First Descriptions of Larva and Pupa of Bagous claudicans Boheman, 1845 (Curculionidae, Bagoinae) and Systematic Position of the Species Based on Molecular and Morphological Data


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Morphological Studies
2.2.2. DNA Extraction, Amplification and Sequencing
2.2.3. Sequence and Data Analysis
3. Results
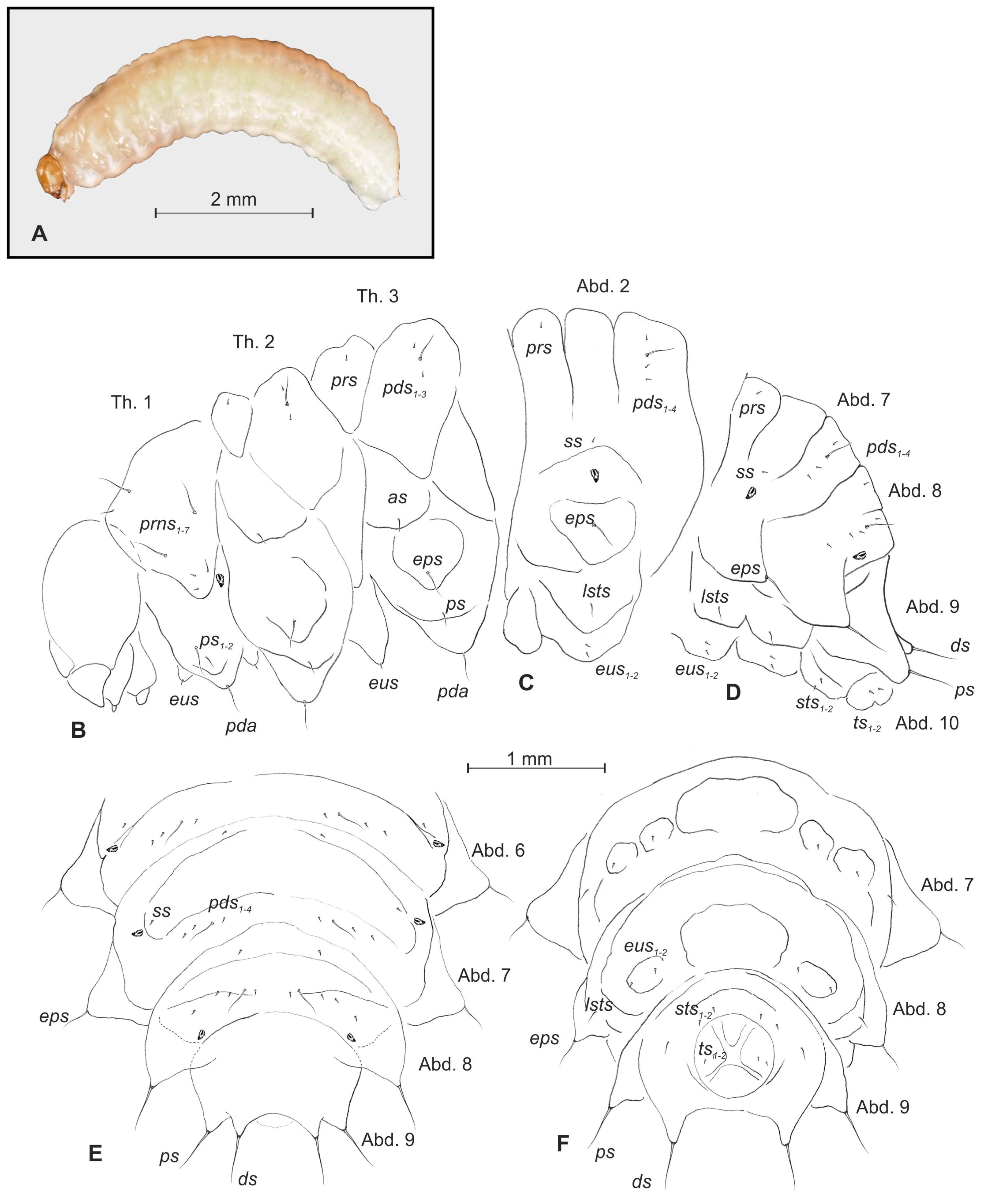
3.1. Description of the Larva of Bagous claudicans
3.2. Description of Pupa of Bagous claudicans
3.3. Genetic Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Alonso-Zarazaga, M.A. Fauna Europaea. Available online: http://www.faunaeur.org (accessed on 4 April 2019).
