Insecticidal Properties of Capsaicinoids and Glucosinolates Extracted from Capsicum chinense and Tropaeolum tuberosum

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Compound Extraction and Quantification
2.2. Laboratory and Field Bioassays
2.3. Statistical Analysis
3. Results
3.1. Content of Capsaicinoids and Glucosinolates
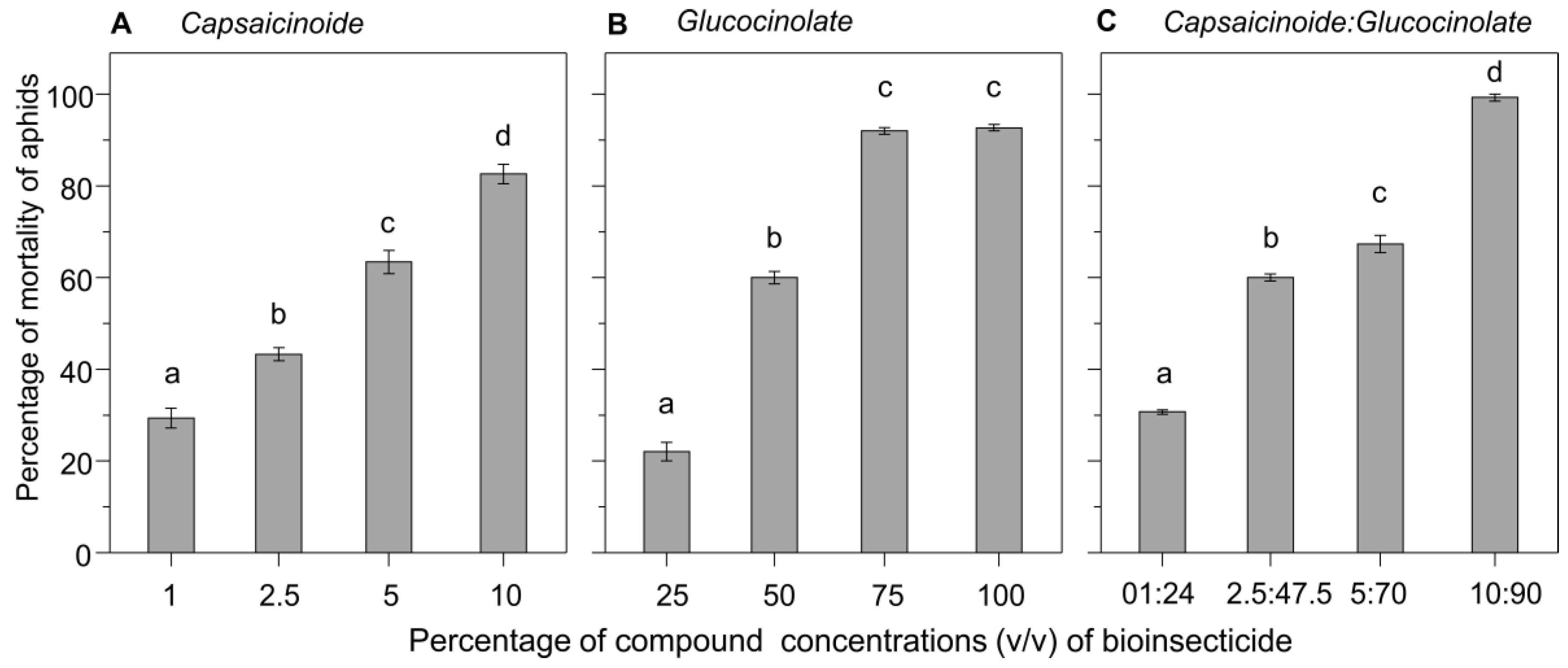
3.2. Toxicity under Laboratory Conditions
3.3. Toxicity under Field Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Bebber, D.P.; Holmes, T.; Gurr, S.J. The global spread of crop pests and pathogens. Glob. Ecol. Biogeogr. 2014, 23, 1398–1407. [Google Scholar] [CrossRef] [Green Version]
- Tito, R.; Vasconcelos, H.L.; Feeley, K.J. Global climate change increases risk of crop yield losses and food insecurity in the tropical Andes. Glob. Chang. Biol. 2018, 24, 592–602. [Google Scholar] [CrossRef]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Bourguet, D.; Guillemaud, T. The Hidden and External Cost of Pesticide Use. In Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer International Publishing: Basel, Switzerland, 2016; Volume 19, pp. 35–120. [Google Scholar]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Dikshit, A.K. Biopesticides: An ecofriendly approach for pest control. J. Biopesticides 2010, 3, 186–188. [Google Scholar]
- Mazid, S.; Kalita, J.C.; Rajkhowa, R.C. A review on the use of biopesticides in insect pest management. Int. J. Sci. Adv. Technol. 2011, 1, 169–178. [Google Scholar]
- Tsao, R.; Reuber, M.; Johnson, L.; Coats, J.R. Insecticidal toxicities of glucosinolate containing extracts from Crambe Seeds. J. Agric. Urban Entomol. 1996, 13, 109–120. [Google Scholar]
- Olszewska, J.; Tegowska, E.; Grajpel, B.; Adamkiewicz, B. Effect of application of capsaicin and pyrethroid on metabolic rate in mealworm Tenebrio molitor. Ecol. Chem. Eng. A 2010, 17, 1355–1359. [Google Scholar]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 2–21. [Google Scholar] [CrossRef]
- Mikkelsen, M.D.; Petersen, B.L.; Olsen, C.E.; Halkier, B.A. Biosynthesis and metabolic engineering of glucosinolates. Amino Acids 2002, 22, 279–295. [Google Scholar] [CrossRef]
- Wen-Zhao, L.; Yi-Deng, D.; Zhang, Y.; Ying-Zhou, C.; Zong-Yi, S.; Wen, C.; Mei-Juan, R.; Hua-Ping, Z. Optimized formation of benzyl isothiocyanate by endogenous enzyme and its extraction from Carica papaya seed. Trop. J. Pharm. Res. 2014, 13, 1303–1311. [Google Scholar]
- Zewdie, Y.; Bosland, P.W. Evaluation of genotype, environment, and genotype-by-environment interaction for capsaicinoids in Capsicum annuum L. Euphytica 2000, 111, 185–190. [Google Scholar] [CrossRef]
