bee sucrose responsiveness and decreases waggle dancing. J. Exp. Biol. 2012, 215, 2022–2029. [Google Scholar] [CrossRef] [Green Version]
- Sherman, G.; Visscher, P.K. Honeybee colonies achieve fitness through dancing. Nature 2002, 419, 920–922. [Google Scholar] [CrossRef]
- Blacquière, T.; van der Steen, J.J.M. Three years of banning neonicotinoidinsecticides based on sub-lethal effects: Can weexpect to see effects on bees? Pest Manag. Sci. 2017, 73, 1299–1304. [Google Scholar] [CrossRef]
- Carreck, N.C. A beekeeper’s perspective on the neonicotinoid ban? Pest Manag. Sci. 2017, 73, 1295–1298. [Google Scholar] [CrossRef]
- Marzaro, M.; Vivan, L.; Targa, A.; Mazzon, L.; Mori, N.; Greatti, M.; Toffolo, E.P.; di Bernardo, A.; Giorio, C.; Marton, D.; et al. Lethal aerial powdering of honey bees with neonicotinoids from fragments of maize seed coat. Bull. Insectology 2011, 64, 119–126. [Google Scholar]
- Botias, C.; David, A.; Horwood, J.; Abdul-Sada, A.; Nicholls, E.; Hill, E.; Goulson, D. Neonicotinoid residues in wildflowers, a potential route of chronic exposure for bees. Environ. Sci. Technol. 2015, 49, 12731–12740. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.J.; Goulson, D. The environmental risks of neonicotinoid pesticides: A review of the evidence post 2013. Environ. Sci. Pollut. Res. 2017, 4, 17285–17325. [Google Scholar] [CrossRef] [PubMed]
- Hederer, M. Zulassung von Clothianidin: Offener Brief von Manfred Hederer, Präsident des Deutschen Berufs-und Erwerbsimkerbundes. 2006. Available online: http://www.cbgnetwork.org/2533.html (accessed on 1 June 2018).
- Xu, T.; Dyer, D.G.; McConnell, L.L.; Bondarenko, S.; Allen, R.; Heinemann, O. Clothianidin in agricultural soils and uptake into corn pollen and canola nectar after multiyear seed treatment applications. Environ. Toxicol. Chem. 2016, 35, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Dively, G.P.; Kamel, A. Insecticide residues in pollen and nectar of a cucurbit crop and their potential exposure to pollinators. J. Agric. Food Chem. 2012, 60, 4449–4456. [Google Scholar] [CrossRef] [PubMed]
- Codling, G.; Al Naggar, Y.; Giesy, J.P.; Robertson, A.J. Concentrations of neonicotinoid insecticides in honey, pollen and honey bees (Apis mellifera L.) in central Saskatchewan, Canada. Chemosphere 2016, 144, 2321–2328. [Google Scholar] [CrossRef]
- Cutler, G.C.; Scott-Dupree, C.D. Exposure to Clothianidin Seed-Treated Canola Has No Long-Term Impact on Honey Bees. J. Econ. Entomol. 2007, 100, 765–772. [Google Scholar] [CrossRef] [Green Version]
- Reetz, J.E.; Zühlke, S.; Spiteller, M.; Wallner, K. Neonicotinoid insecticides translocated in guttated droplets of seed-treated maize and wheat: A threat to honeybees? Apidologie 2011, 42, 596–606. [Google Scholar] [CrossRef]
- Reetz, J.E. Freisetzung von Neonicotinoiden aus der Saatgutbeizung in Guttation von Kulturpflanzen und deren Auswirkungen auf Honigbienen Apis mellifera L. (Hymenoptera: Apidae, in Angewandte Entomologie. Ph.D. thesis, Landesanstalt für Bienenkunde, University Hohenheim, Stuttgart, Germany, 8 September 2015. [Google Scholar]
- Johansson, T.S.K.; Johansson, M.P. Providing Honeybees with Water. Bee World 1978, 59, 11–17. [Google Scholar] [CrossRef]
- Cutler, G.C.; Scott-Dupree, C.D.; Sultan, M.; McFarlane, A.D.; Brewer, L. A large-scale field study examining effects of exposure to clothianidin seed-treated canola on honey bee colony health, development, and overwintering success. PeerJ 2014, 2, e652. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | Amount of Clothianidin per Microliter Sugar Syrup | Amount of Clothianidin per Bee/70 µL |
|---|---|---|
| Control | 0 pg | 0 pg |
| High | 42.86 pg | 3000 pg |
| Medium | 4.286 pg | 300 pg |
| Low | 0.4286 pg | 30 pg |
| Control | 3000 pg/Bee Clothianidin | 300 pg/Bee Clothianidin | 30 pg/Bee Clothianidin | |
|---|---|---|---|---|
| fed bees | 80 | 128 | 83 | 66 |
| experimental bees | 59 | 45 | 41 | 48 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartling, M.T.; Vilcinskas, A.; Lee, K.-Z. Sub-Lethal Doses of Clothianidin Inhibit the Conditioning and Biosensory Abilities of the Western Honeybee Apis mellifera. Insects 2019, 10, 340. https://doi.org/10.3390/insects10100340
Bartling MT, Vilcinskas A, Lee K-Z. Sub-Lethal Doses of Clothianidin Inhibit the Conditioning and Biosensory Abilities of the Western Honeybee Apis mellifera. Insects. 2019; 10(10):340. https://doi.org/10.3390/insects10100340
Chicago/Turabian StyleBartling, Merle T., Andreas Vilcinskas, and Kwang-Zin Lee. 2019. "Sub-Lethal Doses of Clothianidin Inhibit the Conditioning and Biosensory Abilities of the Western Honeybee Apis mellifera" Insects 10, no. 10: 340. https://doi.org/10.3390/insects10100340
APA StyleBartling, M. T., Vilcinskas, A., & Lee, K.-Z. (2019). Sub-Lethal Doses of Clothianidin Inhibit the Conditioning and Biosensory Abilities of the Western Honeybee Apis mellifera. Insects, 10(10), 340. https://doi.org/10.3390/insects10100340