Simple Summary
The strawberry seed bug is a native insect that has recently been causing injury to strawberries in Florida. This study examined the effects of two common cultural practices on the strawberry seed bug: cover cropping and runner removal. Cover cropping is used only in organic strawberry production while runner removal is common to both production systems. The effects of cultivar were also examined. Lastly, small bags were tied around strawberry fruits and different numbers of nymphal and adult seed bugs were released into the bags to examine the effects of their feeding on strawberry fruits. The use of cover crops did not impact seed bug populations. The results for runner removal were inconsistent but it was clear that removing runners does not increase seed bug populations or their injury to fruit. The cultivar “Florida Brilliance” received less injury likely because its seeds are recessed in the flesh of the fruit, making them more difficult for seed bugs to access. Both adults and nymphs caused injury to ripe fruit. Adults did not feed on unripe fruit, so injury to unripe fruit is likely caused by nymphal feeding.
Abstract
The strawberry seed bug, Neopamera bilobata (Say), is an emerging pest of organic and conventional strawberries in Florida. There is limited information on this Rhyparochromidae species. Thus, the type of injury caused is not clearly documented and management recommendations are lacking. In this study, we evaluated the effect of strawberry cultivars, cover crops, and the presence of runners on N. bilobata populations and yield. We also investigated the effect of select cultivars and the presence of runners on N. bilobata injury levels. In addition, we used fruit bagging experiments to investigate the effects of N. bilobata population and life stage (nymph vs. adult) on strawberry fruits. There was no effect of cover crop or cultivar on N. bilobata populations. In the 2017–2018 season, strawberry plots with runners contained higher N. bilobata populations compared with plots without runners, and adult infestation was significantly higher than nymphal infestation. In the 2018–2019 season, the trend was reversed with higher numbers of N. bilobata collected in plots with runners removed. In the 2019–2020 season, there was no significant difference in N. bilobata populations in plots with and without runners. In both 2018–2019 and 2019–2020, nymphal infestation was higher than adult infestation. Less injury was recorded in “Florida Brilliance” compared with the other cultivars tested. In the 2019–2020 season, less injury was recorded from plots without runners while the difference was not significant in 2017–2018 or 2018–2019. Releasing five and ten adult N. bilobata on ripe (red) fruit produced a similar level of injury while no injury to unripe (green) fruit was observed. Both adults and nymphs cause injury to ripe fruit. These findings can help contribute to the development of an integrated pest management program for strawberry N. bilobata.
1. Introduction
Neopamera bilobata Say (Hemiptera: Lygaeoidea: Rhyparochromidae) is native to North America but is also found in South and Central America [1]. It has been recorded from multiple plants, including pinecones, Pinus palustris Mill., [2], torpedograss, Panicum repens L., [2,3], Florida rosemary, Ceratiola ericoides Michx, [4], and figs, Ficus carica L., [5]. In strawberry (Fragaria × ananassa Duch.) fields, N. bilobata shelters under leaves and plastic mulch but disperses quickly when the canopy is disturbed [4]. Neopamera bilobata completes its life cycle on green and ripe strawberry fruit but not on strawberry leaves or flowers [6]. Populations can reach high levels in strawberry fields, as females oviposit about 300 eggs in their lifetime, and development from nymph to adulthood takes an average of 32.8 days on ripe fruit and 36.7 days on green fruit at 23 °C and 70% RH [6].
Research findings from southern Brazil [7] indicated that apical growth of the receptacle was affected by N. bilobata feeding at early fruit stages. Early studies conducted in Florida by Brooks et al. (1929) report that N. bilobata feeding causes drying and brown staining of strawberries in the early fruit stages [8]. In general, Ryparochromidae are mature seed feeders that inject enzymes into the nitrogen-rich portion of the seed so that it becomes liquefied and digestible [9]. Rani et al. (1995) found that the mandibular stylet of the seed predator Odontopus nigricornis (Stall) has barbs, which could help in penetrating hard seed coats, causing circular holes on the seed. Insects belonging to this order frequently cause injury and economic damage to many crops [10]. Currently, there is no economic threshold (ET) or economic injury level (EIL) established for the rhyparochromid seed feeder N. bilobata. The closest related seed predators in strawberries with developed thresholds are tarnished plant bug, Lygus lineolaris (Palisot de Beauvois) and Lygus hesperus (Knight), with an economic threshold of one bug per 20 plants [11].
Cultural practices may be important factors that influence the susceptibility of crops to pests, weeds, and diseases [12]. Cover crops add soil fertility, enhance crop performance, increase biodiversity, and may suppress insect pests [13,14]. In organic Florida strawberry production, summer cover crops are used primarily for nutrient management and weed and plant-pathogenic nematode management (C. Chase, personal observation) but may also affect pests during the following production season. The practice of runner-removal in annual Florida strawberry is common, as runners lead to decreased yields [15]. Daughter-plants from runners are also likely to provide shelter for pests, like N. biolobata, so removing runners has the potential added benefit of disrupting and potentially suppressing N. biolobata populations. This paper is the first to examine the effects of cover crops and runner removal on N. bilobata.
Plant cultivars can vary in their susceptibility to insect pests [16,17,18]. In strawberries, variations in leaf characteristics can affect the twospotted spider mite, Tetraynchus urticae Koch, a major strawberry pest [19]. Fruit produced by different strawberry cultivars can vary in size, sugar content, firmness, and other characteristics [20]. Hata et al. (2020) found higher numbers of N. bilobata on ‘Monterey’ strawberries compared with “Albion” and “San Andres” strawberries [21].
The objectives of this study were to (1) determine if cover crop choice has an effect on N. bilobata populations, (2) examine the effects of cultivar and runner removal on N. bilobata populations and fruit injury and (3) to determine the effects of adult and nymphal feeding on strawberry fruit. We hypothesized that there would be differences in N. bilobata populations and feeding injury among strawberry cultivars, that cover crops would not affect N. bilobata populations, that there would be fewer N. bilobata and less N. bilobata feeding injury in plots where runners were removed, that both adults and nymphs would injure green and ripe strawberry fruit, and that higher numbers of adult seed bugs would cause more injury to ripe strawberry fruits. New information on N. bilobata feeding injury and responses to cultural practices is critical for developing a management program for this important pest.
2. Materials and Methods
2.1. Cover Crops and Strawberry Cultivar
A study on the effects of cover crops and strawberry cultivars on N. bilobata was conducted during the 2016–2019 growing seasons in a certified organic section of the University of Florida, Plant Science Research and Education Unit (PSREU) in Citra, FL. Each season, a split plot design was used, with cover crops (Table 1) as the main plot treatments arranged in a randomized complete block with four replications. The main plots were 9.1 m × 10.7 m rectangles spaced 1.5 m apart in each block with 7.6 m alleys (buffer zones) between blocks. The cover crops were planted on 20 July 2016, 12 July 2017, and 11 July 2018 and were terminated using a flail mower on 30 September 2016, 7 September 2017, and 11 September 2018.

Table 1.
Cover crop treatments.
