Soil Microarthropods and Soil Health: Intersection of Decomposition and Pest Suppression in Agroecosystems



{kind=link}
Abstract
:1. Introduction
2. Soil Health at the Intersection of Decomposition and Pest Suppression
3. Effects on Plant Nutrition, Nutrient Balance, and Soil and Plant Health
3.1. Plant Nutrition
3.2. Effects on Nutrient Balance
3.3. Enhanced Plant Tolerance or Resistance to Pests through Induced Resistance
3.4. Predation
3.5. Support of Natural Enemies of Pests as Alternate Prey
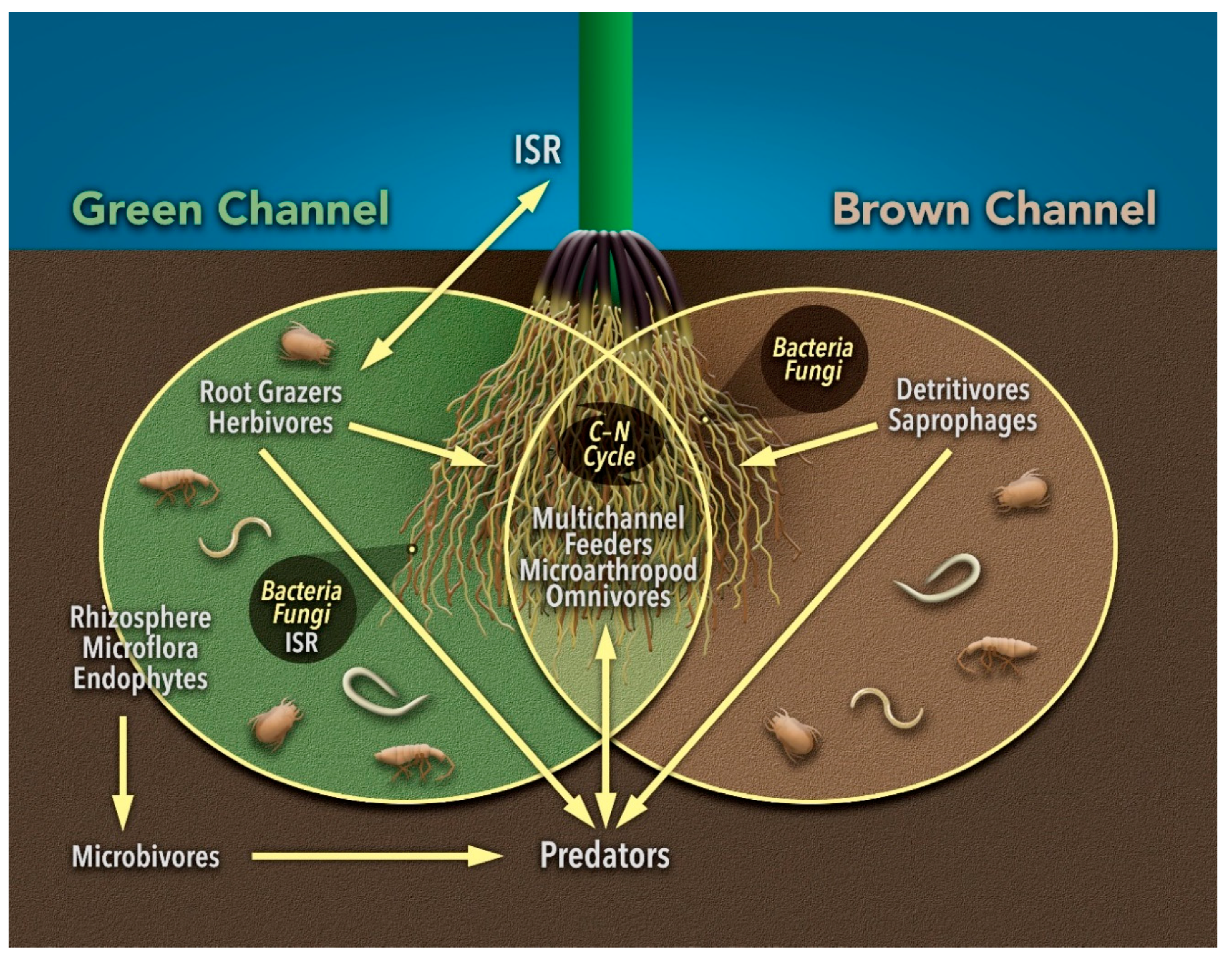
3.6. Multi-Channel Feeding
3.7. Climate Change
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol. Lett. 2013, 16, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Siddiky, M.R.K.; Schaller, J.; Caruso, T.; Rillig, M.C. Arbuscular mycorrhizal fungi and collembola non-additively increase soil aggregation. Soil Biol. Biochem. 2012, 47, 93–99. [Google Scholar] [CrossRef]
- Wolters, V. Invertebrate control of soil organic matter stability. Biol. Fert. Soils 2000, 31, 1–19. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Till. Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Briones, M.J.I. The serendipitous value of soil fauna in ecosystem functioning: The unexplained explained. Front. Environ. Sci. 2018, 6, 149. [Google Scholar] [CrossRef]
- NRCS. Soil Health. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/main/national/soils/health/ (accessed on 7 September 2019).