- Böhme, J. Die Käfer Mitteleuropas, Band K: Katalog (Faunistische Übersicht); 2. Auflage (begründet von Lucht, W.H.); Elsevier GmbH Spektrum: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Bagoinae, C.R. Curculionoidea II. In Catalogue of Palaearctic Coleoptera; Löbl, I., Smetana, A., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2013; Volume 8, pp. 172–176. [Google Scholar]
- Caldara, R.; O’Brien, C.W. Systematics and evolution of weevils of the genus Bagous. VI. Taxonomic treatment of the species of the western Palearctic Region (Coleoptera Curculionidae). Mem. Soc. Entomol. Ital. 1998, 76, 131–347. [Google Scholar]
- Cuppen, J.G.; Heijerman, T. A description of the larva of Bagous brevis Gyllenhal, 1836 (Coleoptera: Curculionidae) with notes on its biology. Elytron 1995, 9, 45–63. [Google Scholar]
- Dieckmann, L. Beiträge zur insektenfauna der DDR: Coleoptera—Curculionidae (tanymecinae, leptopiinae, cleoninae, tanyrhynchinae, cossoninae, raymondionyminae, bagoinae, tanysphyrinae). Beitr. Entomol. 1983, 33, 257–381. [Google Scholar]
- Kodada, J.; Holecova, M.; Behne, L. The genus Dicranthus. I. Taxonomic revision, karyology and notes on the biology (Coleoptera: Curculionidae). Koleopterol. Rundsch. 1992, 62, 195–211. [Google Scholar]
- Oberprieler, R.G.; Caldara, R.; Skuhrovec, J.; Thomson, B.; Lacordaire, G.; Marseul, H. Morphology and systematics (phytophaga). Handbook of zoology: Arthropoda: Insecta. In Coleoptera, Beetles; Leschen, R.A.B., Beutel, R.G., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2014; Volume 3, pp. 452–476. [Google Scholar]
- Marvaldi, A.E.; Sequeira, A.S.; O’Brien, C.W.; Farrell, B.D. Molecular and morphological phylogenetics of weevils (Coleoptera, Curculionoidea): Do niche shifts accompany diversification? Syst. Biol. 2002, 51, 761–785. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.W.; Askevold, I.S. Systematic evolution of weevils of the genus Bagous Germar (Coleoptera: Curculionidae). I. Species of Australia. Trans. Am. Entomol. Soc. 1992, 118, 331–452. [Google Scholar]
- O’Brien, C.W.; Askevold, I.S. Systematic and evolution of weevils of the genus Bagous Germar (Coleoptera: Curculionidae). V. Taxonomic treatment of the species of the Indian Subcontinent. Contrib. Am. Entomol. Inst. 1995, 28, 1–182. [Google Scholar]
- O’Brien, C.W.; Askevold, I.S.; Morimoto, K. Systematic and evolution of weevils of the genus Bagous Germar (Coleoptera: Curculionidae). II. Taxonomic treatment of the species of Japan. Esakia 1994, 34, 1–73. [Google Scholar]
- Caldara, R.; O’Brien, C.W.; Meregalli, M. A phylogenetic analysis of the aquatic weevil tribe Bagoini (Coleoptera: Curculionidae) based on morphological characters of adults. Zootaxa 2017, 4287, 1–63. [Google Scholar] [CrossRef]
- Smreczyński, S. Ryjkowce—Curculionidae: Podrodzina Curculioninae. Klucze do Oznaczania Owadów Polski; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1972; p. 195. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze (Coleoptera) Ryjkowce—Curculionidae, Katalog Fauny Polski; RCiN: Warszawa, Poland, 1995; p. 310. [Google Scholar]
- Gosik, R. Description of the larva and the pupa of Bagous frivaldszkyi Tournier, 1874 (Coleoptera: Curculionidae), with comments on its biology. Belg. J. Entomol. 2006, 8, 107–118. [Google Scholar]
- Gosik, R. Description of the mature larva and pupa of Bagous lutulentus (Gyllenhal.) with comments on its biology. Genus 2009, 20, 125–135. [Google Scholar]