Strawberry cultivars (Table 2) were the sub plot treatments and were randomized within the main plots after cover crops were terminated. Strawberry plugs (Production Lareault, Inc., QC, Canada) were set in plots on 13 October 2016, 10 October 2017, and on 1 October 2018, at 0.3 m row spacing in raised double row beds covered by black plastic mulch. Each year, Nature Safe® (Darling Ingredients Inc. Irving, TX, USA) 10-2-8 all season fertilizer was applied at the rate of 1681 kg/ha after bed formation and before transplanting strawberries. Weekly fertigation with a 50/50 mix of fish fertilizer (5-1-1; N-P-K) and sodium nitrate (3-0-6; N-P-K) was applied at a rate of 0.58 kg/ha.
Table 2.
Strawberry cultivars planted each season.
Diseases were managed with organically approved products detailed in Table 3. Briefly, in 2017–2018 and 2018–2019, transplant plugs were dipped into a biological fungicide, RootShield® Plus, before planting. In all seasons, two days before predicted rain events, RootShield® Plus was applied through the drip irrigation in combination with a foliar application of an organic approved fungicide. In 2018–2019, Cueva® was applied as a foliar application on 27 November and 4 December 2018 to manage an angular leaf spot outbreak. A preventative release (25 predatory mites per m2) of predatory mites Neoseiulus californicus McGregor (Koppert Biological, Howell, MI, USA) was done in mid-November during each growing season to manage twospotted spider mite, Tetranychus urticae Koch, population growth. When the T. urticae population exceeded a threshold of 10 mites per leaflet, N. californicus was released at the rate of 1 per 10 T. urticae.
Table 3.
List of fungicide products used for disease management and the label rate used for each.
Neopamera bilobata Sampling
In 2016–2017, N. bilobata adults and nymphs were counted in situ in a 15 cm radius around 12 randomly chosen strawberry plants per treatment. In 2017–2018 and 2018–2019, 4 plants were sampled using a 30.5 cm × 30.5 cm beat sheet and in a 15 cm radius around each plant. Neopamera bilobata on the plastic mulch under the plants were counted and then the beat cloth was placed next to one side of the strawberry plant and the plant was shaken for 10 s over the beat sheet and N. biolobata that fell on the sheet were counted. The two numbers were added together for each sample for a total per plant. In 2016–2017 and 2017–2018, N. bilobata were sampled from all 64 subplots. In 2018–19, N. bilobata were sampled from a subset of the subplots (24 out of 64) to assess cultivar effects only because data from the previous two years had indicated that cover crops had no effects on N. bilobata populations.
2.2. Strawberry Runner Removal and Cultivar Experiment
The effects of strawberry cultivar and runner removal on N. bilobata were assessed in field plot experiments in the organic section at the PSREU in Citra, FL during the 2017–2020 growing seasons in a separate plot from the cultivar and cover crops experiment. The plots were arranged in a split-plot design with three replications. The main plots were no runner removal and runner removal. Randomization of the main plots was restricted to prevent the movement of seed bugs between runner and no runner treatment plots. Main plots were 9.1 m × 7.5 m rectangles with a 20 m buffer zone between plots with and without runners. The main plots without runners were trimmed back every two weeks whereas those with runners were not trimmed for the entire experiment.
The subplots were the cultivars, which were randomized within each main plot. Individual subplot size was 9.1 m × 1.5 m with a total of 60 plants in each subplot. In 2019–2020, the plot length was reduced to 6.1 m × 1.5 m with 40 plants in each subplot. Three cultivars were planted each season. Winterstar™, Sensation™ “FL-127”, and “Strawberry Festival” were planted in 2017–2018. “Florida Brilliance”, replaced Winterstar™ during the 2018–2019 and 2019–2020 seasons because Winterstar™ was no longer available. Strawberry plugs were planted on 15 October 2017, 8 October 2018, and 10 October 2019 at 0.3 m row spacing in raised double row beds covered with black plastic mulch. Plants were watered three times a day at 30-min intervals using 2 drip lines for irrigation.
Fertilizer was applied on a per hectare basis through fertigation once a week using a 3-0-6 organic liquid fertilizer (Howard Fertilizer Company, Orlando, FL, USA) at the rate of 69.9 kg of nitrogen per hectare and 139.7 kg of potassium per hectare. Weeds were removed by hand as needed. Serenade® Optimum and Double Nickel® were used when needed to manage diseases through the season (Table 1). No insecticides were applied during the experiment. A preventative release (25 predatory mites per m2) of predatory mites N. californicus was done in mid-November during each growing season to manage T. urticae population growth.
2.2.1. N. bilobata Sampling
Neopamera bilobata were assessed every other week, 20 November through 1 March 2017–2018 and 2018–2019 and 21 November 2019 through 12 March 2020, by vacuuming 15 randomly selected strawberry plants per plot for 10 sec each with a lithium ion handheld vacuum (Stanley Black & Decker, New Britain, CT, USA). Samples were transferred to plastic containers with water (300 mL) and detergent (1%) by removing the side panel of the vacuum, quickly pulling out the filter, and then dumping N. bilobata and other debris into the sample containers. Samples were then taken back to the laboratory where N. bilobata adults and nymphs were sorted and counted.
2.2.2. Yield and Injury Evaluation
Ripe strawberries were harvested twice weekly from mid-November to March each season. Strawberries were counted, weighed, and graded in the field. Fruit was considered cull (unmarketable) if it was too small (<10 g), showed symptoms of disease, or had injury caused by insects, mammals, or abiotic factors. All culled strawberries were examined for missing achenes (seeds) due to N. bilobata feeding and rated by an injury rating system. Culled strawberries with no achenes removed were given an injury rating of 0. Culled strawberries with less than five achenes removed were given an injury rating of 1, those with more than five but less than 10 achenes removed were given an injury rating of 3, and those with more than ten achenes removed were given an injury rating of 5.
2.3. Fruit Bagging Experiments
2.3.1. N. bilobata Colony
An N. bilobata colony was maintained in the laboratory beginning in late February of 2018. The colony was kept in (W30 × D30 × H30 cm) Woven Mesh Polypropylene BugDorm-1 rearing cages (MegaView Science Co., Ltd., Taichung, Taiwan) in an environmental chamber at 25 ± 1 °C, 75% RH and 14 h: 10 h, light: dark. DI water was placed in a souffle cup (50 mL) with a modified cotton wick lid and used as a water source for the insect colony. Twice a week, two or three ripe strawberries from the field were washed in deionized water and placed in the cage as a food source for the insects. Before discarding, each fruit used for feeding the colony, was carefully wiped with a 15.24 cm long plastic paintbrush (Colorations®, Gainesville, FL, USA) to keep all eggs and insects inside the cages.