- Lavelle, P.; Decaens, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Culliney, T. Role of arthropods in maintaining soil fertility. Agriculture 2013, 3, 629–659. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef]
- Seastedt, T.R. The role of microarthropods in decomposition and mineralization processes. Annu. Rev. Entomol. 1984, 29, 25–46. [Google Scholar] [CrossRef]
- Neher, D.A. Soil community composition and ecosystem processes—Comparing agricultural ecosystems with natural ecosystems. Agroforest. Syst. 1999, 45, 159–185. [Google Scholar] [CrossRef]
- Coleman, D.C.; Callaham, M.A.J.; Crossley, D.A.J. Fundamentals of Soil Ecology, 3rd ed.; Academic Press: New York, NY, USA, 2018. [Google Scholar]
- Castro-Huerta, R.A.; Falco, L.B.; Sandler, R.V.; Coviella, C.E. Differential contribution of soil biota groups to plant litter decomposition as mediated by soil use. PeerJ 2015, 3, e826. [Google Scholar] [CrossRef] [PubMed]
- Kampichler, C.; Bruckner, A. The role of microarthropods in terrestrial decomposition: A meta-analysis of 40 years of litterbag studies. Biol. Rev. 2009, 84, 375–389. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Wall, D.H.; Six, J. Soil biodiversity and the environment. Annu. Rev. Env. Resour. 2015, 40, 63–90. [Google Scholar] [CrossRef]
- Brussaard, L.; Pulleman, M.M.; Ouedraogo, E.; Mando, A.; Six, J. Soil fauna and soil function in the fabric of the food web. Pedobiologia 2007, 50, 447–462. [Google Scholar] [CrossRef]
- Moore, J.C.; Walter, D.E.; Hunt, H.W. Arthropod regulation of micro- and mesobiota in below-ground detrital food webs. Annu. Rev. Entomol. 1988, 33, 419–439. [Google Scholar] [CrossRef]
- Feeny, P. Plant apparency and chemical defense. In Biochemical Interaction between Plants and Insects; Springer: New York, NY, USA, 1976; pp. 1–40. [Google Scholar]
- Root, R.B. Organization of a plant-arthropod association in simple and diverse habitats: The fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A. Aboveground-Belowground Linkages: Biotic Interactions, Ecosystem Processes; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Bezemer, T.M.; van Dam, N.M. Linking aboveground and belowground interactions via induced plant defenses. Trends Ecol. Evol. 2005, 20, 617–624. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Vet, L.E.M.; Harvey, J.A.; Wackers, F.L. Linking above- and belowground multitrophic interactions of plants, herbivores, pathogens, and their antagonists. Trends Ecol. Evol. 2001, 16, 547–554. [Google Scholar] [CrossRef]
- Erb, M.; Lenk, C.; Degenhardt, J.; Turlings, T.C. The underestimated role of roots in defense against leaf attackers. Trends Plant Sci. 2009, 14, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Moreau, G.; Eveleigh, E.S.; Lucarotti, C.J.; Quiring, D.T. Ecosystem alteration modifies the relative strengths of bottom-up and top-down forces in a herbivore population. J. Anim. Ecol. 2006, 75, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Morris, W.F.; Hufbauer, R.A.; Agrawal, A.A.; Bever, J.D.; Borowicz, V.A.; Gilbert, G.S.; Maron, J.L.; Mitchell, C.E.; Parker, I.M.; Power, A.G.; et al. Direct and interactive effects of enemies and mutualists on plant performance: A meta-analysis. Ecology 2007, 88, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Phelan, P.L. Soil-management history and the role of plant mineral balance as a determinant of maize susceptibility to the European corn borer. Biol. Agric. Hortic. 1997, 15, 25–34. [Google Scholar] [CrossRef]
- Bezemer, T.M.; Fountain, M.T.; Barea, J.M.; Christensen, S.; Dekker, S.C.; Duyts, H.; van Hal, R.; Harvey, J.A.; Hedlund, K.; Maraun, M.; et al. Divergent composition but similar function of soil food webs of individual plants: Plant species and community effects. Ecology 2010, 91, 3027–3036. [Google Scholar] [CrossRef] [PubMed]
