Short-Term Cold Stress Affects Parasitism on the Asian Chestnut Gall Wasp Dryocosmus kuriphilus





Simple Summary
Abstract
1. Introduction
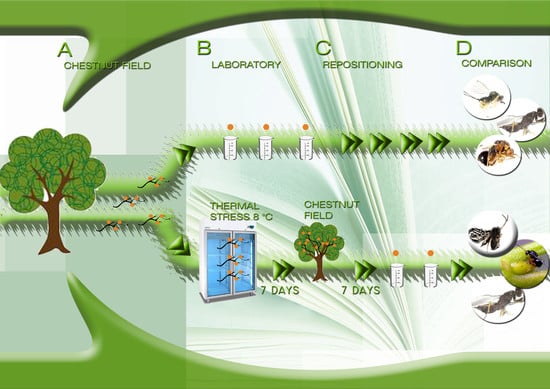
2. Materials and Methods
2.1. Study Area
2.2. Study Design
2.3. Field Sampling and Seasonal Parasitism
2.4. Cold Stress Experiment
2.5. Emergence of the ACGWs and Parasitoids
2.6. Historical Climate Data
2.7. Statistical Analysis
3. Results
3.1. Field Sampling and Seasonal Parasitism
3.2. Cold Stress Experiment
3.3. Emergence of the ACGW and Its Parasitoids
3.4. Historical Climate Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dietz, H.; von Arx, G. Climatic fluctuation causes large scale synchronous variation in radial root increments of perennial forbs. Ecology 2005, 86, 327–333. [Google Scholar] [CrossRef]
- Reusch, T.B.H.; Ehlers, A.; Hämmerli, A.; Worm, B. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. Proc. Natl. Acad. Sci. USA 2005, 102, 2826–2831. [Google Scholar] [CrossRef] [PubMed]
- Rigby, J.R.; Porporato, A. Spring frost risk in a changing climate. Geophys. Res. Lett. 2008, 35, L12703. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Addo-Bediako, A.; Chown, S. Climatic variability and the evolution of insect freeze tolerance. Biol. Rev. Camb. Philos. Soc. 2003, 78, 181–195. [Google Scholar] [CrossRef]
- Blackburn, D.G. Evolution of viviparity in squamate reptiles: Reversibility reconsidered. J. Exp. Zool. Part B 2015, 324, 473–486. [Google Scholar] [CrossRef]
- Klapwijk, M.J.; Grobler, B.C.; Ward, K.; Wheeler, D.; Lewis, O.T. Influence of experimental warming and shading on host-parasitoid synchrony. Glob. Chang. Biol. 2010, 16, 102–112. [Google Scholar] [CrossRef]
- Chen, C.; Gols, R.; Biere, A.; Harve, J.A. Differential effects of climate warming on reproduction and functional responses on insects in the fourth trophic level. Funct. Ecol. 2019, 33, 693–702. [Google Scholar] [CrossRef]
- Tougeron, K.; Brodeur, J.; Le Lann, C.; Baaren, J. How climate change affects the seasonal ecology of insect parasitoids. Ecol. Entomol. 2020, 45, 167–181. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 2008, 11, 1351–1363. [Google Scholar] [CrossRef]
- Renner, S.S.; Zohner, C.M. Climate change and phenological mismatch in trophic interactions among plants, insects, and vertebrates. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 165–182. [Google Scholar] [CrossRef]
- Zhang, C.; McPhaden, M.J. Intraseasonal surface cooling in the equatorial western Pacific. J. Clim. 2000, 13, 2261–2276. [Google Scholar] [CrossRef]
- Abarca, M.; Lill, J.T. Latitudinal variation in the phenological responses of eastern tent caterpillars and their egg parasitoids. Ecol. Entomol. 2019, 44, 50–61. [Google Scholar] [CrossRef]
- Bonsignore, C.P.; Vono, G.; Bernardo, U. Environmental thermal levels affect the phenological relationships between the chestnut gall wasp and its parasitoids. Physiol. Entomol. 2019, 44, 87–98. [Google Scholar] [CrossRef]
