Effect of Diallyl Trisulfide on the Reproductive Behavior of the Grain Moth, Sitotroga cerealella (Lepidoptera: Gelechiidae)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Garlic Compound
2.3. Evaluation of the Fecundity of S. cerealella after DATS Treatment
2.4. Observation of the Circadian Courtship Rhythm of Females and Mating Frequency of S. cerealella after DATS Treatment
2.5. Determination of the Periodicity and Percentage of Calling Behavior of S. cerealella after Treatment
2.6. Observation of Mating Duration of S. cerealella after DATS Treatment
2.7. Effect of DATS on the Amount of Sex Pheromones Secreted by Female S. cerealella
2.8. Statistical Analysis
3. Results
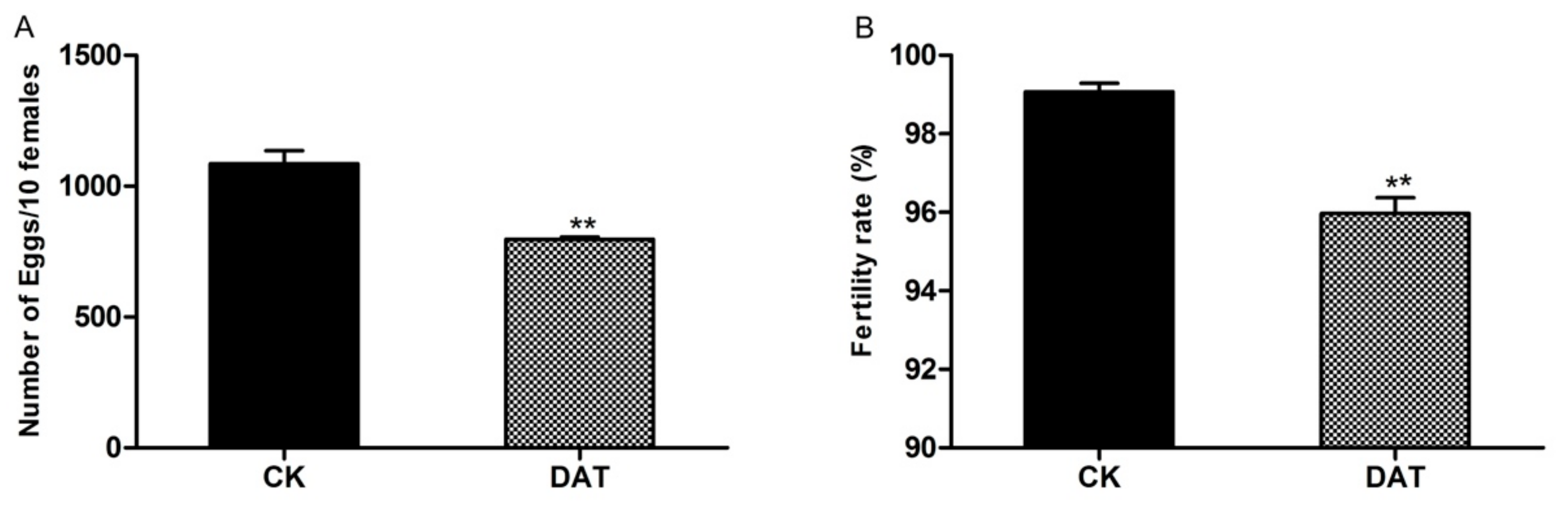
3.1. Efficacy of DATS on the Fecundity of S. cerealella
3.2. Effect of DATS on the Circadian Mating Rhythm and Frequency of Female S. cerealella
3.3. Effect of DATS on the Periodicity and Percentage of Calling of S. cerealella
3.4. Effect of DATS on the Mating Duration of S. cerealella
3.5. Effect of DATS on the Amount of Sex Pheromone Secreted by Female S. cerealella
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ahmed, S.S.; Hashem, M.Y.; El-Sherif, S.I. Comparative effects of different modified atmosphere exposures at 20 and 34 C on the immature stages of angoumois grain moth Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae). J. Stored Prod. Res. 2014, 56, 54–59. [Google Scholar] [CrossRef]
- El-Aziz, S.E.A. Control strategies of stored product pests. J. Entomol. 2011, 8, 101–122. [Google Scholar]
- Boshra, S. Effect of high-temperature pre-irradiation on reproduction and mating competitiveness of male Sitotroga cerealella (Olivier) and their F1 progeny. J. Stored Prod. Res. 2007, 43, 73–78. [Google Scholar] [CrossRef]
- Phillips, T.W.; Throne, J.E. Biorational approaches to managing stored-product insects. Annu. Rev. Entomol. 2010, 55, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Roitberg, B.D. Why pest management needs behavioral ecology and vice versa. Entomol. Res. 2007, 37, 14–18. [Google Scholar] [CrossRef]
- Rafaeli, A.; Gileadi, C. Factors affecting pheromone production in the stored product moth, Plodia interpunctella: A preliminary study. J. Stored Prod. Res. 1995, 31, 243–247. [Google Scholar] [CrossRef]
- Rutowski, R.L. Mate choice and lepidopteran mating behavior. Fla. Entomol. 1982, 65, 72–82. [Google Scholar] [CrossRef]
- Hemptinne, J.-L.; Lognay, G.; Dixon, A. Mate recognition in the two-spot ladybird beetle, Adalia bipunctata: Role of chemical and behavioural cues. J. Insect Physiol. 1998, 44, 1163–1171. [Google Scholar] [CrossRef]
- Wang, Q.; Shi, G. Mating frequency, duration, and circadian mating rhythm of New Zealand wheat bug Nysius huttoni White (Heteroptera: Lygaeidae). N. Z. Entomol. 2004, 27, 113–117. [Google Scholar] [CrossRef]
- Rafaeli, A. Pheromone biosynthesis activating neuropeptide (PBAN): Regulatory role and mode of action. Gen. Comp. Endocrinol. 2009, 162, 69–78. [Google Scholar] [CrossRef]