- Scherf, H. Die entwicklungsstadien der mitteleuropäischen curculioniden (morphologie, bionomie, ökologie). Abh. Senckenb. Naturforsch. Ges. 1964, 506, 1–335. [Google Scholar]
- Wanat, M. New records on larval exophagy in the subfamilies Bagoinae and Ceutorhynchinae (Coleoptera: Curculionidae). Weevil News 2011, 67, 1–5. [Google Scholar]
- Gosik, R. Morphology of Larval and Pupal Stages of Selected Species of Bagoinae Thomson, 1895 with Comments on Their Biology and Taxonomy; Wydawnictwo Uniwersytetu Marii Curie–Skłodowskiej: Lublin, Poland, 2013; p. 127. [Google Scholar]
- Skuhrovec, J.; Gosik, R.; Stejskal, R.; Sprick, P.; Caldara, R.; East CURCULIO Team. Digital-weevil-determination for curculionoidea of west palaearctic. Transalpina: Bagous (bagoinae). Snudebiller 2011, 12, 39–56. [Google Scholar]
- Sprick, P. Suitability of an insect group for the habitats directive of the EU: The weevil subfamily bagoinae (Col., Curculionidae). Contributions to the ecology of phytophagous beetles VII. Snudebiller 2001, 2, 7–40. [Google Scholar]
- Wanat, M. Bagous elegans (Fabricius, 1801). In Polska Czerwona Księga Zwierząt—Bezkręgowce; Głowaciński, Z., Nowacki, J., Eds.; Instytut Ochrony Przyrody PAN & Akademia Rolnicza im. A. Cieszkowskiego: Kraków, Poland; Poznań, Poland, 2004; pp. 166–168. [Google Scholar]
- Askevold, I.S.; O’Brien, C.W.; Morimoto, K. Systematics and evolution of weevils of the genus Bagous Germar (Coleoptera, Curculionidae). III. Phylogenetic analysis of the species from Japan. Esakia 1994, 34, 75–129. [Google Scholar]
- Buckingham, G.R.; Bennett, C.A. Host range studies with Bagous affinis (Coleoptera: Curculionidae), an Indian weevil that feeds on Hydrilla tubers. Environ. Entomol. 1998, 27, 469–479. [Google Scholar] [CrossRef]
- Buckingham, G.R. Reunion in Florida—Hydrilla, a weevil, and a fly. Aquatics 1988, 10, 19–25. [Google Scholar]
- Center, T.D.; Parys, K.; Grodowitz, M.; Wheeler, G.S.; Dray, F.A.; O’Brien, C.W.; Johnson, S.; Cofrancesco, A. Evidence of establishment of Bagous hydrillae (Coleoptera: Curculionidae), a biological control agent of Hydrilla verticillata (Hydrocharitales: Hydrocharitaceae) in North America? Fla. Entomol. 2013, 96, 180–186. [Google Scholar] [CrossRef]
- Cock, M.J.W.; Hinz, H.L.; Grosskopf, G.; Häfliger, P. Development of a Biological Control Program for Eurasian Watermilfoil (Myriophyllum spicatum); Aquatic Plant Control Research Program, CABI Europe: Delémont, Switzerland, 2008; p. 33. [Google Scholar]
- O’Brien, C.W.; Pajni, R.H. Two Indian Bagous weevils (Coleoptera, Curculionidae), tuber feeders of Hydrilla verticillata (Hydrocharitaceae), one a potential biocontrol agent in Florida. Entomologist 1989, 72, 462–468. [Google Scholar] [CrossRef]
- Wheeler, G.S.; Center, T.D. Growth and development of the biological control agent Bagous hydrillae as influenced by Hydrilla (Hydrilla verticillata) stem quality. Biol. Control 1997, 8, 52–57. [Google Scholar] [CrossRef]
- Gosik, R. Comparative morphology of the developmental stages of Bagous alismatis (Marsham, 1802) (Coleoptera, Curculionidae) and other Bagous species. Dtsch. Entomol. Z. 2008, 55, 249–262. [Google Scholar] [CrossRef]
- May, B.M. An introduction to the immature stages of Australian Curculionoidea. In Australian Weevils; Apionidae, B.E., Zimmerman, E.C., Eds.; CSIRO: Melbourne, Australia, 1994; Volume 2, pp. 365–659. [Google Scholar]