- Gilman, S.E.; Urban, M.C.; Tewksbury, J.; Gilchrist, G.W.; Holt, R.D. A framework for community interactions under climate change. Trends Ecol. Evol. 2010, 25, 325–331. [Google Scholar] [CrossRef]
- Plantard, O.; Rasplus, J.W.; Hochberg, M.E. Resource partitioning in the parasitoid assemblage of the oak galler Neuroterus quercusbaccarum (L.) (Hymenoptera: Cynipidae). Acta Oecol. 1996, 17, 1–15. [Google Scholar]
- van der Putten, W.H.; Macel, M.; Visser, M. Predicting species distribution and abundance responses to climate change: Why it is essential to include biotic interactions across trophic levels. Philos. Trans. R. Soc. Lond. B 2010, 365, 2025–2034. [Google Scholar] [CrossRef]
- Johnson, C.A.; Coutinhoc, R.M.; Berlin, E.; Dolphin, K.E.; Heyer, J.; Kim, B.; Leung, A.; Sabellon, J.L.; Amarasekare, P. Effects of temperature and resource variation on insect population dynamics: The bordered plant bug as a case study. Funct. Ecol. 2016, 30, 1122–1131. [Google Scholar] [CrossRef]
- Bonsignore, C.P.; Bernardo, U. Effects of environmental parameters on the chestnut gall wasp and its complex of indigenous parasitoids. Sci. Nat. 2018, 105, 20. [Google Scholar] [CrossRef]
- Torii, T. Studies on the biological control of the chestnut gall wasp, Dryocosmus kuriphilus Yasumatsu (Hym. Cynipidae), with particular reference to the utilization of its indigenous natural enemies. J. Fac. Agric. Shinshu Univ. 1959, 2, 71–149. [Google Scholar]
- Aebi, A.; Schonrogge, K.; Melika, G.; Quacchia, A.; Alma, A.; Stone, G.N. Native and introduced parasitoids attacking the invasive chestnut gall wasp Dryocosmus kuriphilus. EPPO Bull. 2007, 37, 166–171. [Google Scholar] [CrossRef]
- Abe, Y.; Melika, G.; Stone, G.N. The diversity and phylogeography of cynipid gall wasps (Hymenoptera, Cynipidae) of the eastern Palaearctic and their associated communities. Orient. Insects 2007, 41, 196–212. [Google Scholar] [CrossRef]
- Fan, B.; Zhu, D.H. Molecular phylogenetic relationships of Dryocosmus kuriphilus Yasumatsu of different geographical populations based on partial sequences of 16S rRNA gene. J. Hunan Univ. Nat. Sci. 2010, 33, 78–82. [Google Scholar]
- Battisti, A.; Benvegnù, I.; Colombari, F.; Haack, R.A. Invasion by the chestnut gall wasp in Italy causes significant yield loss in Castanea sativa nut production. Agric. For. Entomol. 2014, 16, 75–79. [Google Scholar] [CrossRef]
- Gehring, E.; Bellosi, B.; Quacchia, A.; Conedera, M. Evaluating Dryocosmus kuriphilus-induced damage on Castanea Sativa. Jove J. Vis. Exp. 2018, 138, e57564. [Google Scholar] [CrossRef]
- Yasumatsu, K. A new Dryocosmus injurious to chestnut trees in Japan (Hym., Cynipidae). Mushi 1951, 22, 89–93. [Google Scholar]
- Askew, R.R.; Plantard, O.; Gómez, J.F.; Nieves, M.H.; Nieves-Aldrey, J.L. Catalogue of parasitoids and inquilines in galls of Aylacini, Diplolepidini and Pediaspidini (Hym., Cynipidae) in the West Palaearctic. Zootaxa 2006, 1301, 60. [Google Scholar] [CrossRef]