2.3.2. Adult Density Field Experiment
The effect of adult N. bilobata density on injury to unripe and ripe strawberry fruit was determined using two subplots of Sensation™ strawberries from experiment 2.2. Sensation™ was selected because a high amount of injury was noticed during yield sampling. This experiment was set up as a completely randomized design with split plot restrictions. The main plot treatments were N. bilobata densities of 0, 5, and 10 adults per plant. There were two sub plot treatments, 1 ripe and 1 unripe fruit per plant. A total of 12 plants were used for this experiment and the experiment was conducted twice per season for 2 seasons: 20–25 February 2018, 21–24 February 2018, 7–10 March 2019, and 12–15 March 2019. In 2017–2018, field caught N. bilobata were gathered from a near-by strawberry plot. Field caught N. bilobata adults were chosen at random. In the 2018–2019 season, a total of 180 adult N. bilobata (4 weeks old) from the colony were chosen at random for use in the experiment.
Uninjured full-sized green fruit and ripe red fruit still attached to the peduncle were covered with BugDorm (MegaView Science Co.) insect rearing bags (length 15 cm × width 6 cm). Once all fruit were covered, 0, 5, or 10 adult N. bilobata were released into each rearing bag. After five days, the rearing bags were collected, and injury was observed on the surface of each fruit (360° turn) and assessed using the 0, 1, 3, 5 grading system described previously in Section 2.2.2. Collection of rearing bags was adjusted to 3 days after the first trial in 2017 due to the berries being too ripe after 5 days.
2.3.3. Adult vs. Nymph, Single Ripe Fruit Experiment
The effects of adults and nymphs on ripe fruit was determined in a subplot of Sensation™ strawberries from the runner removal experiment. The experimental design was a randomized complete block design with four replications (7–11 March 2018) and six replications (12–15 March 2019) of three treatments. The treatments were no N. bilobata (control), five adult N. bilobata, and five third-instar N. bilobata. Colony reared N. bilobata adults and third instar nymphs were chosen at random and placed in BugDorm insect rearing bags covering one red healthy (no physical damage or disease) strawberry still attached to the peduncle. After three days, the rearing bags with strawberries were collected and taken back to the laboratory. The entire surface of the strawberry fruits was examined and graded on a scale of 0, 1, 3, and 5 for injury as described in experiment 2.2.2.
2.4. Data Analysis
2.4.1. Cover Crops and Strawberry Cultivar
For the 2016–2017 and 2017–2018 data, the number of N. bilobata per plant was averaged over the entire season and analyzed using a split plot analysis of variance (ANOVA) with the appropriate error term correction for the main plot factor (SAS version 9.4, SAS Institute, Inc., Cary, NC, USA) with cover crop type as the main plot factor and strawberry cultivar as the subplot factor. Means were separated using a least significant difference (LSD) test. In 2018–2019, since only strawberry cultivar was evaluated, N. bilobata per plant data were averaged over the entire season and analyzed using a one-way ANOVA and LSD means separation test. All means were considered significant when p < 0.05.
2.4.2. Strawberry Runner Removal, Cultivar, Injury, and Bag Fruit Data Analysis
A repeated measures analysis using the GLIMMIX procedure with SAS Statistical Software (SAS version 9.4) was used to examine the effects of cultural practice (runners and no runners), selected cultivars, and insect life stage on N. bilobata per plot data. A split plot analysis using the GLIMMIX procedure was used to examine the effects of cultural practice and cultivars on mean total injury rating per plot. Data were examined for normality using the student panel option. A first order autoregressive covariance structure was used for all repeated measures analyses. Least squared means were separated using a Tukey–Kramer test [22]. All means were considered significant when p < 0.05.
The GLIMMIX procedure was used to compare the three densities of N. bilobata to determine effects of N. bilobata population and injury on Sensation™ strawberry (SAS version 9.4). Data were square root transformed. The GLIMMIX procedure was also used to compare adults vs. nymphs to determine which life stage caused more injury.
3. Results
3.1. The Effect of Cover Crops and Strawberry Cultivars on Seed Bug Populations
There was no significant interaction between cover crop and cultivar in 2016–2017 or 2017–2018. In 2016–2017, there were no significant differences in N. bilobata numbers among cover crop (Table 4) treatments or cultivars. In 2017–2018, there were no significant differences in N. bilobata numbers among cover crop treatments but more N. bilobata were found in “Florida Radiance” and WinterstarTM (Figure 1) than in Sensation™ (F = 4.02, df = 3, 36, p = 0.013). In 2018–2019, there was no difference in N. bilobata numbers among cultivars.
Table 4.
Mean ± SEM N. bilobata per plant over all sample dates in each cover crop treatment.
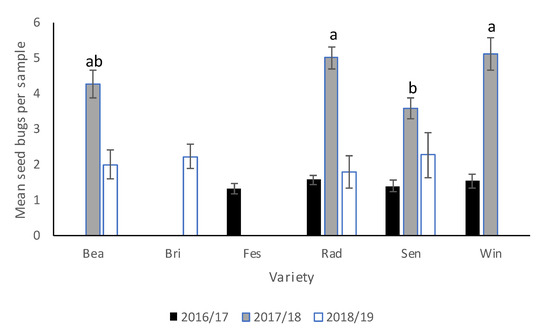
Figure 1.
Mean (±SEM) N. bilobata per plant over the sample dates in each cultivar (Bea = “Florida Beauty”, Bri = “Florida Brilliance”, Fes = “Strawberry Festival”, Rad = “Florida Radiance”, Sen = Sensation™, and Win = WinterstarTM) for all three seasons (2016–2017, 2017–2018, and 2018–2019) in the cover crop trial conducted at the Citra Plant Science Research and Education Unit in organic strawberries. Means with different letters were significantly different at p < 0.05 according to ANOVA and LSD analyses.
3.2. The Effect of Runners and Strawberry Cultivars on Seed Bug Populations, Fruit Injury and Fruit Yield
3.2.1. N. bilobata Populations
We saw no significant interactions between cultivar*runners, cultivar*stage, runners*stage, and cultivar*runners*stage for any of the three seasons, so the main effects were presented. There was no significant difference in N. bilobata numbers among cultivars in 2017–2018, 2018–2019, or 2019–2020 (Figure 2). In 2017–2018, there were significantly more N. bilobata captured in strawberry field plots where runners were not removed compared to plots where runners were removed (F = 16.49; df = 1,50.8; p = 0.0002) (Figure 3). There were approximately 1.7 times more adults captured than nymphs (F = 13.10; df = 1,50.8; p = 0.0007) (Figure 4). In 2018–2019, there were significantly more N. bilobata captured in strawberry field plots where runners were removed compared with plots where runners were not removed (F = 5.73; df = 1,24.8; p = 0.024). There were approximately 3.3 times more nymphs collected than adults (F = 15.81 df = 1,24.8; p = 0.0005). In 2019–2020, there were no differences in N. bilobata numbers among plots with and without runners. There were approximately twice as many nymphs as adults (F = 18.04; df = 1,53.78; p < 0.0001).
