Detection of the Lychee Erinose Mite, Aceria litchii (Keifer) (Acari: Eriophyidae) in Florida, USA: A Comparison with Other Alien Populations


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
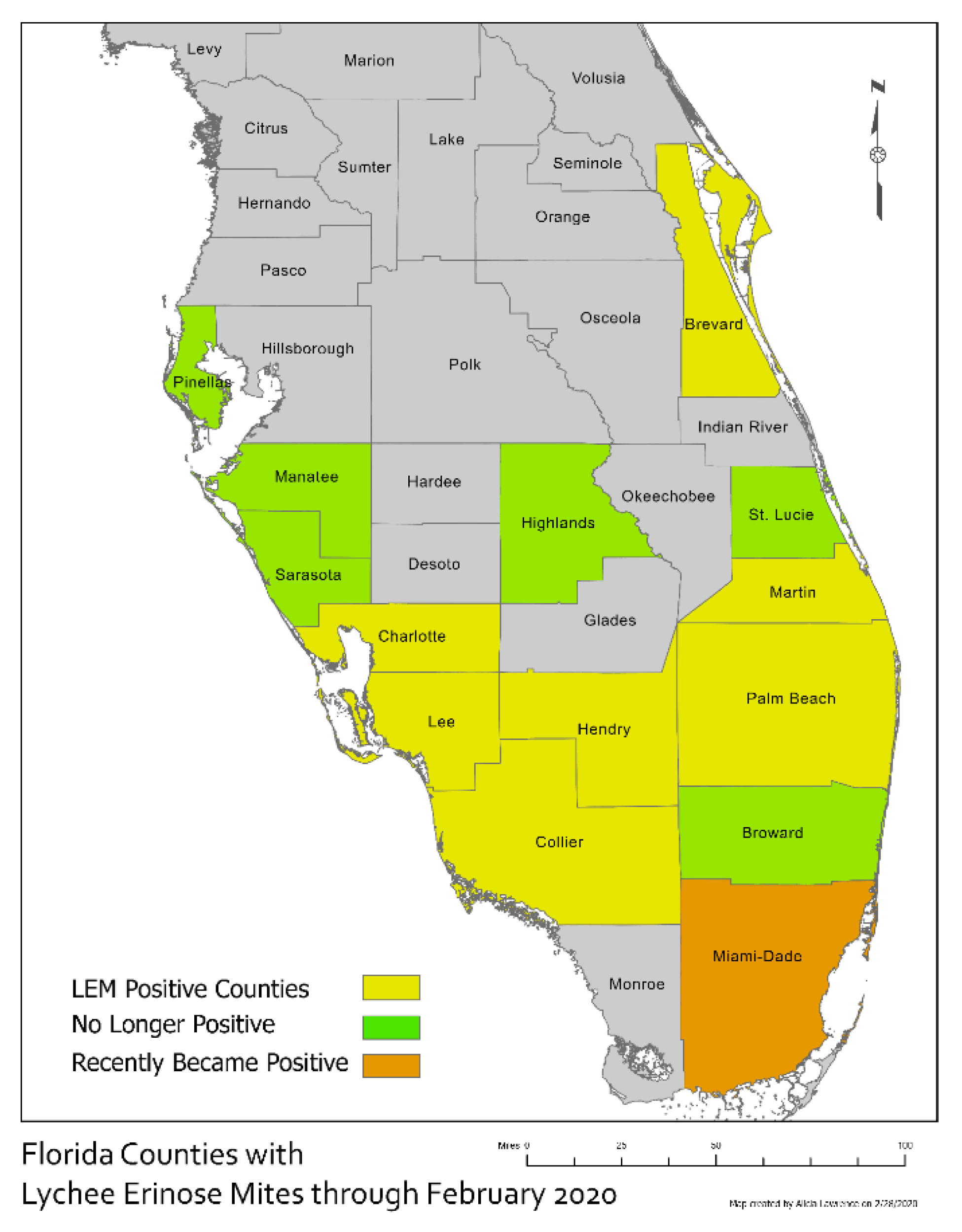
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharma, D.D. Major pests of litchi in Bihar. Indian Farming 1985, 35, 25–26. [Google Scholar]
- Alam, Z.M.; Wadud, M.A. On the biology of Litchi mite, Aceria litchii Keifer (Eriophyidae: Acarina) in East Pakistan. Pak. J. Sci. 1963, 15, 232–240. [Google Scholar]
- Haque, M.M.; Das, B.C.; Khalequzzaman, M.; Chakrabarti, S. Eriophyoid mites (Acari: Eriophyoidea) from Bangladesh. Orient. Insects 1998, 32, 35–40. [Google Scholar] [CrossRef]
- Keifer, H.H.; Knorr, L.C. Eriophyid Mites of Thailand. Plant Prot. Serv. Tech. Bull. (Bangkok) 1978, 38, 1–36. [Google Scholar]
- Huang, T.A. Study on Morphological Features of Erinose Mite of Litchi (Eriophyes litchii Keifer) and an observation on the conditions of its damage. Plant Prot. Bull. 1967, 9, 35–46. [Google Scholar]
- Keifer, H.H. Eriophyid studies XIII. State Calif. Dep. Agric. Bull. 1943, 32, 212–222. [Google Scholar] [CrossRef]
- Pinese, B. Erinose mite—A serious litchi pest. Queensl. Agric. J. 1981, 107, 79–81. [Google Scholar]
- Raga, A.; Mineiro, J.L.D.C.; Sato, M.E.; De Moraes, G.J.; Flechtmann, C.H.W. Primeiro relato de Aceria litchii (Keifer) (prostigmata: Eriophyidae) em plantas de lichia no Brasil. Rev. Bras. Frutic. 2010, 32, 628–629. [Google Scholar] [CrossRef] [Green Version]
- Navia, D.; Júnior, A.L.M.; Gondim, M.G.C., Jr.; de Mendonza, R.s.; da Silva Pereira, P.R.V. Recent mite invasions in South America. In Potential Invasive Pests Agric Crop, 1st ed.; Peña, J.E., Ed.; CABI: Oxfordshire, UK, 2013; pp. 251–287. [Google Scholar]
- Prasad, V.G.; Singh, R.K. Prevalence and control of litchi mite, Aceria litchii (Keifer) in Bihar. Indian J. Entomol. 1981, 43, 67–75. [Google Scholar]