- Askew, R.R.; Melika, G.; Pujade-Villar, J.; Schoenrogge, K.; Stone, G.N.; Nieves-Aldrey, J.L. Catalogue of parasitoids and inquilines in cynipid oak galls in the West Palaearctic. Zootaxa 2013, 3643, 1–133. [Google Scholar] [CrossRef]
- Ferracini, C.; Bertolino, S.; Bernardo, U.; Bonsignore, C.P.; Faccoli, M.; Ferrari, E.; Lupi, D.; Maini, S.; Mazzone, L.; Nugnes, F.; et al. Do Torymus sinensis (Hymenoptera: Torymidae) and agroforestry system affect native parasitoids associated with the Asian chestnut gall wasp? Biol. Control 2018, 121, 36–43. [Google Scholar] [CrossRef]
- Palmeri, V.; Cascone, P.; Campolo, O.; Grande, S.B.; Laudani, F.; Malacrinò, A.; Guerrieri, E. Hymenoptera wasps associated with the Asian gall wasp of chestnut (Dryocosmus kuriphilus) in Calabria, Italy. Phytoparasitica 2014, 42, 699–702. [Google Scholar] [CrossRef]
- Cooper, W.R.; Rieske, L.K. Gall structure affects ecological associations of Dryocosmus kuriphilus (Hymenoptera: Cynipidae). Environ. Entomol. 2010, 39, 787–797. [Google Scholar] [CrossRef]
- Gehring, E.; Kast, C.; Kilchenmann, V.; Bieri, K.; Gehrig, R.; Pezzatti, G.B.; Conedera, M. Impact of the Asian Chestnut Gall Wasp, Dryocosmus kuriphilus (Hymenoptera, Cynipidae), on the chestnut component of honey in the Southern Swiss Alps. J. Econ. Entomol. 2018, 111, 43–52. [Google Scholar] [CrossRef]
- Zhu, D.; He, Y.; Fan, Y.; Ma, M.; Peng, D. Negative evidence of parthenogenesis induction by Wolbachia in a gallwasp species, Dryocosmus kuriphilus. Entomol. Exp. Appl. 2007, 124, 279–284. [Google Scholar] [CrossRef]
- Bernardo, U.; Iodice, L.; Sasso, R.; Tutore, V.A.; Cascone, P.; Guerrieri, E. Biology and monitoring of Dryocosmus kuriphilus on Castanea sativa in Southern Italy. Agric. For. Entomol. 2013, 15, 65–76. [Google Scholar] [CrossRef]
- Nugnes, F.; Gualtieri, L.; Bonsignore, C.P.; Parillo, R.; Annarumma, R.; Griffo, R.; Bernardo, U. Resistance of a local ecotype of Castanea sativa to Dryocosmus kuriphilus (Hymenoptera: Cynipidae) in Southern Italy. Forests 2018, 9, 94. [Google Scholar] [CrossRef]
- Viggiani, G.; Nugnes, F. Description of the larval stages of Dryokosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae), with notes on their phenology. J. Entomol. Acarol. Res. 2010, 42, 39–45. [Google Scholar] [CrossRef]
- Reale, L.; Tedeschini, E.; Rondoni, G.; Ricci, C.; Bin, F.; Frenguelli, G.; Ferranti, F. Histological investigation on gall development induced by a worldwide invasive pest, Dryocosmus kuriphilus, on Castanea sativa. Plant Biosyst. 2016, 150, 35–42. [Google Scholar] [CrossRef]
- EPPO. Dryocosmus kuriphilus. EPPO Bull. 2005, 35, 422–424. [Google Scholar] [CrossRef]
- Gil-Tapetado, D.; Cabrero-Sañudo, F.; Polidori, C.; Gómez, J.; Nieves-Aldrey, J. Climate as a possible driver of gall morphology in the chestnut pest Dryocosmus kuriphilus across Spanish invaded areas. Bull. Entomol. Res. 2020, 1–14. [Google Scholar] [CrossRef]