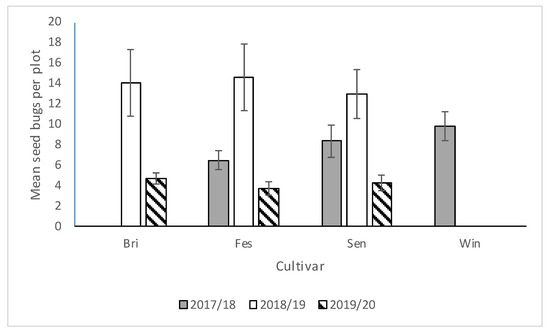
Figure 2.
Mean (±SEM) N. bilobata per plot over the sample dates in each cultivar (Bri = “Florida Brilliance”, Fes = “Strawberry Festival”, Sen = Sensation™, and Win = WinterstarTM) for all three seasons (2017–2018, 2018–2019, and 2019–2020) in the runners vs. no-runners trial conducted at the Citra PSREU in organic strawberries. There were no differences at p < 0.05 according to repeated measures and Tukey’s tests.
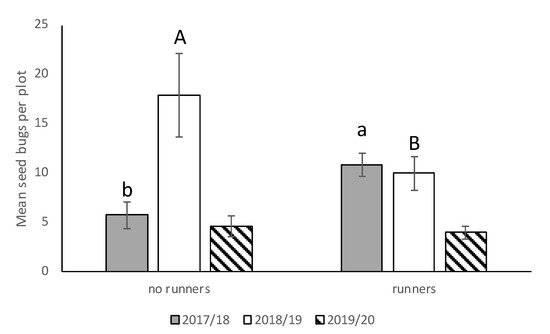
Figure 3.
Mean (±SEM) N. bilobata per plot over the sample dates in plots with and without runners for all three seasons (2017–2018, 2018–2019, and 2019–2020) in the runners vs. no-runners trial conducted at the Citra PSREU in organic strawberries. Means with different lower- or upper-case letters were significantly different at p < 0.05 according to repeated measures and Tukey’s tests.
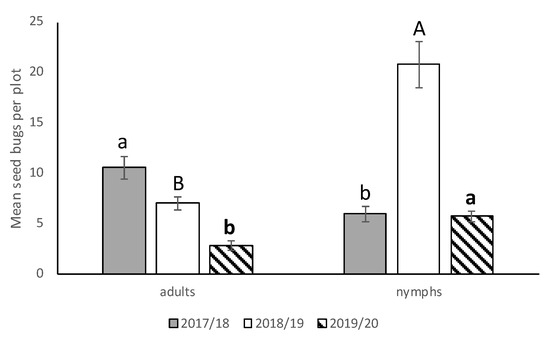
Figure 4.
Mean (±SEM) adult and nymphal N. bilobata per plot over the sample dates for all three seasons (2017–2018, 2018–2019, and 2019–2020) in the runners vs. no runners trial conducted at the Citra PSREU in organic strawberries. Means with different letters were significantly different at p < 0.05 according to repeated measures and Tukey’s tests.
3.2.2. Fruit Injury
In 2017–2018, N. bilobata caused more injury to Sensation™ compared to “Strawberry Festival” and WinterstarTM strawberries (F = 67.49: df = 2,10; p < 0.0001) (Figure 5). No difference in injury occurred in strawberry plots with runners compared to plots with runners removed (F = 0.93; df = 1,10; p = 0.36) (Figure 6). The interaction of cultivar*runners was also significant (F = 4.65; df = 2,10; p = 0.04). Higher injury was seen in SensationTM plots with and without runners compared with all the other treatments (Figure 7).
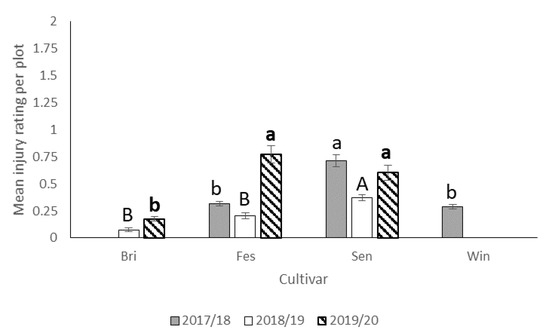
Figure 5.
Mean injury rating (±SEM) in each cultivar for all three seasons (2017–2018, 2018–2019, and 2019–2020) in the runners vs. no runners trial conducted at the Citra PSREU in organic strawberries. Means with different letters were significantly different at p < 0.05 according to repeated measures and Tukey’s tests.
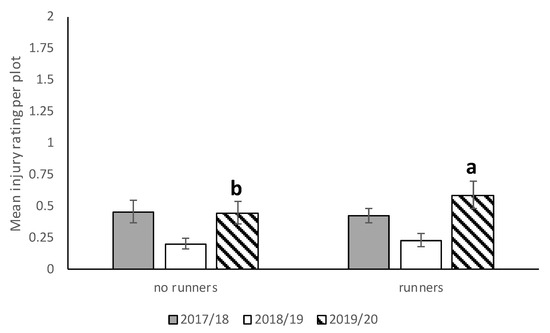
Figure 6.
Mean injury rating (±SEM) in plots with and without runners for all three seasons (2017–2018, 2018–2019, and 2019–2020) in the runners vs. no runners trial conducted at the Citra PSREU in organic strawberries. Means with different letters were significantly different at p < 0.05 according to repeated measures and Tukey’s tests.
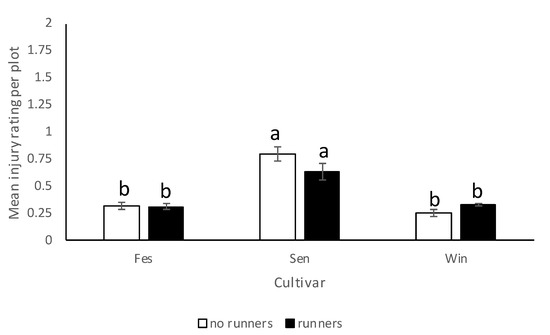
Figure 7.
Mean injury rating (±SEM) in each cultivar in plots with and without runners for the 2017–2018 season in the runners vs. no runners trial conducted at the Citra PSREU in organic strawberries. Means with different letters were significantly different at p < 0.05 according to repeated measures and Tukey’s tests.
During the 2018–2019 season, there was no significant difference between strawberry plots with runners removed and those with runners (Figure 6) still attached with respect to average injury level. The highest N. bilobata injury was observed on Sensation™ fruit (Figure 5) and the lowest on “Florida Brilliance” fruit (F = 29.14; df = 2,12; p < 0.0001). There was no significant interaction for cultivar*runners during the 2018–19 growing season.
During the 2019–2020 season, significantly higher injury was recorded in plots with runners compared (Figure 6) to those without runners (F = 6.53; df = 1,10; p = 0.03). Higher injury was recorded on “Strawberry Festival” and Sensation™ (Figure 5) compared with “Florida Brilliance” (F = 40.68; df = 2,10; p < 0.0001). There was no significant interaction for cultivar*runners during the 2019–2020 growing season.