- Li, F.; Lin, Y.; Wang, X.; Geng, Y.; Wang, D. Preparative isolation and purification of capsaicinoids from Capsicum frutescens using high-speed counter-current chromatography. Sep. Purif. Technol. 2009, 64, 304–308. [Google Scholar] [CrossRef]
- Sganzerla, M.; Coutinho, J.P.; de Melo, A.M.T.; Godoy, H.T. Fast method for capsaicinoids analysis from Capsicum chinense fruits. Food Res. Int. 2014, 64, 718–725. [Google Scholar] [CrossRef]
- Blackman, R.L.; Easptop, V.F. Aphids on the World’s Crops: An Identification and Information Guide, 2nd ed.; John Wiley and Sons Ltd.: Chicheste, UK, 2000. [Google Scholar]
- Asin, L.; Pons, X. Effect of high temperature on the growth and reproduction of corn aphids (Homoptera: Aphididae) and implications for their population dynamics on the northeastern Iberian peninsula. Environ. Entomol. 2001, 30, 1127–1134. [Google Scholar] [CrossRef]
- Blanchard, S.; Lognay, G.; Verheggen, F.; Detrain, C. Today and tomorrow: Impact of climate change on aphid biology and potential consequences on their mutualism with ants. Physiol. Entomol. 2019. [Google Scholar] [CrossRef]
- Douglas, A.E. The nutritional quality of phloem sap utilized by natural aphid populations. Ecol. Entomol. 1993, 18, 31–38. [Google Scholar] [CrossRef]
- Züst, T.; Agrawal, A.A. Mechanisms and evolution of plant resistance to aphids. Nat. Plants 2016, 2, 1–9. [Google Scholar] [CrossRef]
- Ng, J.C.K.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Ryalls, J.M.W.; Harrington, R. Climate and atmospheric change impacts on aphids as vectors of plant diseases. In Global Climate Change and Terrestrial Invertebrates; Johnson, S.N., Jones, T.H., Eds.; Wiley: Oxford, UK, 2016; pp. 148–175. [Google Scholar]
- Wood, B.W.; Tedders, W.L.; Relly, C.C. Sooty mold fungus on pecan foliage suppresses light penetration and net photosynthesis. Hort. Sci. 1988, 23, 851–853. [Google Scholar]
- Tito, R.; de la Torre-Mayorga, F. Effects of an aphid pest on an invasive plant in the Peruvian Andes. Plant Ecol. 2016, 217, 817–823. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Mallqui, K.V.; Cobián, C.V. Los áfidos (hemiptera: Aphididae) en el callejón de Huaylas—Ancash, Perú. Ecol. Apl. 2011, 10, 93–98. (In Spanish) [Google Scholar] [CrossRef]
- Tambs-Lyche, H. Aphids from the Canary Islands. Entomol. Scand. 1971, 2, 125–131. [Google Scholar] [CrossRef]
- Fernández, M.V.S.; Durante, M.P.M.; Zumeta, J.B.; Nafría, J.M.N. Pulgones (hemiptera: Aphididae) de un sabinar de Juniperus thurifera L. de los Monegros (Zaragoza). Zapateri. Revta. Aragon. Ent. 1998, 8, 113–119. (In Spanish) [Google Scholar]