- EPPO. New data on quarantine pests and pests of the EPPO Alert List. EPPO Rep. Serv. 2018, 152, 8. [Google Scholar]
- Matošević, D.; Melika, G. Recruitment of native parasitoids to a new invasive host: First results of Dryocosmus kuriphilus parasitoid assemblage in Croatia. Bull. Insectol. 2013, 66, 231–238. [Google Scholar]
- Quacchia, A.; Ferracini, C.; Nicholls, J.A.; Piazza, E.; Saladini, M.A.; Tota, F.; Melika, G.; Alma, A. Chalcid parasitoid community associated with the invading pest Dryocosmus kuriphilus in north-western Italy. Insect Conserv. Divers. 2013, 6, 114–123. [Google Scholar] [CrossRef]
- Panzavolta, T.; Croci, F.; Bracalini, M.; Melika, G.; Benedettelli, S.; Tellini Florenzano, G.; Tiberi, R. Population dynamics of native parasitoids associated with the Asian Chestnut Gall Wasp (Dryocosmus kuriphilus) in Italy. Psyche 2018, 2018. [Google Scholar] [CrossRef]
- Dorado, J.F.; Pujade-Villar, J.; Muñoz-Adalia, E.J.; Vinagrero, J.C.; Diez-Casero, J.J.; Fernández-Fernández, M.M. Characterization of native parasitoid community associated with the invasive pest Dryocosmus kuriphilus (Hymenoptera: Cynipidae) in Cantabria (northern Spain). Scand. J. For. Res. 2020. [Google Scholar] [CrossRef]
- Budroni, M.A.; Loru, L.; Pantaleoni, R.A.; Rustici, M. Effects of an asynchronous alien host on a native host-parasitoid system. Ecol. Complex. 2018, 33, 84–92. [Google Scholar] [CrossRef]
- Gil-Tapetado, D.; Gómez, J.F.; Cabrero-Sañudo, F.J.; Nieves-Aldrey, J.L. Distribution and dispersal of the invasive Asian chestnut gall wasp, Dryocosmus kuriphilus (Hymenoptera: Cynipidae), across the heterogeneous landscape of the Iberian Peninsula. Eur. J. Entomol. 2018, 115, 575–586. [Google Scholar] [CrossRef]
- Bailey, R.; Schönrogge, K.; Cook, J.M.; Melika, G.; Csóka, G.; Thuróczy, C.; Stone, G.N. Host niches and defensive extended phenotypes structure parasitoid wasp communitie. PLoS Biol. 2019, 7, e1000179. [Google Scholar] [CrossRef]
- Cooper, W.R.; Rieske, L.K. A native and an introduced parasitoid utilize an exotic gall-maker host. BioControl 2011, 56, 725–734. [Google Scholar] [CrossRef]
- Cornell, H.V.; Hawkins, B.A. Accumulation of native parasitoid species on introduced herbivores: A comparison of hosts as natives and hosts as invaders. Am. Nat. 1993, 141, 847–865. [Google Scholar] [CrossRef]
- Murakami, Y.; Ao, H.B.; Chang, C.H. Natural enemies of the chestnut gall wasp in Hopei Province, China (Hymenoptera: Chalcidoidea). Appl. Entomol. Zool. 1980, 15, 184–186. [Google Scholar] [CrossRef][Green Version]
- Murakami, Y.; Toda, S.; Gyoutoku, Y. Colonization of imported Torymus (Syntomaspis) sinensis Kamijo (Hymenoptera: Torymidae) parasitic on the chestnut gall wasp (Hymenoptera: Cynipidae). Success in the eighteenth year after release in Kumamoto. Proc. Ass. Plant Protec. Kyush 2001, 47, 132–134. [Google Scholar] [CrossRef]