- Ruf, A.; Kuzyakov, Y.; Lopatovskaya, O. Carbon fluxes in soil food webs of increasing complexity revealed by C-14 labelling and C-13 natural abundance. Soil Biol. Biochem. 2006, 38, 2390–2400. [Google Scholar] [CrossRef]
- Drigo, B.; Pijl, A.S.; Duyts, H.; Kielak, A.; Gamper, H.A.; Houtekamer, M.J.; Boschker, H.T.S.; Bodelier, P.L.E.; Whiteley, A.S.; van Veen, J.A.; et al. Shifting carbon flow from roots into associated microbial communities in response to elevated atmospheric CO2. Proc. Natl. Acad. Sci. USA 2010, 107, 10938–10942. [Google Scholar] [CrossRef]
- Ostle, N.; Briones, M.J.I.; Ineson, P.; Cole, L.; Staddon, P.; Sleep, D. Isotopic detection of recent photosynthate carbon flow into grassland rhizosphere fauna. Soil Biol. Biochem. 2007, 39, 768–777. [Google Scholar] [CrossRef]
- Bonkowski, M.; Clarholm, M. Stimulation of plant growth through interactions of bacteria and protozoa: Testing the auxiliary microbial loop hypothesis. Acta Protozool. 2012, 51, 237–247. [Google Scholar] [CrossRef]
- Chamberlain, P.M.; McNamara, N.P.; Chaplow, J.; Stott, A.W.; Black, H.I.J. Translocation of surface litter carbon into soil by Collembola. Soil Biol. Biochem. 2006, 38, 2655–2664. [Google Scholar] [CrossRef]
- Wickings, K.; Grandy, A.S. The oribatid mite Scheloribates moestus (Acari: Oribatida) alters litter chemistry and nutrient cycling during decomposition. Soil Biol. Biochem. 2011, 43, 351–358. [Google Scholar] [CrossRef]
- Kinnebrew, E.; Palawat, K.; Neher, D.A.; Galford, G.L. Detritivores’ contributions to carbon cycling: Implications for ecosystem servicese and agricultural benefits. Environ. Res. Lett. 2019. under review. [Google Scholar]
- Bradford, M.A.; Berg, B.; Maynard, D.S.; Wieder, W.R.; Wood, S.A. Understanding the dominant controls on litter decomposition. J. Ecol. 2016, 104, 229–238. [Google Scholar] [CrossRef]
- Dominati, E.; Patterson, M.; Mackay, A. A framework for classifying and quantifying the natural capital and ecosystem services of soils. Ecol. Econ. 2010, 69, 1858–1868. [Google Scholar] [CrossRef]
- Filser, J. The role of Collembola in carbon and nitrogen cycling in soil. Pedobiologia 2002, 46, 234–245. [Google Scholar] [CrossRef]
- Frouz, J. Effects of soil macro- and mesofauna on litter decomposition and soil organic matter stabilization. Geoderma 2018, 332, 161–172. [Google Scholar] [CrossRef]
- Kuťáková, E.; Cesarz, S.; Munzbergova, Z.; Eisenhauer, N. Soil microarthropods alter the outcome of plant-soil feedback experiments. Sci. Rep. 2018, 8, 11898. [Google Scholar] [CrossRef]
- Teuben, A.; Verhoef, H.A. Direct contribution by soil arthropods to nutrient availability through body and faecal nutrient content. Biol. Fert. Soils 1992, 14, 71–75. [Google Scholar] [CrossRef]
- Rodríguez, I.; Crespo, G.; Fraga, S.; Rodríguez, C.; Prieto, D. Activity of the mesofauna and the macrofauna in dung patches during their decomposition process. Cuban J. Agric. Sci. 2003, 37, 315–322. [Google Scholar]
- Beanland, L.; Phelan, P.L.; Salminen, S. Micronutrient interactions on soybean growth and the developmental performance of three insect herbivores. Environ. Entomol. 2003, 32, 641–651. [Google Scholar] [CrossRef]
- Busch, J.W.; Phelan, P.L. Mixture models of soybean growth and herbivore performance in response to nitrogen–sulphur–phosphorous nutrient interactions. Ecol. Entomol. 1999, 24, 132–145. [Google Scholar] [CrossRef]
- Chaboussou, F. Healthy Crops: A New Agricultural Revolution; Jon Carpenter Publishing: Oxford, UK, 2004. [Google Scholar]