3.2.3. Yield
In the 2017–2018 season, significant differences were observed among cultivars with respect to marketable yield (F = 13.06; df = 2,12; p = 0.0010). Significantly lower yield was recorded from “Strawberry Festival” compared with Sensation™ and Winterstar™ (Figure 8). Significantly more marketable yield was recorded from strawberry plots with runners removed compared with strawberry plots with runners still attached (Figure 9). Plots that had no runners produced approximately 1.4 times as much marketable fruit yield as those without (F = 14.01; df = 1,12; p = 0.0028). No significant interaction was observed for cultivar*runners (F = 0.71; df = 2,12; p = 0.51) in 2017–2018.
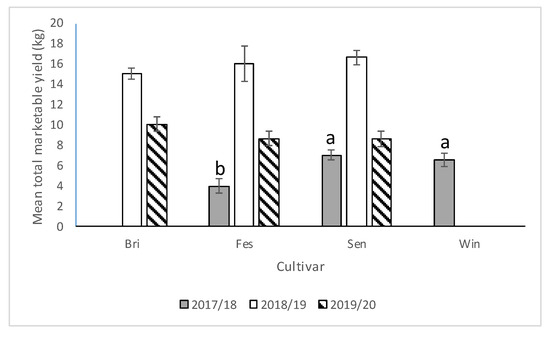
Figure 8.
Mean total marketable yield (±SEM) per cultivar (Bri = “Florida Brilliance”, Fes = “Strawberry Festival”, Sen = Sensation™, and Win = WinterstarTM) for all three seasons (2017–2018, 2018–2019, and 2019–2020) in the runners vs. no runners trial conducted at the Citra PSREU in organic strawberries. Means with different letters were significantly different at p < 0.05 according to repeated measures and Tukey’s tests.
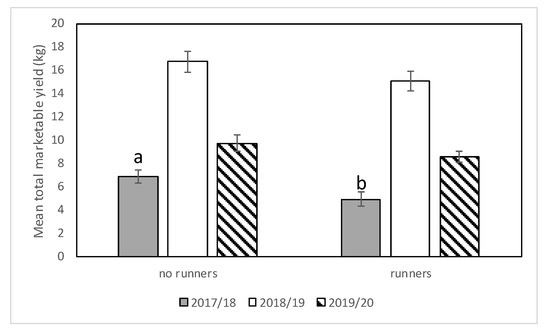
Figure 9.
Mean total marketable yield (±SEM) in plots with and without runners for all three seasons (2017–2018, 2018–2019, and 2019–2020) in the runners vs. no runners trial conducted at the Citra PSREU in organic strawberries. Means with different letters were significantly different at p < 0.05 according to repeated measures and Tukey’s tests.
In the 2018–2019 season, there was a significant interaction between cultivar and cultural practice (F = 5.34; df = 2,10; p = 0.0265) but no differences between plots with and without runners (F = 4.05; df = 1,10; p = 0.07) nor among cultivars (F = 1.32; df = 2,10; p = 0.31). Average marketable strawberry yield (kg/per plot) was significantly higher in “Strawberry Festival” plots where runners were removed compared with “Strawberry Festival” plots where runners were still attached (Figure 10). However, no significant difference in average marketable weight (kg/per plot) was observed for “Florida Brilliance” or Sensation™ with or without runners.
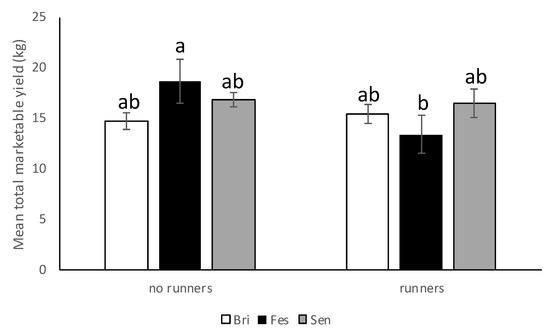
Figure 10.
Mean total marketable yield (±SEM) per cultivar (Bri = “Florida Brilliance”, Fes = “Strawberry Festival”, Sen = Sensation™, and Win = WinterstarTM) in plots with and without runners for the 2018–19 season in the runners vs. no-runners trial conducted at the Citra PSREU in organic strawberries. Means with different letters were significantly different at p < 0.05 according to repeated measures and Tukey’s tests.
In the 2019–2020 season, as in the 2017–2018 season, no significant interaction between cultivar and presence of runners was observed (F = 0.83; df = 2,12; p = 0.46). There were no differences in marketable yield among cultivars (F = 1.2; df = 2,12; p = 0.33) nor between plots with and without runners (F = 1.9; df = 1,12; p = 0.19) (Figure 8 and Figure 9).
3.3. Fruit Bagging Experiments
3.3.1. Adult Density Experiment
There were no significant interactions between season and density, and thus data were averaged across both seasons. During the 2017–2018 and 2018–2019 seasons (Table 5), injury was significantly higher on red fruit compared with green fruit (F = 22.45; df = 2,22.5; p < 0.0001). With red fruit, injury incidence was higher with N. bilobata densities of five and ten when compared with the control. The control without N. bilobata did not have any injury. Zero green fruit were affected.
Table 5.
Mean ± SEM injury rating per fruit for the adult density experiment. Means with different letters were significantly different at p < 0.05 according to ANOVA and Tukey’s tests.
3.3.2. Effects of Adults and Nymphs on Mature Fruit
In the 2017–2018 season (Table 6), significantly more injury was caused by adult N. bilobata compared with nymphs and by both adults and nymphs compared with the control treatment (F = 31.42; df = 2,33.7; p < 0.0001). In the 2018–2019 season, there was no significant difference in injury between adults and nymphs, but a significant difference for both treatments compared with the control (mean injury rating 0 ± 0) (F = 12.40; df = 2,63.1; p < 0.0001).
Table 6.
Mean ± SEM injury rating per fruit for the adults vs, nymphs experiment. Means with different letters were significantly different at p < 0.05 according to ANOVA and Tukey’s tests.
4. Discussion
This study examined the effects of cultivar, cover crops, and runner removal on N. bilobata populations. The effects of cultivar, runner removal, developmental stage, and numbers of N. bilobata on peudofruit injury were also examined. As hypothesized, there was no effect of the cover crop on N. bilobata populations, there were differences in fruit injury among cultivars, and both adult and nymphal N. bilobata caused injury to ripe fruit. Contrary to our hypothesis, there was no effect of cultivar on N. bilobata populations and adult N. bilobata did not injure green fruit. The effect of runner removal on N. bilobata populations varied among the three seasons. Fruit injury was either no different or lower in plots where runners were removed depending on the season.
Strawberry cultivar had no effect on N. bilobata populations with one exception, the presence of fewer N. bilobata on Sensation™ plants in 2017–2018 in the cover crop trial. Though statistically significant, the difference was only 1 N. bilobata per plot, which is likely not ecologically relevant. Another factor could have been the sampling methods used. The vacuum collects N. bilobata from the entire plant and the surrounding mulch and in-situ counts examined the same things. A more specific sampling method might have revealed differences. For example, [21] found differences in N. bilobata populations on different strawberry cultivars when they counted N. bilobata in-situ specifically on fruit.