- Moriya, S.; Inoue, K.; Ôtake, A.; Shiga, M.; Mabuchi, M. Decline of the chestnut gall wasp population, Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) after the establishment of Torymus sinensis Kamijo (Hymenoptera: Torymidae). Appl. Entomol. Zool. 1989, 24, 231–233. [Google Scholar] [CrossRef]
- Borowiec, N.; Thaon, M.; Brancaccio, L.; Warot, S.; Vercken, E.; Fauvergue, X.; Malausa, J.C. Classical biological control against the chestnut gall wasp Dryocosmus kuriphilus (Hymenoptera, Cynipidae) in France. Plant Prot. Q. 2014, 29, 9–12. [Google Scholar]
- Matošević, D.; Lacković, N.; Melika, G.; Kos, K.; Franić, I.; Kriston, E.; Rot, M. Biological control of invasive Dryocosmus kuriphilus with introduced parasitoid Torymus sinensis in Croatia, Slovenia and Hungary. Period. Biol. 2015, 117, 471–477. [Google Scholar] [CrossRef]
- Pérez-Otero, R.; Crespo, D.; Mansilla, J.P. Dryocosmus kuriphilus Yasumatsu, 1951 (Hymenoptera: Cynipidae) in Galicia (NW Spain): Pest dispersion, associated parasitoids and first biological control attempts. Arquivos Entomolóxicos 2017, 17, 439–448. [Google Scholar]
- Gehring, E.; Bellosi, B.; Reynaud, N.; Conedera, M. Chestnut tree damage evolution due to Dryocosmus kuriphilus attacks. J. Pest Sci. 2019. [Google Scholar] [CrossRef]
- Avtzis, D.N.; Melika, G.; Matošević, D.; Coyle, D.R. The Asian chestnut gall wasp Dryocosmus kuriphilus: A global invader and a successful case of classical biological control. J. Pest Sci. 2019, 92, 107–115. [Google Scholar] [CrossRef]
- Ferracini, C.; Ferrari, E.; Pontini, M.; Saladini, M.A.; Alma, A. Effectiveness of Torymus sinensis: A successful long-term control of the Asian chestnut gall wasp in Italy. J. Pest Sci. 2019, 92, 353–359. [Google Scholar] [CrossRef]
- Yara, K.; Yano, E.; Sasawaki, T.; Shiga, M. Detection of hybrids between introduced Torymus sinensis and native T. beneficus (Hymenoptera: Torymidae) in central Japan, using malic enzyme. Appl. Entomol. Zool. 2000, 35, 201–206. [Google Scholar] [CrossRef][Green Version]
- Ferracini, C.; Ferrari, E.; Saladini, M.A.; Pontini, M.; Corradetti, M.; Alma, A. Non-target host risk assessment for the parasitoid Torymus sinensis. BioControl 2015, 60, 583–594. [Google Scholar] [CrossRef]
- Ferracini, C.; Ferrari, E.; Pontini, M.; Hernández Nova, L.K.; Saladini, M.A.; Alma, A. Post-release evaluation of non-target effects of Torymus sinensis, the biological control agent of Dryocosmus kuriphilus in Italy. BioControl 2017, 62, 445–456. [Google Scholar] [CrossRef]
- Askew, R.R. A study of the biology of species of the genus Mesopolobus Westwood (Hymenoptera: Pteromalidae) associated with cynipid galls on oak. Trans. R. Entomol. Soc. Lond. 1961, 113, 155–173. [Google Scholar] [CrossRef]
- de Vere Graham, M.W.R.; Gijswijt, M.J. Revision of the European species of Torymus Dalman (s. Lat.) (Hymenoptera: Torymidae). Zool. Verh. 1998, 317, 1–202. [Google Scholar]
- Askew, R.R.; Nieves-Aldrey, J.L. The genus Eupelmus Dalman, 1820 (Hymenoptera, Chalcidoidea, Eupelmidae) in peninsular Spain and the Canary Islands, with taxonomic notes and descriptions of new species. Graellsia 2000, 56, 49–61. [Google Scholar] [CrossRef]