- Waring, G.L.; Cobb, N.S. The impact of plant stress on herbivore population dynamics. In Insect-Plant Interactions; Bernays, E.A., Ed.; CRC Press: Boca Raton, FL, USA, 1992; Volume 4, pp. 167–226. [Google Scholar]
- Alyokhin, A.; Atlihan, R. Reduced fitness of the Colorado potato beetle (Coleoptera: Chrysomelidae) on potato plants grown in manure-amended soil. Environ. Entomol. 2005, 34, 963–968. [Google Scholar] [CrossRef] [Green Version]
- Boiteau, G.; Lynch, D.H.; Martin, R.C. Influence of fertilization on the Colorado potato beetle, Leptinotarsa decemlineata, in organic potato production. Environ. Entomol. 2008, 37, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Pangesti, N.; Pineda, A.; Pieterse, C.M.; Dicke, M.; van Loon, J.J. Two-way plant mediated interactions between root-associated microbes and insects: From ecology to mechanisms. Front. Plant Sci. 2013, 4, 414. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulou, G.V.; van Dam, N.M. Mechanisms and ecological implications of plant-mediated interactions between belowground and aboveground insect herbivores. Ecol. Res. 2017, 32, 13–26. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Pineda, A.; Zheng, S.J.; van Loon, J.J.; Pieterse, C.M.; Dicke, M. Helping plants to deal with insects: The role of beneficial soil-borne microbes. Trends Plant Sci. 2010, 15, 507–514. [Google Scholar] [CrossRef]
- Van Dam, N.M.; Heil, M. Multitrophic interactions below and above ground: En route to the next level. J. Ecol. 2011, 99, 77–88. [Google Scholar] [CrossRef]
- Briones, M.J. Soil fauna and soil functions: A jigsaw puzzle. Front. Environ. Sci. 2014, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Bukovinszky, T.; van Veen, F.J.F.; Jongema, Y.; Dicke, M. Direct and indirect effects of resource quality on food web structure. Science 2008, 319, 804–807. [Google Scholar] [CrossRef]
- Kaplan, I.; Halitschke, R.; Kessler, A.; Rehill, B.J.; Sardanelli, S.; Denno, R.F. Physiological integration of roots and shoots in plant defense strategies links above- and belowground herbivory. Ecol. Lett. 2008, 11, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Soler, R.; Van der Putten, W.H.; Harvey, J.A.; Vet, L.E.; Dicke, M.; Bezemer, T.M. Root herbivore effects on aboveground multitrophic interactions: Patterns, processes and mechanisms. J. Chem. Ecol. 2012, 38, 755–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurst, S. Plant-mediated links between detritivores and aboveground herbivores. Front. Plant Sci. 2013, 4, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Annu. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuman, M.C.; Baldwin, I.T. The layers of plant responses to insect herbivores. Annu. Rev. Entomol. 2016, 61, 373–394. [Google Scholar] [CrossRef]
- Balbi, V.; Devoto, A. Jasmonate signalling network in Arabidopsis thaliana: Crucial regulatory nodes and new physiological scenarios. New Phytol. 2008, 177, 301–318. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Mescher, M.C.; Tumlinson, J.H. Caterpillar-induced nocturnal plant volatiles repel conspecific females. Nature 2001, 410, 577–580. [Google Scholar] [CrossRef]
- Turlings, T.C.; Tumlinson, J.H.; Lewis, W.J. Exploitation of herbivore-induced plant odors by host-seeking parasitic wasps. Science 1990, 250, 1251–1253. [Google Scholar] [CrossRef] [Green Version]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef]
- Hiltpold, I.; Erb, M.; Robert, C.A.M.; Turlings, T.C.J. Systemic root signalling in a belowground, volatile-mediated tritrophic interaction. Plant Cell Environ. 2011, 34, 1267–1275. [Google Scholar] [CrossRef] [Green Version]
- Rasmann, S.; Turlings, T.C.J. First insights into specificity of belowground tritrophic interactions. Oikos 2008, 117, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [PubMed]