In terms of N. bilobata injury to fruit, however, there were cultivar differences. “Florida Brilliance” had less injury than the other cultivars. This could be due to the orientation of the achenes on the fruit. The achenes of “Florida Brilliance” are positioned below the fruit surface [23], which may make them less accessible to the seed bug. In contrast, “Strawberry Festival”, Sensation™, and WinterstarTM have similarly positioned achenes on the surface of the fruit that are easier for N. bilobata to access. The abundance and feeding study conducted by Rhainds et al. (2003) found that injury by L. lineolaris in strawberry fields varied greatly by cultivar [24].
There was no effect of the cover crop treatment on N. bilobata populations. Cover crops are terminated and then incorporated into the soil before the main crop, strawberry in this case, is planted. Therefore, the use of cover crops primarily affects soil dwelling pests like plant parasitic nematodes. Adult N. bilobata were not observed in strawberry fields until at least December when fruit was present in the field. It is likely that they moved from other hosts into strawberries once the fruit became available, which is perhaps the reason why cover crops had no effect on their populations.
Our findings on the effect of runners on seed bug populations varied by season. During 2017–2018, N. bilobata populations were higher in field plots where runners remained attached to the strawberry plant while in 2018–2019, N. bilobata populations were higher in plots where runners were removed. There were no differences in N. bilobata populations in 2019–2020. Neopamera bilobata injury was also higher in plots with runners compared to plots where runners were removed only in 2019–2020. Oghiakhe et al. (1991) found that vegetative growth such as canopy played a role in the infestation of cowpea, Vigna unguiculata (L.) Walp, by Maruca testulalis Geyer [25]. Similarly, the increased canopy cover and shelter provided by the presence of daughter plants from runners may result in protection for N. bilobata from predators and the environment (e.g., freezing, precipitation) [26]. Connecting corridors or habitat patches with interconnections, like runners could also be important for N. bilobata nymphal stages because they are wingless and have short dispersal distances [27]. Several factors could have contributed to the variation in results among seasons. Changes in production or pest management practices in a crop, such as replacing WinterstarTM with “Florida Brilliance”, can profoundly alter the pest’s population dynamics causing shifts in pest populations [28,29]. Another change that could have affected seed bug populations was that the strawberry field was moved ~100 m) to the east between the 2017–2018 and 2018–2019 seasons. This change in location could have changed the dispersal pattern of N. bilobata into the field. Pasek (1998) found that patterns of insect population can be affected by local fields, wind speed, insect behavior, and vegetative composition [30].
Higher numbers of adults compared with nymphs were captured in 2017–2018 field-season. Higher adult captures could have been due to the ability of adults to withstand more adverse weather conditions due to their highly sclerotized and pruinose body surface [31,32,33], and/or lack of predators at this life stage. Adults may also have flourished because field conditions in 2017–18 were ideal with relative humidity above 65%, temperature above 25 °C, and an abundant food source [6,34,35]. Our findings are consistent with the assertion in a review by Kennedy and Storer (2000) [36] that habitat suitability for insects is dramatically affected by different factors, such as predators [37] and weather [38].
In contrast, higher numbers of nymphs were captured in 2018–2019 and 2019–2020. Research with the hemipteran insect Lygaeus equestris indicates that parental and offspring populations vary between habitat each year [39]. When conditions are harsh (e.g., weather, host quality), female arthropods tend to compensate by laying more eggs [40]. This was shown in work done by Norhisham et al. (2014) with the milkweed bug Oncopeltus fasciatus Dallas, where females must locate an adequate supply of high quality seeds before reproduction can occur because nymphal growth rates depend on the presence of seeds and their nutritional quality [40]. Since N. bilobata feed on strawberry achenes, it is possible that fruit quality affected host preference and abundance [41,42]. In the future, more studies need to be conducted on oviposition preference and behavior of N. bilobata.
The results from field studies conducted in the 2017–2018 season showed that Winterstar™ and Sensation™ cultivars produced higher yields than “Strawberry Festival” and there were higher yields in plots where runners were removed. In 2018–2019, there was an interaction between cultivar and cultural practice, where “Strawberry Festival” produced higher yields when runners were removed than when they remained attached. There were no differences in yield among cultivars nor between plots with and without runners in 2019–2020. Runner removal is a common practice in Florida strawberry production because it results in higher yields [43]. Albregts and howard (1986) reported that removing runners increased total marketable yields of “Albion” strawberries in each planting year [44]. Runner removal can reduce competition within the plant for resources between runner production and flowering, consequently improving yields [26,44].
The field density studies conducted in the 2017–2018 and 2018–2019 seasons indicated no statistical difference in mean injury rating inflicted by five or ten N. bilobata although the mean injury rating was numerically higher for ten N. bilobata per fruit. On caged Gossypium hirsutum L. plants, [45] found that when densities of Creontiades signatus Distant (Hemiptera: Miridae) were increased from 0.5 to 4 bugs per plant, cotton boll injury was increased. Several other studies indicate a decrease in yield when Hemipteran density increases [46,47].
It appears that adult N. bilobata do not cause injury to green fruit. This could be due to the change in fruit physiology as N. bilobata may need achene development at a certain stage for feeding. Fait et al. (2008) identified changes in the primary and secondary metabolites for the receptacle and achenes of strawberries, where they found major fluctuations in primary compounds in the achene like glucose, aspartate, anthocyanin synthase, and tricarboxylic acid or TCA [48]. The secondary metabolite that was in high fluctuations in the achene was catechin phenylpropanoid. It is possible that these changes within the achenes could be detected by the sensory organs of the adult N. biolbata and, consequently, cause the adult N. biolbata to feed on mature (ripe) fruit. The phenology of the achene may also play a role in why more injury occurred on ripe fruit compared with the green. The authors of [4] observed adult N. bilobata feeding on small dry fruit seeds instead of fleshy fruit seeds in Florida rosemary. More research, including choice experiments and gut content analysis, would need to be conducted to determine seed bug feeding behavior.
Adult N. bilobata caused more injury to ripe strawberries than 3rd instar nymphs in 2017–2018 but there were no differences in adult and 3rd instar nymph injury during the 2018–2019 season. Results from studies conducted by [6] showed that nymphal seed bugs could complete their life cycle on green fruit. Another seed predator study found that Edessa meditabunda (F.) nymphs did not reach the third instar on mature soybean pod seeds or on mature sunflower seeds [49]. It is possible that nymphs may prefer feeding on green immature achenes over mature achenes, whereas adults prefer to feed on ripe achenes compared to green.
5. Conclusions
Our research showed that the use of summer cover crops does not appear to affect N. bilobata populations. The effects of removing runners were variable among seasons, but the practice is not likely to increase fruit injury caused by N. bilobata. Therefore, both practices can continue to be used even on farms where N. bilobata is economically damaging. However, on farms where N. bilobata populations are high, planting cultivars with recessed achenes like “Florida Brilliance” may reduce fruit injury. Both N. bilobata nymphs and adults cause injury to ripe strawberries in Florida and nymphs are also capable of injuring unripe strawberries. Therefore, both life stages should be targeted for management to prevent economic damage.