- Roques, A.; Skrzypczyńska, M. Seed-infesting chalcids of the genus Megastigmus Dalman, 1820 (Hymenoptera: Torymidae) native and introduced to the West Palearctic region: Taxonomy, host specificity and distribution. J. Nat. Hist. 2003, 37, 127–238. [Google Scholar] [CrossRef]
- Al Khatib, F.; Fusu, L.; Cruaud, A.; Gibson, G.; Borowiec, N.; Rasplus, J.Y.; Ris, N.; Delvare, G. An integrative approach to species discrimination in the Eupelmus urozonus complex (Hymenoptera, Eupelmidae), with the description of 11 new species from the Western Palaearctic. Syst. Entomol. 2014, 39, 806–862. [Google Scholar] [CrossRef]
- Gebiola, M.; Bernardo, U.; Burks, R.A. A reevaluation of the generic limits of Pnigalio Schrank (Hymenoptera: Eulophidae) based on molecular and morphological evidence. Zootaxa 2010, 2484, 35–44. [Google Scholar] [CrossRef]
- Guerrieri, E.; Bernardo, U.; Iodice, L.; Gebiola, M. Identificazione morfo-bio-molecolare ed interazioni trofiche degli antagonisti autoctoni di Dryocosmus kuriphilus Yasumatsu in Campania: Metodologia e risultati preliminari. Att. Accad. Naz. Ital. Entomol. 2011, 58, 115–120. [Google Scholar]
- Kaartinen, R.; Stone, G.; Hearn, J.; Lohse, K.; Roslin, T. Revealing secret liaisons: DNA barcoding changes our understanding of food webs. Ecol. Entomol. 2010, 35, 623–638. [Google Scholar] [CrossRef]
- SPSS v23; SPSS Inc.: Chicago, IL, USA, 2015.
- Sigmaplot 13.0; Systat Copyright © Systat Software: San Jose, CA, USA, 2018.
- Jalili, A.; Jamzad, Z.; Thompson, K.; Araghi, M.K.; Ashrafi, S.; Hasaninejad, M.; Panahi, P.; Hooshang, N.; Azadi, R.; Tavakol, M.S.; et al. Climate change, unpredictable cold waves and possible brakes on plant migration. Glob. Ecol. Biogeogr. 2010, 19, 642–648. [Google Scholar] [CrossRef]
- Bernardo, U.; Nugnes, F.; Gualtieri, L.; Scarpato, S.; Gargiulo, G.; Griffo, R. Cinipide del castagno, cresce il controllo biologico in Campania. L’Inf. Agric. 2017, 27, 51–53. [Google Scholar]
- Matošević, D.; Lacković, N.; Kos, K.; Kriston, E.; Melika, G.; Rot, M.; Pernek, M. Success of classical biocontrol agent Torymus sinensis within its expanding range in Europe. J. Appl. Entomol. 2017, 141, 758–767. [Google Scholar] [CrossRef]
- Nieves-Aldrey, J.-L.; Gil-Tapetado, D.; Gavira, O.N.; Boyero, J.R.; Polidori, C.; Lombardero, M.J.; Blanco, D.; Rey del Castillo, C.; Rodríguez-Rojo, M.P.; Vela, J.M.; et al. Torymus sinensis Kamijo, a biocontrol agent against the invasive chestnut gall wasp Dryocosmus kuriphilus Yasumatsu in Spain: Its natural dispersal from France and first data on establishment after experimental releases. For. Syst. 2019, 28, e001. [Google Scholar] [CrossRef]
- Cooper, W.R.; Rieske, L.K. Community associates of an exotic gallmaker, Dryocosmus kuriphilus (Hymenoptera: Cynipidae), in eastern North America. Ann. Entomol. Soc. Am. 2007, 100, 236–244. [Google Scholar] [CrossRef]