- Rodriguez, P.A.; Rothballer, M.; Chowdhury, S.P.; Nussbaumer, T.; Gutjahr, C.; Falter-Braun, P. Systems biology of plant-microbiome interactions. Mol. Plant 2019, 12, 804–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef]
- Ourry, M.; Lebreton, L.; Chaminade, V.; Guillerm-Erckelboudt, A.Y.; Herve, M.; Linglin, J.; Marnet, N.; Ourry, A.; Paty, C.; Poinsot, D.; et al. Influence of belowground herbivory on the dynamics of root and rhizosphere microbial communities. Front. Ecol. Evol. 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Endlweber, K.; Ruess, L.; Scheu, S. Collembola switch diet in presence of plant roots thereby functioning as herbivores. Soil Biol. Biochem. 2009, 41, 1151–1154. [Google Scholar] [CrossRef]
- Janoušková, M.; Kohout, P.; Moradi, J.; Doubková, P.; Frouz, J.; Vosolsobě, S.; Rydlová, J. Microarthropods influence the composition of rhizospheric fungal communities by stimulating specific taxa. Soil Biol. Biochem. 2018, 122, 120–130. [Google Scholar] [CrossRef]
- Ngosong, C.; Gabriel, E.; Ruess, L. Collembola grazing on arbuscular mycorrhiza fungi modulates nutrient allocation in plants. Pedobiologia 2014, 57, 171–179. [Google Scholar] [CrossRef]
- Hedlund, K.; Öhrn, M.S. Tritrophic interactions in a soil community enhance decomposition rates. Oikos 2000, 88, 585–591. [Google Scholar] [CrossRef]
- Maraun, M.; Martens, H.; Migge, S.; Theenhaus, A.; Scheu, S. Adding to ‘the enigma of soil animal diversity’: Fungal feeders and saprophagous soil invertebrates prefer similar food substrates. Eur. J. Soil Biol. 2003, 39, 85–95. [Google Scholar] [CrossRef]
- Losey, J.E.; Vaughan, M. The economic value of ecological services provided by insects. Bioscience 2006, 56, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, S.E.; Ellsworth, P.C.; Frisvold, G.B. Economic value of biological control in integrated pest management of managed plant systems. Annu. Rev. Entomol. 2015, 60, 621–645. [Google Scholar] [CrossRef] [Green Version]
- Berg, M.P.; Stoffer, M.; van den Heuvel, H.H. Feeding guilds in Collembola based on digestive enzymes. Pedobiologia 2004, 48, 589–601. [Google Scholar] [CrossRef]
- Köehler, H.H. Mesostigmata (Gamasina, Uropodina), efficient predators in agroecosystems. Agric. Ecol. Ecosyst. 1997, 62, 105–117. [Google Scholar] [CrossRef]
- Read, D.S.; Sheppard, S.K.; Bruford, M.W.; Glen, D.M.; Symondson, W.O.C. Molecular detection of predation by soil micro-arthropods on nematodes. Mol. Ecol. 2006, 15, 1963–1972. [Google Scholar] [CrossRef]
- Brust, G.E. Effects of below-ground predator–weed interactions on damage to peanut by southern corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 1990, 19, 1837–1844. [Google Scholar] [CrossRef]
- Welbourne, W.C. Potential use of trombidioid and erythraeoid mites as biological control agents of insect pests. In Biological Control of Pests by Mites; Hoy, M.A., Cunningham, G.L., Knutsen, L., Eds.; Special Publication 3304; Agricultural Experiment Station, Division of Agriculture and Natural Resources, University of California: Berkeley, CA, USA, 1983; pp. 103–140. [Google Scholar]
- Brust, G.E.; House, G.J. A study of Tyrophagus putrescentiae (Acari: Acaridae) as a facultative predator of southern corn rootworm eggs. Exp. Appl. Acarol. 1988, 4, 335–344. [Google Scholar] [CrossRef]
- Heidemann, K.; Scheu, S.; Ruess, L.; Maraun, M. Molecular detection of nematode predation and scavenging in oribatid mites: Laboratory and field experiments. Soil Biol. Biochem. 2011, 43, 2229–2236. [Google Scholar] [CrossRef]
- Kaneda, S.; Kaneko, N. Collembolans feeding on soil affect carbon and nitrogen mineralization by their influence on microbial and nematode activities. Biol. Fert. Soils 2008, 44, 435–442. [Google Scholar] [CrossRef]