Author Contributions
Conceptualization, O.E.L. and H.R.T.; methodology, O.E.L., H.R.T., E.M.R., and J.M.R.; formal analysis, H.R.T. and E.M.R.; investigation, H.R.T. and E.M.R.; resources, C.A.C., M.E.S., O.E.L., and J.M.R.; data curation, O.E.L.; writing—original draft preparation, H.R.T. and E.M.R.; writing—review and editing, O.E.L., J.M.R., C.A.C., and M.E.S.; visualization, H.R.T. and E.M.R.; supervision, O.E.L., C.A.C., and M.E.S.; project administration, O.E.L. and C.A.C.; funding acquisition, C.A.C., O.E.L., J.M.R., and M.E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by the Organic Agriculture Research and Extension Initiative grant no. 2015-51300-24134/project accession no. 1007441 from the USDA National Institute of Food and Agriculture.
Acknowledgments
The authors thank all the staff and students of the Small Fruit and Vegetable IPM Laboratory for help with sample collection and processing. Thanks to the staff of the Plant Science Research and Education Unit for plot preparation and maintenance. Thank you to Rachel Mallinger for review of early drafts of this manuscript and to James Colee of UF/IFAS Statistics for statistical consulting.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dellape, P.M. Biodiversidad, Relationships Genetics and Biogeographic Aspects of Rhyparochromidae (Lygaeoidea: Heteroptera) with Special Reference to the Genus Neopamera Harrington 1980. Master’s. Thesis, Facultad de Ciencias Naturales y Museo, National University of La Silver, La Plata, Argentina, 2005. [Google Scholar]
- Wheeler, A.G., Jr.; Stoops, C.A. Cnemodus hirtipes Blatchley and C. mavortius (Say) (Hemiptera: Lygaeoidea: Rhyparochromidae) in fallen pinecones with consideration of the biological significance of cone occupancy. Proc. Entomol. Soc. Wash. 2010, 112, 155–168. [Google Scholar] [CrossRef]
- Cuda, J.P.; Dunford, J.C.; Leavengood, J.M., Jr. Invertebrate fauna associated with torpedograss, Panicum repens (Cyperales: Poaceae), in Lake Okeechobee, Florida, and prospects for biological control. Fla. Entomol. 2007, 90, 238–248. [Google Scholar] [CrossRef]
- Wheeler, A.G. Neopamera bilobata (Say) and Ozophora trinotata Barber (Hemiptera: Lygaeoidea: Rhyparochromidae) as frugivores on Florida Rosemary (Ceratiola ericoides; Ericaceae). Proc. Entomol. Soc. Wash. 2016, 118, 424–435. [Google Scholar] [CrossRef]
- Slater, J.A. Lygaeid bugs (Hemiptera: Lygaeidae) as seed predators of figs. Biotropica 1972, 4, 145–151. [Google Scholar] [CrossRef]
- Kuhn, T.M.; Loeck, A.E.; Zawadneak, M.A.; Garcia, S.; Button, M. Parameters biological and fertility life table of Neopamera bilobata (Hemiptera: Rhyparochromidae) in strawberry. Brasilia 2014, 49, 422–427. [Google Scholar]
- Botton, M.; Kuhn, T.M.; Zawadneak, M.A.C.; Loeck, A.E. Bioecology and characterization of Neopamera bilobata (Say 1832) (Hemiptera: Rhyparochromidae) damage in strawberry. Comun. Tec. Embrapa Uva Vinho 2016, 194, 10. [Google Scholar]
- Brooks, A.N.; Watson, J.M.R.; Mowry, H. Strawberries in Florida: Culture, Diseases, and Insects; University of Florida Agricultural Experiment Station: Gainesville, FL, USA, 1929; p. 523. [Google Scholar]
- Brambila, J.; Hodges, G.S. Bugs (Hemiptera). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; Volume 2, pp. 591–611. [Google Scholar]
- Rani, P.U.; Madhavendra, S.S. Morphology and distribution of antennal sense organs and diversity of mouthpart structures in Odontopus nigricornis (Stall) and Nezara viridula L. (Hemiptera). Int. J. Insect. Morphol. Embryol. 1995, 24, 119–132. [Google Scholar] [CrossRef]
- Burrack, H.; Toennison, A. Tarnished Plant Bug in Strawberries. NC State Extension Publications; University of North Carolina State Cooperative Extension: Raleigh, NC, USA, 2014. [Google Scholar]
- White, J.C.; Liburd, O.E. Effect of soil moisture and temperature on reproduction and development of twospotted spider mite (Acari: Tetranychidae) in strawberries. J. Econ. Entomol. 2005, 98, 154–158. [Google Scholar] [CrossRef]
- Ingels, C.; van Horn, M.; Bugg, R.; Miller, P.R. Selecting the right cover crop gives multiple benefits. Calif. Agric. 1994, 48, 43–48. [Google Scholar] [CrossRef]
- McNeill, C.A.; Liburd, O.E.; Chase, C.A. Effect of cover crops on aphids, whiteflies, and their associated natural enemies in organic squash. J. Sustain. Agric. 2012, 36, 382–403. [Google Scholar] [CrossRef]
- Buckley, B., III; Moore, J.N. The effects of bed height, bed width, plant spacing, and runner removal on strawberry yield and fruit size. Adv. Strawb. Prod. 1982, 1, 4–5. [Google Scholar]
- Kordan, B.; Wroblewska-Kurdyk, A.; Bocianowski, J.; Ster, K.; Jankowski, K.; Gabrys, B. Variation in susceptibility of rapeseed cultivars to the peach potato aphid. J. Pest Sci. 2020. [Google Scholar] [CrossRef]
- Rhodes, E.M.; Liburd, O.E.; England, G.K. Effects of southern highbush blueberry cultivar and treatment threshold on flower thrips populations. J. Econ. Entomol. 2012, 105, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Raina, A.; Thindwa, H.; Othieno, S.; Douglass, L. Resistance in sorghum to sorghum shoot fly (Diptera: Muscidae) oviposition on selected cultivars. J. Econ. Entomol. 1984, 77, 648–651. [Google Scholar] [CrossRef]
- Figueiredo, A.S.T.; Resende, J.T.V.; Morales, R.G.F.; Gonçalves, A.P.S.; Da Silva, P.R. The role of glandular and non-glandular trichomes in the negative interactions between strawberry cultivars and spider mite. Arthropod-Plant Interact. 2013, 7, 53–58. [Google Scholar] [CrossRef]
- Childers, N.F. (Ed.) The Strawberry a Book for Growers, Others; E. O. Painter Printer Company Inc.: DeLeon Springs, FL, USA, 2003; p. 246. [Google Scholar]
- Hata, F.T.; Ventura, M.U.; Ferreira, N.Z.B.; Bega, V.L.; Camacho, I.M.; de Paula, M.T.; Machado, R.R.; de Lima, D.P. Neopamera bilobata Say (Hemiptera: Rhyparochromidae) habit and density on strawberry cultivars. Phytoparasitica 2020, 48, 183–190. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT Users Guide; Version 9.4; SAS Institute: Cary, NC, USA, 2014. [Google Scholar]