- Ridley, M. Mating frequency and fecundity in insects. Biol. Rev. 1988, 63, 509–549. [Google Scholar] [CrossRef]
- Edvardsson, M.; Canal, D. The effects of copulation duration in the bruchid beetle Callosobruchus maculatus. Behav. Ecol. 2006, 17, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Foster, S.; Harris, M. Behavioral manipulation methods for insect pest-management. Annu. Rev. Entomol. 1997, 42, 123–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Chen, Z. Non-host plant essential oil volatiles with potential for a ‘push-pull’ strategy to control the tea green leafhopper, Empoasca vitis. Entomol. Exp. Appl. 2015, 156, 77–87. [Google Scholar] [CrossRef]
- Yang, F.L.; Zhu, F.; Lei, C.L. Insecticidal activities of garlic substances against adults of grain moth, Sitotroga cerealella (Lepidoptera: Gelechiidae). Insect Sci. 2012, 19, 205–212. [Google Scholar] [CrossRef]
- Haynes, K.F. Sublethal effects of neurotoxic insecticides on insect behavior. Annu. Rev. Entomol. 1988, 33, 149–168. [Google Scholar] [CrossRef]
- Ma, M.; Chang, M.M.; Lei, C.L.; Yang, F.L. A garlic substance disrupts odorant-binding protein recognition of insect pheromones released from adults of the angoumois grain moth, Sitotroga cerealella (Lepidoptera: Gelechiidae). Insect Mol. Biol. 2016, 25, 530–540. [Google Scholar] [CrossRef]
- Ayertey, J. Egg laying by unmated females of Sitotroga cerealella (Lepidoptera: Gelechiidae). J. Stored Prod. Res. 1975, 11, 211–215. [Google Scholar] [CrossRef]
- Kou, R.; Chow, Y.-S. Calling behavior of the cotton bollworm, Heliothis armigera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 1987, 80, 490–493. [Google Scholar] [CrossRef]
- Zhao, N.N.; Zhang, H.; Zhang, X.C.; Luan, X.B.; Zhou, C.; Liu, Q.Z.; Shi, W.P.; Liu, Z.L. Evaluation of acute toxicity of essential oil of garlic (Allium sativum) and its selected major constituent compounds against overwintering Cacopsylla chinensis (Hemiptera: Psyllidae). J. Econ. Entomol. 2013, 106, 1349–1354. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, R.; Zayed, A.; El-Namaky, A.; Ismail, H.; Mady, H. Biochemical studies on Culex pipiens (L.)(Diptera: Culicidae) exposed to Allium sativum, Citrus limon and Bacillus thuringiensis israelensis with reference to assessment of the biosafety on albino mice. Glob. Vet. 2010, 4, 22–33. [Google Scholar]
- Ramakrishnan, V.; Chintalwar, G.; Banerji, A. Environmental persistence of diallyl disulfide, an insecticidal principle of garlic and its metabolism in mosquito, Culex pipiens quinquifasciatus Say. Chemosphere 1989, 18, 1525–1529. [Google Scholar] [CrossRef]
- Borges, L.M.F.; Sousa, L.A.D.; Barbosa, C.S. Perspectives for the use of plant extracts to control the cattle tick Rhipicephalus (Boophilus) microplus. Rev. Bras. Parasitol. Vet. 2011, 20, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evenden, M.; Haynes, K. Potential for the evolution of resistance to pheromone-based mating disruption tested using two pheromone strains of the cabbage looper, Trichoplusia ni. Entomol. Exp. Appl. 2001, 100, 131–134. [Google Scholar] [CrossRef] [Green Version]