- Sayre, R.M.; Walter, D.E. Factors affecting the efficacy of natural enemies of nematodes. Annu. Rev. Phytopathol. 1991, 29, 149–166. [Google Scholar] [CrossRef]
- Schneider, K.; Migge, S.; Norton, R.A.; Scheu, S.; Langel, R.; Reineking, A.; Maraun, M. Trophic niche differentiation in soil microarthropods (Oribatida, Acari): Evidence from stable isotope ratios (15N/14N). Soil Biol. Biochem. 2004, 36, 1769–1774. [Google Scholar] [CrossRef]
- Doncaster, C.C.; Murphy, P.W. A culture method for soil meiofauna and its application to the study of nematode predators 1. Nematologica 1957, 2, 202–214. [Google Scholar] [CrossRef]
- Chen, B.; Snider, R.J.; Snider, R.M. Food preference and effects of food type on the life history of some soil Collembola. Pedobiologia 1995, 39, 496–505. [Google Scholar]
- Polis, G.A.; Strong, D.R. Food web complexity and community dynamics. Am. Nat. 1996, 147, 813–846. [Google Scholar] [CrossRef]
- Chen, B.R.; Wise, D.H. Bottom-up limitation of predaceous arthropods in a detritus-based terrestrial food web. Ecology 1999, 80, 761–772. [Google Scholar] [CrossRef]
- Rypstra, A.L.; Marshall, S.D. Augmentation of soil detritus affects the spider community and herbivory in a soybean agroecosystem. Entomol. Exp. Appl. 2005, 116, 149–157. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Wright, D.J.; Leather, S.R. The effects of farming system and fertilisers on pests and natural enemies: A synthesis of current research. Agric. Ecol. Ecosyst. 2011, 141, 261–270. [Google Scholar] [CrossRef]
- Wise, D.H.; Snyder, W.E.; Tuntibunpakul, P.; Halaj, J. Spiders in decomposition food webs of agroecosystems: Theory and evidence. J. Arachnol. 1999, 27, 363–370. [Google Scholar]
- Muñoz-Cárdenas, K.; Ersin, F.; Pijnakker, J.; van Houten, Y.; Hoogerbrugge, H.; Leman, A.; Pappas, M.L.; Duarte, M.V.A.; Messelink, G.J.; Sabelis, M.W. Supplying high-quality alternative prey in the litter increases control of an above-ground plant pest by a generalist predator. Biol. Control 2017, 105, 19–26. [Google Scholar] [CrossRef]
- Bilde, T.; Axelsen, J.A.; Toft, S. The value of Collembola from agricultural soils as food for a generalist predator. J. Appl. Ecol. 2000, 37, 672–683. [Google Scholar] [CrossRef]
- Halaj, J.; Wise, D.H. Impact of a detrital subsidy on trophic cascades in a terrestrial grazing food web. Ecology 2002, 83, 3141–3151. [Google Scholar] [CrossRef]
- McNabb, D.M.; Halaj, J.; Wise, D.H. Inferring trophic positions of generalist predators and their linkage to the detrital food web in agroecosystems: A stable isotope analysis. Pedobiologia 2001, 45, 289–297. [Google Scholar] [CrossRef]
- Koss, A.M.; Snyder, W.E. Alternative prey disrupt biocontrol by a guild of generalist predators. Biol. Control 2005, 32, 243–251. [Google Scholar] [CrossRef]
- Symondson, W.O.; Cesarini, S.; Dodd, P.W.; Harper, G.L.; Bruford, M.W.; Glen, D.M.; Wiltshire, C.W.; Harwood, J.D. Biodiversity vs. biocontrol: Positive and negative effects of alternative prey on control of slugs by carabid beetles. Bull. Entomol. Res. 2006, 96, 637–645. [Google Scholar] [CrossRef]
- Birkhofer, K.; Wise, D.H.; Scheu, S. Subsidy from the detrital food web, but not microhabitat complexity, affects the role of generalist predators in an aboveground herbivore food web. Oikos 2008, 117, 494–500. [Google Scholar] [CrossRef]
- Scheunemann, N.; Digel, C.; Scheu, S.; Butenschoen, O. Roots rather than shoot residues drive soil arthropod communities of arable fields. Oecologia 2015, 179, 1135–1145. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Allesina, S.; Cottingham, K.L.; Moore, J.C.; Sandin, S.A.; de Mazancourt, C. Linking the green and brown worlds: The prevalence and effect of multichannel feeding in food webs. Ecology 2014, 95, 3376–3386. [Google Scholar] [CrossRef]