- Fernandez-Conradi, P.; Capdevielle, X.; Castagneyrol, B.; Maltoni, A.; Robin, C.; Selvi, F.; Van Halder, I.; Vettillard, F.; Jactel, H. Plant neighbour identity and invasive pathogen infection affect associational resistance to an invasive gall wasp. Biol. Invasions 2017, 20, 1459–1473. [Google Scholar] [CrossRef]
- Gibson, G.A.P.; Gates, M.W.; Buntin, G.D. Parasitoids (Hymenoptera: Chalcidoidea) of the Cabbage Seedpod Weevil (Coleoptera: Curculionidae) in Georgia, USA. J. Hymenopt. Res. 2006, 15, 187–207. [Google Scholar]
- Cusumano, A.; Harvey, J.A.; Dicke, M.; Poelman, E.H. Hyperparasitoids exploit herbivore-induced plant volatiles during host location to assess host quality and non-host identity. Oecologia 2019, 189, 699–709. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Df | FI | p | |
|---|---|---|---|---|
| 2017 | Intercept | 1 | 5135.96 | <0.001 |
| Gall type (Simple, Compound) | 1 | 172.26 | <0.001 | |
| Chestnut field | 2 | 13.55 | <0.001 | |
| Ordinal direction | 3 | 0.89 | 0.45 | |
| Gall type * chestnut field | 2 | 6.30 | 0.02 | |
| 2018 | Intercept | 1 | 3531.15 | <0.001 |
| Gall type (Simple, Compound) | 1 | 119.92 | <0.001 | |
| Chestnut field | 2 | 2.06 | 0.13 | |
| Ordinal direction | 3 | 0.48 | 0.70 | |
| Gall type * chestnut field | 2 | 0.885 | 0.413 |
| Source | Df | Wald Chi-Square | p | |
|---|---|---|---|---|
| 2017 | Intercept | 1 | 296.93 | <0.001 |
| Gall type (Simple, Compound) | 1 | 0.88 | 0.349 | |
| Chestnut field | 2 | 162.75 | <0.001 | |
| Date | 1 | 296.78 | <0.001 | |
| Collection site | 4 | 2.49 | 0.647 | |
| 2018 | Intercept | 1 | 244.93 | <0.001 |
| Gall type (Simple, Compound) | 1 | 0.87 | 0.350 | |
| Chestnut Field | 2 | 21.68 | <0.001 | |
| Date | 1 | 244.98 | <0.001 | |
| Collection site | 3 | 4.22 | 0.239 |
| Year | ||||
|---|---|---|---|---|
| Chestnut Field | 2017 | 2018 | ||
| n | n | |||
| A | 0.18 | 1435 | 0.57 | 1031 |
| B | 0.33 | 1348 | 0.59 | 1048 |
| C | 0.40 | 1001 | 0.67 | 974 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonsignore, C.P.; Vizzari, G.; Vono, G.; Bernardo, U. Short-Term Cold Stress Affects Parasitism on the Asian Chestnut Gall Wasp Dryocosmus kuriphilus. Insects 2020, 11, 841. https://doi.org/10.3390/insects11120841
Bonsignore CP, Vizzari G, Vono G, Bernardo U. Short-Term Cold Stress Affects Parasitism on the Asian Chestnut Gall Wasp Dryocosmus kuriphilus. Insects. 2020; 11(12):841. https://doi.org/10.3390/insects11120841
Chicago/Turabian StyleBonsignore, Carmelo Peter, Giusi Vizzari, Gregorio Vono, and Umberto Bernardo. 2020. "Short-Term Cold Stress Affects Parasitism on the Asian Chestnut Gall Wasp Dryocosmus kuriphilus" Insects 11, no. 12: 841. https://doi.org/10.3390/insects11120841
APA StyleBonsignore, C. P., Vizzari, G., Vono, G., & Bernardo, U. (2020). Short-Term Cold Stress Affects Parasitism on the Asian Chestnut Gall Wasp Dryocosmus kuriphilus. Insects, 11(12), 841. https://doi.org/10.3390/insects11120841