- Leiler, T.E. Entwicklung und Lebensweise des Rüsselkäfers Bagous frit Hbst. (Col.). Entomol. Blätter 1987, 83, 13–16. [Google Scholar]
- Gosik, R. Description of the larva and the pupa of Bagous nodulosus Gyllenhal in Schoenherr, 1836 (Coleoptera: Curculionidae), with comments on its biology. Balt. J. Coleopterol. 2006, 6, 143–153. [Google Scholar]
- Gosik, R. Morphology of the larval instars of Bagous binodulus (Herbst, 1795) and B. robustus H. Brisout, 1863 with notes on their biology (Coleoptera: Curculionidae). Genus 2010, 21, 293–308. [Google Scholar]
- Mantovani, R.; Galanti, G.; Nocentini, A. Biological observations on Bagous rufimanus Hoffmann (Coleoptera, Curculionidae) with description of its immature stages. Aquat. Insects 1992, 14, 117–127. [Google Scholar] [CrossRef]
- Staniec, B.; Gosik, R. Morphology of the mature larva and pupa of Bagous subcarinatus Gyllenhal, 1836 (Coleoptera: Curculionidae). Genus 2003, 14, 455–464. [Google Scholar]
- Gosik, R.; Wanat, M. First descriptions of immature stages of the weevils Bagous elegans, B. Aliciae and B. Lutulosus. J. Insect Sci. 2014, 14, 1–13. [Google Scholar] [CrossRef]
- Wanat, M. Nowe stanowisko Bagous aliciae Cmoluch, 1983 (Coleoptera, Curculionidae) w Polsce. Wiad. Entomol. 2005, 24, 122–123. [Google Scholar]
- Anderson, W.H. A terminology for the anatomical characters useful in the taxonomy of weevil larvae. Proc. Entomol. Soc. Wash. 1947, 49, 123–132. [Google Scholar]
- Marvaldi, A.E. Higher level phylogeny of Curculionidae (Coleoptera: Curculionoidea) based mainly on larval characters, with special reference to broad–nosed weevils. Cladistics 1997, 13, 285–312. [Google Scholar] [CrossRef]
- Marvaldi, A.E. Larvae of South American Rhytirrhininae (Coleoptera: Curculionidae). Coleopt. Bull. 1998, 52, 71–89. [Google Scholar]
- Marvaldi, A.E. Morfología larval en Curculionidae. Acta Zool. Lilloana 1999, 45, 7–24. [Google Scholar]
- Marvaldi, A.E. Key to larvae of the South American subfamilies of weevils (Coleoptera, Curculionoidea). Rev. Chil. Hist. Nat. 2003, 76, 603–612. [Google Scholar] [CrossRef]
- Skuhrovec, J.; Gosik, R.; Caldara, R.; Košťál, M. Immatures of palaearctic species of the weevil genus sibinia (Coleoptera, Curculionidae): New descriptions and new bionomic data with suggestions on their potential value in a phylogenetic reconstruction of the genus. Zootaxa 2015, 3955, 151–187. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Antennae and sensilla. In Comparative Insects Physiology, Chemistry and Pharmacology 6; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 1–69. [Google Scholar]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals criptic species in the Neotropical skipper bitterfly, Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain–terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Bensasson, D.; Zhang, D.-X.; Hartl, D.L.; Hewitt, G.M. Mitochondrial pseudogenes: Evolution’s misplaced witnesses. Trends Ecol. Evol. 2001, 16, 314–321. [Google Scholar] [CrossRef]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike information criterion and bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J. TreeAnnotator Version 1.10.1. Computer Software and Manual. 2002–2018. Available online: http://beast.bio.ed.ac.uk/treeannotator (accessed on 7 February 2019).