- Wolkovich, E.M. Reticulated channels in soil food webs. Soil Biol. Biochem. 2016, 102, 18–21. [Google Scholar] [CrossRef]
- Zou, K.; Thébault, E.; Lacroix, G.; Barot, S. Interactions between the green and brown food web determine ecosystem functioning. Funct. Ecol. 2016, 30, 1454–1465. [Google Scholar] [CrossRef] [Green Version]
- Bardgett, R.D. The Biology of Soil: A Community and Ecosystem Approach; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Digel, C.; Curtsdotter, A.; Riede, J.; Klarner, B.; Brose, U. Unravelling the complex structure of forest soil food webs: Higher omnivory and more trophic levels. Oikos 2014, 123, 1157–1172. [Google Scholar] [CrossRef]
- Glavatska, O.; Muller, K.; Butenschoen, O.; Schmalwasser, A.; Kandeler, E.; Scheu, S.; Totsche, K.U.; Ruess, L. Disentangling the root- and detritus-based food chain in the micro-food web of an arable soil by plant removal. PLoS ONE 2017, 12, e0180264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chahartaghi, M.; Langel, R.; Scheu, S.; Ruess, L. Feeding guilds in Collembola based on nitrogen stable isotope ratios. Soil Biol. Biochem. 2005, 37, 1718–1725. [Google Scholar] [CrossRef]
- Klarner, B.; Maraun, M.; Scheu, S. Trophic diversity and niche partitioning in a species rich predator guild–Natural variations in stable isotope ratios (13C/12C, 15N/14N) of mesostigmatid mites (Acari, Mesostigmata) from Central European beech forests. Soil Biol. Biochem. 2013, 57, 327–333. [Google Scholar] [CrossRef]
- Schneider, K.C.R.; Scheu, S.; Maraun, M. Feeding biology of oribatid mites: A minireview. Phytophaga 2004, 14, 247–256. [Google Scholar]
- Albers, D.; Schaefer, M.; Scheu, S. Incorporation of plant carbon into the soil animal food web of an arable system. Ecology 2006, 87, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Walter, D.E. Trophic behavior of mycophagous microarthropods. Ecology 1987, 68, 226–229. [Google Scholar] [CrossRef]
- Lee, Q.; Widden, P. Folsomia candida, a “fungivorous” collembolan, feeds preferentially on nematodes rather than soil fungi. Soil Biol. Biochem. 1996, 28, 689–690. [Google Scholar] [CrossRef]
- Moore, J.C.; Berlow, E.L.; Coleman, D.C.; de Ruiter, P.C.; Dong, Q.; Hastings, A.; Johnson, N.C.; McCann, K.S.; Melville, K.; Morin, P.J. Detritus, trophic dynamics and biodiversity. Ecol. Lett. 2004, 7, 584–600. [Google Scholar] [CrossRef]
- Rooney, N.; McCann, K.S. Integrating food web diversity, structure and stability. Trends Ecol. Evol. 2012, 27, 40–46. [Google Scholar] [CrossRef]
- McKenzie, D.C. Visual soil examination techniques as part of a soil appraisal framework for farm evaluation in Australia. Soil Till. Res. 2013, 127, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Van der Putten, W.H.; Macel, M.; Visser, M.E. Predicting species distribution and abundance responses to climate change: Why it is essential to include biotic interactions across trophic levels. Philos. Trans. R. Soc. B-Biol. Sci. 2010, 365, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- Dungait, J.A.J.; Hopkins, D.W.; Gregory, A.S.; Whitmore, A.P. Soil organic matter turnover is governed by accessibility not recalcitrance. Glob. Chang. Biol. 2012, 18, 1781–1796. [Google Scholar] [CrossRef]
- Ekelund, F.; Saj, S.; Vestergård, M.; Bertaux, J.; Mikola, J. The “soil microbial loop” is not always needed to explain protozoan stimulation of plants. Soil Biol. Biochem. 2009, 41, 2336–2342. [Google Scholar] [CrossRef]