- The USDA 2012 Prioritized Offshore Pest List. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/pest_detection/downloads/farmbill/PrioritizedOffshorePestList.pdf (accessed on 18 June 2019).
- Evans, E.; Degner, R.; Morgan, K. Six Ways to Improve the Profitability of Lychee in South Florida. 2004. EDIS 18. Available online: https://journals.flvc.org/edis/article/view/114112 (accessed on 8 April 2020).
- FDACS (2020) Lychee Erinose Mite Quarantine Information. Available online: https://www.fdacs.gov/Agriculture-Industry/Pests-and-Diseases/Plant-Pests-and-Diseases/Lychee-Erinose-Mite (accessed on 18 March 2020).
- Karioti, A.; Tooulakou, G.; Bilia, A.R.; Psaras, G.K.; Karabourniotis, G.; Skaltsa, H. Erinea formation on Quercus ilex leaves: Anatomical, physiological and chemical responses of leaf trichomes against mite attack. Phytochemistry 2011, 72, 230–237. [Google Scholar] [CrossRef]
- Azevedo, L.H.; Moraes, G.J.; Yamamoto, P.T.; Zanardi, O.Z. Development of a Methodology and Evaluation of Pesticides Against Aceria litchii and Its Predator Phytoseius intermedius (Acari: Eriphyidae, Phytoseiidae). J. Econ. Entomol. 2013, 106, 2183–2189. [Google Scholar] [CrossRef] [PubMed]
- Waite, G.K.; McAlpine, J.D. Honey bees as carriers of lychee erinose mite Eriophyes litchii (Acari: Eriophyiidae). Exp. Appl. Acarol. 1992, 15, 299–302. [Google Scholar] [CrossRef]
- Waite, G.K. New evidence further incriminates honey bees as vectors of lychee erinose mite Aceria litchii (Acari: Eriophyiidae). Exp. Appl. Acarol. 1999, 23, 145–147. [Google Scholar] [CrossRef]
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975; p. 360. [Google Scholar]
- Oldfield, G.N. Diversity and host plant specificity. In Eriophyoid Mites—Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests vol 6; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; pp. 199–216. [Google Scholar]
- Crane, J.H.; Balerdi, C.F.; Maguire, I. Lychee Growing in the Florida Home Landscape. EDIS, 2016(HS6). Available online: https://edis.ifas.ufl.edu/pdffiles/MG/MG05100.pdf (accessed on 8 April 2020).