- Ortego, J.; Difabio, M.E.; Mier Durante, M.P. Nuevos registros y actualización de la lista faunística de los pulgones (Hemiptera: Aphididae) de la Argentina. Rev. Soc. Entomol. Argent. 2004, 63, 19–30. (In Spanish) [Google Scholar]
- Grau, A.; Dueñas, R.O.; Nieto, C.C.; Hermann, M. Mashua (Tropaeolum Tuberosum Ruíz & Pav.). Promoting the Conservation and Use of Underutilized and Neglected Crops; International Plant Genetic Resources Institute (IPGRI), International Potato Center: Lima, Peru, 2003; p. 25. [Google Scholar]
- Bulacio, E.; Ayarde, H. Aspectos ecológicos y distribución de Tropaeolum tuberosum Ssp. silvestre (Tropaeolaceae) en Argentina. Bol. Soc. Arg. Bot. 2012, 47, 97–101. (In Spanish) [Google Scholar]
- Gamarra, M.N.; Roque, L.B. Avances de Investigación en Especies de ajíes del Perú: Evaluación Morfobotánica y Fitoquímica de Especies y Variedades de ajíes (Capsicum) Nativos y Domesticados de la Provincia de Oxapampa, Región Pasco (Perú); CONCYTEC: Lima, Peru, 2012. (In Spanish) [Google Scholar]
- Guillen, N.G.; Tito, R.; Mendoza, N.G. Capsaicinoids and pungency in Capsicum chinense and Capsicum baccatum fruits. Pesqui. Agropecu. Trop. 2018, 48, 237–244. [Google Scholar] [CrossRef]
- Pandhair, V.; Sharma, S. Accumulation of capsaicin in seed, pericarp and placenta of Capsicum annuum L Fruit. J. Plant Biochem. Biotechnol. 2008, 17, 23–27. [Google Scholar] [CrossRef]
- Gudeva, L.K.; Mitrev, S.; Maksimova, V.; Spasov, D. Content of capsaicin extracted from hot pepper (Capsicum annuum ssp. microcarpum L.) and its use as an ecopesticide. Hem. Ind. 2013, 67, 671–675. [Google Scholar] [CrossRef]
- Ramallo, R.; Wathelet, J.; Le Boulengé, E.; Torres, E.; Marlier, M.; Ledent, J.-F.; Guidi, A.; Larondelle, Y. Glucosinolates in isano (Tropaeolum tuberosum) tubers: Qualitative and quantitative content and changes after maturity. J. Sci. Food Agric. 2004, 84, 701–706. [Google Scholar] [CrossRef]
- Al Othman, Z.A.; Ahmed, Y.B.H.; Habila, M.A.; Ghafar, A.A. Determination of capsaicin and dihydrocapsaicin in Capsicum fruit samples using high performance liquid chromatography. Molecules 2011, 16, 8919–8929. [Google Scholar] [CrossRef] [PubMed]
- Juangsamoot, J.; Ruangviriyachai, C.; Techawongstien, S.; Chanthai, S. Determination of capsaicin and dihydrocapsaicin in some hot chilli varieties by RP-HPLC-PDA after magnetic stirring extraction and clean up with C18 cartridge. Int. Food Res. J. 2012, 19, 1217–1226. [Google Scholar]
- Maliszewska, J.; Tęgowska, E. Capsaicin as an organophosphate synergist against colorado potato beetle (Leptinotarsa decemlineata Say). J. Plant Prot. Res. 2012, 52, 28–34. [Google Scholar] [CrossRef]