- Osler, G.H.; Sommerkorn, M. Toward a complete soil C and N cycle: Incorporating the soil fauna. Ecology 2007, 88, 1611–1621. [Google Scholar] [CrossRef]
- ICPP. Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation: Special Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V., Stocker, T.F., Dahe, Q., Eds.; Cambridge University Press: Cambridge, UK, 2012; p. 582. [Google Scholar]
- Aerts, R. The freezer defrosting: Global warming and litter decomposition rates in cold biomes. J. Ecol. 2006, 94, 713–724. [Google Scholar] [CrossRef]
- Taylor, A.R.; Schroter, D.; Pflug, A.; Wolters, V. Response of different decomposer communities to the manipulation of moisture availability: Potential effects of changing precipitation patterns. Glob. Chang. Biol. 2004, 10, 1313–1324. [Google Scholar] [CrossRef]
- Coûteaux, M.M.; Bolger, T. Interactions between atmospheric CO2 enrichment and soil fauna. Plant Soil 2000, 224, 123–134. [Google Scholar] [CrossRef]
- Darby, B.J.; Neher, D.A.; Housman, D.C.; Belnap, J. Few apparent short-term effects of elevated soil temperature and increased frequency of summer precipitation on the abundance and taxonomic diversity of desert soil micro-and meso-fauna. Soil Biol. Biochem. 2011, 43, 1474–1481. [Google Scholar] [CrossRef]
- Ferguson, S.H.; Joly, D.O. Dynamics of springtail and mite populations: The role of density dependence, predation, and weather. Ecol. Entomol. 2002, 27, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Sjursen, H.; Michelsen, A.; Jonasson, S. Effects of long-term soil warming and fertilisation on microarthropod abundances in three sub-arctic ecosystems. Appl. Soil Ecol. 2005, 30, 148–161. [Google Scholar] [CrossRef]
- Lensing, J.R.; Todd, S.; Wise, D.H. The impact of altered precipitation on spatial stratification and activity-densities of springtails (Collembola) and spiders (Araneae). Ecol. Entomol. 2005, 30, 194–200. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A., Jr. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Lavelle, P. Ecological challenges for soil science. Soil Sci. 2000, 165, 73–86. [Google Scholar] [CrossRef]
- Wall, D.H.; Bradford, M.A.; St John, M.G.; Trofymow, J.A.; Behan-Pelletier, V.; Bignell, D.D.E.; Dangerfield, J.M.; Parton, W.J.; Rusek, J.; Voigt, W.; et al. Global decomposition experiment shows soil animal impacts on decomposition are climate-dependent. Glob. Chang. Biol. 2008, 14, 2661–2677. [Google Scholar] [CrossRef] [Green Version]
- Grandy, A.S.; Wieder, W.R.; Wickings, K.; Kyker-Snowman, E. Beyond microbes: Are fauna the next frontier in soil biogeochemical models? Soil Biol. Biochem. 2016, 102, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Soong, J.L.; Nielsen, U.N. The role of microarthropods in emerging models of soil organic matter. Soil Biol. Biochem. 2016, 102, 37–39. [Google Scholar] [CrossRef] [Green Version]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neher, D.A.; Barbercheck, M.E. Soil Microarthropods and Soil Health: Intersection of Decomposition and Pest Suppression in Agroecosystems. Insects 2019, 10, 414. https://doi.org/10.3390/insects10120414
Neher DA, Barbercheck ME. Soil Microarthropods and Soil Health: Intersection of Decomposition and Pest Suppression in Agroecosystems. Insects. 2019; 10(12):414. https://doi.org/10.3390/insects10120414
Chicago/Turabian StyleNeher, Deborah A., and Mary E. Barbercheck. 2019. "Soil Microarthropods and Soil Health: Intersection of Decomposition and Pest Suppression in Agroecosystems" Insects 10, no. 12: 414. https://doi.org/10.3390/insects10120414
APA StyleNeher, D. A., & Barbercheck, M. E. (2019). Soil Microarthropods and Soil Health: Intersection of Decomposition and Pest Suppression in Agroecosystems. Insects, 10(12), 414. https://doi.org/10.3390/insects10120414