- Whitaker, V.M.; Peres, N.A.; Agehara, S. ‘Florida Brilliance’ Strawberry. University of Florida IFAS Extension Publication #HS1322; University of Florida: Gainesville, FL, USA, 2018. [Google Scholar]
- Rhainds, M.; English-Loeb, G. Variation in Abundance and Feeding Impact of Tarnished Plant Bug (Hemiptera: Miridae) for Different Cultivars of Strawberry: Role of Flowering Phenology and Yield Attributes. J. Econ. Entomol. 2003, 96, 433–440. [Google Scholar] [CrossRef]
- Oghiakhe, S.; Jackai, L.E.N.; Makanjuola, W.A. Cowpea plant architecture in relation to infestation and damage by the legume pod borer, Maruca testulalis Geyer (Lepidoptera: Pyralidae)—I. Effect of canopy structure and pod position. Int. J. Trop. Insect Sci. 1991, 12, 193–199. [Google Scholar] [CrossRef]
- Lyu, C.B.; Yang, W.J.; Li, K.T. Partial defoliation and runner removal affect runnering, fruiting, leaf photosynthesis and root growth in ‘Toyonoka’ strawberries for subtropical winter production. Hortic. Environ. Biotechnol. 2014, 55, 372–379. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape ecology: The effect of pattern on process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Hardee, D.D.; Bryan, W.W. Influence of Bacillus thuringiensis-transgenic plants and nectariless cotton on insect populations with emphasis on the tarnished plant bug (Heteroptera: Miridae). J. Econ. Entomol. 1997, 90, 663–668. [Google Scholar] [CrossRef]
- Velasco, L.R.I.; Walter, G.H. Availability of different host plant species and changing abundance of the polyphagous bug Nezara viridula (Hemiptera: Pentatomidae). Environ. Entomol. 1992, 21, 751–759. [Google Scholar] [CrossRef]
- Pasek, J.E. Influence of wind and windbreaks on local dispersal of insects. In Windbreak Technology; Brandle, J.M.R., Hintz, D.L., Sturrock, J.W., Eds.; Elsevier Science: New York, NY, USA, 1998. [Google Scholar]
- Harrington, B.J. A generic level revision and cladistic analysis of the Myodochini of the world (Hemiptera, Lygaeidae, Rhyparochrominae). Bull. Am. Mus. Nat. Hist. 1980, 167, 45–116. [Google Scholar]
- Sweet, M.H. The tribal classification of the Rhyparochrominae (Heteroptera: Lygaeidae). Ann. Entomol. Soc. Am. 1967, 60, 208–226. [Google Scholar] [CrossRef]
- Rodriguez, R.L.S. On the fore legs of seed bugs (Heteroptera: Lygaeidae): Aggression and allometric scaling in Scolopostethus affinis Schilling. J. Kans. Entomol. Soc. 2000, 73, 6–10. [Google Scholar]
- Kuhn, T.M. Bioecology, Characterization of Injuries and Effect of Insecticides on Neopamera sp. aff. bilobata (Say 1832) (Hemiptera: Rhyparochromidae) in Strawberry. Master’s Thesis, Federal University of Pelotas, Pelotas, Brazil, 2014. [Google Scholar]
- Wilson, J.W. Notes on Pamera populations on various types of plant communities in the vicinity of Plant City. Fla. Entomol. 1938, 21, 2830. [Google Scholar] [CrossRef]
- Kennedy, G.G.; Storer, N.P. Life systems of polyphagous arthropod pests in temporally unstable cropping systems. Annu. Rev. Entomol. 2000, 45, 467–493. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Cornell, H.V.; Hochberg, M.E. Predators, parasitoids, and pathogens as mortality agents in phytophagous insect populations. Ecology 1997, 78, 2145–2152. [Google Scholar] [CrossRef]
- Drake, V.A. The influence of weather and climate on agriculturally important insects: An Australian perspective. Aust. J. Agric. Res. 1994, 45, 487–509. [Google Scholar] [CrossRef]
- Solbreck, C.; Sillen-Tullberg, B. Population-dynamics of a seed feeding bug, Lygaeus equestris. Temporal dynamics. Oikos 1990, 58, 210–218. [Google Scholar] [CrossRef]
- Norhisham, A.R.; Abood, F.; Rita, M.; Hakeem, K.R. Effect of humidity on egg hatchability and reproductive biology of the bamboo borer (Dinoderus minutus Fabricius). SpringerPlus 2013, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Blakley, N. Life history significance of size-triggered metamorphosis in milkweed bugs (Oncopeltus). Ecology 1981, 62, 57–64. [Google Scholar] [CrossRef]
- Burdfield-Steel, E.R.; Shuker, D.M. The evolutionary ecology of the Lygaeidae. Ecol. Evol. 2014, 4, 2278–2301. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B.R.; Zandstra, J.; Taghavi, T.; Dale, A. Effects of runner removal on productivity and plant growth of two day-neutral strawberry cultivars in Ontario, Canada. Acta Hortic. 2017, 1156, 327–332. [Google Scholar] [CrossRef]
- Albregts, E.E.; Howard, C.M. Effect of runner removal on strawberry fruiting response. HortScience 1986, 21, 97–98. [Google Scholar]
- Brewer, M.J.; Anderson, D.J.; Armstrong, J.S. Plant Growth Stage-Specific Injury and Economic Injury Level for Verde Plant Bug, Creontiades signatus (Hemiptera: Miridae), on Cotton: Effect of Bloom Period of Infestation. J. Econ. Entomol. 2013, 106, 2077–2083. [Google Scholar] [CrossRef]
- Russin, J.S.; Layton, M.B.; Orr, D.B.; Boethel, D.J. Within-plant Distribution of, and Partial Compensation for, Stink Bug (Heteroptera: Pentatomidae) Damage to Soybean Seeds. J. Econ. Entomol. 1987, 80, 215–220. [Google Scholar] [CrossRef]
- Butts, R.A.; Lamb, R.J. Pest Status of Lygus Bugs (Hemiptera: Miridae) in Oilseed Brassica Crops. J. Econ. Entomol. 1991, 84, 1591–1596. [Google Scholar] [CrossRef]
- Fait, A.; Hanhineva, K.; Beleggia, R.; Dai, N.; Rogachev, I.; Nikiforova, V.J.; Fernie, A.R.; Aharoni, A. Reconfiguration of the Achene and Receptacle Metabolic Networks during Strawberry Fruit Development. Plant Physiol. 2008, 148, 730–750. [Google Scholar] [CrossRef]
- Panizzi, A.R.; Machado-Neto, E. Development of nymphs and feeding habits of nymphal and adult Edessa meditabunda (Heteroptera: Pentatomidae) on soybean and sunflower. Ann. Entomol. Soc. Am. 1992, 85, 477–481. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).