- Pane, C.; Rongai, D.; Zaccardelli, M. Foliar spray application of glucosinolates and essential oils on processing tomato in open field production system. Agric. Sci. 2013, 4, 149–153. [Google Scholar] [CrossRef]
- Tsao, R.; Peterson, C.J.; Coats, J.R. Glucosinolate breakdown products as insect fumigants and their effect on carbon dioxide emission of insects. BMC Ecol. 2002, 2, 1–7. [Google Scholar] [CrossRef]
- Musso, D.; Cao-Lormeau, V.M.; Gubler, D.J. Zika virus: Following the path of dengue and chikungunya? Lancet 2015, 386, 243–244. [Google Scholar] [CrossRef]
- Marcondes, C.B.; Ximenes, M.D.F.F.D. Zika virus in Brazil and the danger of infestation by Aedes (Stegomyia) mosquitoes. Ver. Soc. Bras. Med. Trop. 2016, 49, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Edelson, J.V.; Duthie, J.; Roberts, W. Toxicity of biorational insecticides: Activity against the green peach aphid, Myzus persicae (Sulzer). Pest Manag. Sci. 2002, 58, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Jeeatid, N.; Techawongstien, S.; Suriharn, B.; Bosland, P.W.; Techawongstein, S. Light Intensity Affects Capsaicinoid Accumulation in Hot Pepper (Capsicum chinense Jacq.) Cultivars. Hortic. Environ. Biotechnol. 2017, 58, 103–110. [Google Scholar] [CrossRef]
- Alexandratos, N.; Bruinsma, J. World Agriculture towards 2015/2030: The 2012 Revision; 12-03, ESA Working Paper; FAO: Rome, Italy, 2012; p. 147. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claros Cuadrado, J.L.; Pinillos, E.O.; Tito, R.; Mirones, C.S.; Gamarra Mendoza, N.N. Insecticidal Properties of Capsaicinoids and Glucosinolates Extracted from Capsicum chinense and Tropaeolum tuberosum. Insects 2019, 10, 132. https://doi.org/10.3390/insects10050132
Claros Cuadrado JL, Pinillos EO, Tito R, Mirones CS, Gamarra Mendoza NN. Insecticidal Properties of Capsaicinoids and Glucosinolates Extracted from Capsicum chinense and Tropaeolum tuberosum. Insects. 2019; 10(5):132. https://doi.org/10.3390/insects10050132
Chicago/Turabian StyleClaros Cuadrado, José L., Elder O. Pinillos, Richard Tito, Carlos Seguil Mirones, and Norma N. Gamarra Mendoza. 2019. "Insecticidal Properties of Capsaicinoids and Glucosinolates Extracted from Capsicum chinense and Tropaeolum tuberosum" Insects 10, no. 5: 132. https://doi.org/10.3390/insects10050132
APA StyleClaros Cuadrado, J. L., Pinillos, E. O., Tito, R., Mirones, C. S., & Gamarra Mendoza, N. N. (2019). Insecticidal Properties of Capsaicinoids and Glucosinolates Extracted from Capsicum chinense and Tropaeolum tuberosum. Insects, 10(5), 132. https://doi.org/10.3390/insects10050132