- Tipping, P.; Rodriguez, J.; Poneleit, C.; Legg, D. Feeding activity of the maize weevil (Coleoptera: Curculionidae) on two dent corn lines and some of their mutants. J. Econ. Entomol. 1988, 81, 830–833. [Google Scholar] [CrossRef]
- Slansky, F., Jr. Food consumption and utilization. Compr. Insect Physiol. Biochem. Pharmacol. 1985, 4, 87–163. [Google Scholar]
- Bushra, S.; Aslam, M. Management of Sitotroga cerealella in stored cereal grains: A review. Arch. Phytopathol. Plant Prot. 2014, 47, 2365–2376. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The Principles of Insect Physiology; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar]
- Burpee, D.M.; Sakaluk, S.K. Repeated matings offset costs of reproduction in female crickets. Evol. Ecol. 1993, 7, 240–250. [Google Scholar] [CrossRef]
- Wagner, W.E., Jr.; Kelley, R.J.; Tucker, K.R.; Harper, C.J. Females receive a life-span benefit from male ejaculates in a field cricket. Evolution 2001, 55, 994–1001. [Google Scholar] [CrossRef]
- Tang, Q.Y.; Zhang, C.X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Miyatake, T. Correlated responses to selection for developmental period in Bactrocera cucurbitae (Diptera: Tephritidae): Time of mating and daily activity rhythms. Behav. Genet. 1997, 27, 489–498. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Peak of Mating (Hours into the Scotophase) | |||
|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | Day 4 | |
| CK | 6.20 ± 0.15 | 7.53 ± 0.10 | 8.01 ± 0.14 | 8.20 ± 0.05 |
| DATS | 5.32 ± 0.30 * | 8.04 ± 0.17 * | 8.76 ± 0.25 * | 8.62 ± 0.23 |
| Treatment | Copulation Duration (h) | |||
|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | Day 4 | |
| CK | 4.58 ± 0.25 | 2.39 ± 0.24 | 2.30 ± 0.22 | 2.36 ± 0.20 |
| DATS | 3.59 ± 0.40 * | 1.45 ± 0.32 * | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, M.-M.; Shah, S.; Wu, M.-Y.; Zhang, S.-S.; Wu, G.; Yang, F.-L. Effect of Diallyl Trisulfide on the Reproductive Behavior of the Grain Moth, Sitotroga cerealella (Lepidoptera: Gelechiidae). Insects 2020, 11, 21. https://doi.org/10.3390/insects11010021
Chang M-M, Shah S, Wu M-Y, Zhang S-S, Wu G, Yang F-L. Effect of Diallyl Trisulfide on the Reproductive Behavior of the Grain Moth, Sitotroga cerealella (Lepidoptera: Gelechiidae). Insects. 2020; 11(1):21. https://doi.org/10.3390/insects11010021
Chicago/Turabian StyleChang, Meng-Meng, Sakhawat Shah, Meng-Ya Wu, Su-Su Zhang, Gang Wu, and Feng-Lian Yang. 2020. "Effect of Diallyl Trisulfide on the Reproductive Behavior of the Grain Moth, Sitotroga cerealella (Lepidoptera: Gelechiidae)" Insects 11, no. 1: 21. https://doi.org/10.3390/insects11010021
APA StyleChang, M.-M., Shah, S., Wu, M.-Y., Zhang, S.-S., Wu, G., & Yang, F.-L. (2020). Effect of Diallyl Trisulfide on the Reproductive Behavior of the Grain Moth, Sitotroga cerealella (Lepidoptera: Gelechiidae). Insects, 11(1), 21. https://doi.org/10.3390/insects11010021