- Guindon, D.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Aassessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Dieckmann, L. Revision der mitteleuropäischen arten der Bagous collignensis—Gruppe (insecta, Coleoptera, Curculionidae: Bagoinae). Reichenbachia 1990, 27, 141–145. [Google Scholar]
- De Meijere, J.C.H. Über in Equisetum parasitierende insekten, Dolerus palustris Kl. und Bagous claudicans Boh. Tijdschr. Entomol. 1912, 55, 208–216. [Google Scholar]
- Wanat, M.; Gosik, R. Materiały do znajomości ryjkowców (Insecta: Coleoptera: Curculionoidea) doliny Bugu. Nowy Pamiet. Fizjogr. 2003, 2, 31–52. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | B. rgillaceus | B. angustus | B. bagdatensis | B. binodulus | B. claudicans | B. collignensis | B. elegans | B. exilis | B. frit | B. glabrirostris | B. limosus | B. longitarsis | B. lutelentus | B. monanthiphagus | B. nodulosus | B. petro | B. robustus | B. subcarinatus |
| B. argillaceus | ||||||||||||||||||
| B. angustus | 31 | |||||||||||||||||
| B. bagdatensis | 36 | 36 | ||||||||||||||||
| B. binodulus | 30 | 38 | 28 | |||||||||||||||
| B. claudicans | 29 | 39 | 36 | 37 | ||||||||||||||
| B. collignensi | 27 | 36 | 36 | 38 | 5 | |||||||||||||
| B. elegans | 29 | 24 | 37 | 30 | 38 | 34 | ||||||||||||
| B. exilis | 24 | 35 | 32 | 28 | 28 | 24 | 33 | |||||||||||
| B. frit | 30 | 41 | 45 | 38 | 22 | 23 | 40 | 30 | ||||||||||
| B. glabrirostris | 27 | 40 | 32 | 16 | 45 | 40 | 34 | 30 | 39 | |||||||||
| B. limosus | 33 | 38 | 32 | 35 | 28 | 24 | 31 | 23 | 32 | 42 | ||||||||
| B. longitarsis | 27 | 38 | 38 | 38 | 12 | 8 | 31 | 26 | 25 | 43 | 27 | |||||||
| B. lutelentus | 32 | 38 | 30 | 18 | 42 | 39 | 33 | 36 | 44 | 21 | 33 | 40 | ||||||
| B. monanthiphagus | 24 | 30 | 32 | 24 | 28 | 28 | 30 | 21 | 28 | 26 | 31 | 30 | 27 | |||||
| B. nodulosus | 27 | 30 | 27 | 31 | 34 | 35 | 32 | 24 | 38 | 35 | 32 | 34 | 32 | 25 | ||||
| B. petro | 27 | 30 | 38 | 31 | 34 | 37 | 26 | 36 | 31 | 34 | 36 | 31 | 32 | 26 | 29 | |||
| B. robustus | 32 | 38 | 37 | 23 | 42 | 42 | 36 | 43 | 40 | 25 | 38 | 40 | 27 | 29 | 37 | 31 | ||
| B. subcarinatus | 25 | 31 | 24 | 25 | 31 | 30 | 36 | 35 | 39 | 27 | 38 | 32 | 30 | 29 | 28 | 37 | 34 | |
| B. tempestivus | 28 | 31 | 27 | 30 | 29 | 27 | 34 | 27 | 35 | 34 | 31 | 30 | 41 | 26 | 31 | 32 | 39 | 31 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gosik, R.; Mazur, M.A.; Sawka-Gądek, N. First Descriptions of Larva and Pupa of Bagous claudicans Boheman, 1845 (Curculionidae, Bagoinae) and Systematic Position of the Species Based on Molecular and Morphological Data. Insects 2019, 10, 166. https://doi.org/10.3390/insects10060166
Gosik R, Mazur MA, Sawka-Gądek N. First Descriptions of Larva and Pupa of Bagous claudicans Boheman, 1845 (Curculionidae, Bagoinae) and Systematic Position of the Species Based on Molecular and Morphological Data. Insects. 2019; 10(6):166. https://doi.org/10.3390/insects10060166
Chicago/Turabian StyleGosik, Rafał, Miłosz A. Mazur, and Natalia Sawka-Gądek. 2019. "First Descriptions of Larva and Pupa of Bagous claudicans Boheman, 1845 (Curculionidae, Bagoinae) and Systematic Position of the Species Based on Molecular and Morphological Data" Insects 10, no. 6: 166. https://doi.org/10.3390/insects10060166
APA StyleGosik, R., Mazur, M. A., & Sawka-Gądek, N. (2019). First Descriptions of Larva and Pupa of Bagous claudicans Boheman, 1845 (Curculionidae, Bagoinae) and Systematic Position of the Species Based on Molecular and Morphological Data. Insects, 10(6), 166. https://doi.org/10.3390/insects10060166