- Huang, K. Aceria (Acarina: Eriophyoidea) in Taiwan: Five new species and plant abnormalities caused by sixteen species. Zootaxa 2008, 1829, 1–30. [Google Scholar] [CrossRef]
- Bolton, S.J.; Klompen, H.; Bauchan, G.R.; Ochoa, R. A new genus and species of Nematalycidae (Acari: Endeostigmata). J. Nat. Hist. 2014, 48, 1359–1373. [Google Scholar] [CrossRef]
- Carew, M.E.; Goodisman, M.A.D.; Hoffmann, A.A. Species status and population genetic structure of grapevine eriophyoid mites. Entomol. Exp. Appl. 2004, 111, 87–96. [Google Scholar] [CrossRef]
- Navajas, M.; Gutierrez, M.; Lagnel, J.; Boursot, P. Mitochondrial cytochrome oxidase 1 in tetranychid mites: A comparison between molecular phylogeny and changes of morphological and life history traits. Bull. Ent. Res. 1996, 86, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Fenton, B.; Malloch, G.; Moxey, E. Analysis of eriophyid mite rDNA internal transcribed spacer sequences reveals multiple simple sequence repeats. Insect. Mol. Biol. 1997, 6, 23–32. [Google Scholar] [CrossRef]
- Sonnenberg, R.; Nolte, A.W.; Tautz, D. An evaluation of LSU rDNA D1-D2 sequences for their use in species identification. Front. Zool. 2007, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Chetverikov, P.E.; Cvrković, T.; Makunin, A.; Sukhareva, S.; Vidović, B.; Petanović, R. Basal divergence of Eriophyoidea (Acariformes, Eupodina) inferred from combined partial COI and 28S gene sequences and CLSM genital anatomy. Exp. Appl. Acarol. 2015, 67, 219–245. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Bonfield, J.K.; Smith, K.F.; Staden, R. A new DNA sequence assembly program. Nucleic Acids Res. 1995, 23, 4992–4999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2017, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating the human-ape split by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Rach, J.; Bergmann, T.; Paknia, O.; DeSalle, R.; Schierwater, B.; Hadrys, H. The marker choice: Unexpected resolving power of an unexplored CO1 region for layered DNA barcoding approaches. PLoS ONE 2017, 12, e017484. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Subhadrabandhu, S.; Mitra, S.K.; Ben-Aire, R.; Stern, R.A. Origin, History, Production and Processing. In Litchi and Longan: Botany, Production and Uses; Menzel, C.M., Waite, G.K., Eds.; CABI Publishing: Oxfordshire, UK, 2005; pp. 1–25. [Google Scholar]
- Dekle, G.W. Twenty-third biennial Report for the period July 1, 1958-June 30, 1960. State Plant Board Fla. Bull. 1960, 14, 65–70. [Google Scholar]
- Mead, F.A. Entomology Section. Triy-ology 1993, 32, 3. [Google Scholar]
- Waite, G.K. Pests. In Litchi and Longan: Botany, Production and Uses; Menzel, C.M., Waite, G.K., Eds.; CABI Publishing: Oxfordshire, UK, 2005; pp. 237–259. [Google Scholar]
- Castro, B.M.C.; Plata-Rueda, A.; Meloni-Silva, W.; de Menezes, C.W.G.; Wilcke, C.F.; Zanuncio, J.C. Manejo del ácaro Aceria litchii (Acari: Eriophyidae) en Litchi chinensis. Rev. Colomb. Entomol. 2018, 44, 2. [Google Scholar] [CrossRef]
- Schulte, M.J.; Martin, K.; Sauerborn, J. Efficacy of spiromesifen on Aceria litchii (Keifer) in relation to Cephateuros virescens Kunze colonization on leaves of litchi (Litchi chinensis Sonn.). J. Plant. Dis. Prot. 2007, 114, 133–137. [Google Scholar] [CrossRef]
- Picoli, P.R.F.; Vieira, M.R.; da Silva, E.A.; da Mota, M.S.d.O. Ácaros Predadores Associados Ao Ácaro-Da-Erinose Da Lichia. Pesqui Agropecu Bras 2010, 45, 1246–1252. [Google Scholar] [CrossRef] [Green Version]
- Nishida, T.; Holdaway, F.G. The Erinose Mite of Lychee. Circular No. 48; Hawaii Agriculture Experiment Station: Honolulu, HI, USA, 1955; p. 10. [Google Scholar]
- Revynthi, A.M.; Duncan, R.E.; Mannion, C.; Kendra, P.E.; Carrillo, D. Post-harvest paraffinic oil dips to disinfest lychee fruit from lychee erinose mite. Fla. Entomol. 2020, (in press). [Google Scholar]
- Lall, B.S.; Rahman, M.F. Studies on the bionomics and control of the erinose mite Eriophyes litchii Keifer (Acarina: Eriophyidae). Pesticides 1975, 9, 49–54. [Google Scholar]
- Schicha, E. Phytoseiidae of Australia and Neighbouring Areas; Indira Publishing House: West Bloomfield, MI, USA, 1987; 187p. [Google Scholar]
- Waite, G.K.; Gerson, U. The predator guild associated with Aceria litchii (Acari: Eriophyidae) in Australia and China. Entomophaga 1994, 39, 275–280. [Google Scholar] [CrossRef]
- Azevedo, L.H.; Maeda, E.Y.; Inomoto, M.M.; De Moraes, G.J. A Method to Estimate the Population Level of Aceria litchii (Prostigmata: Eriophyidae) and a Study of the Population Dynamics of This Species and Its Predators on Litchi Trees in Southern Brazil. J. Econ. Entomol. 2014, 107, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Waite, G.K.; Hwang, J.S. Pests of litchi and longan. In Tropical Fruit Pests and Pollinators: Biology Economic Importance, Natural Enemies and Control; Peña, J.E., Sharp, J.L., Wysoki, M., Eds.; CAB International: Wallingford, UK, 2002; pp. 331–359. [Google Scholar]
- Cheng, L.; Zhang, X.; Sha, L.; Lu, A.; Chen, P. Functional and numerical response of Amblyseius largoensis to Aceria litchii. J. Trop. Crop. 2005, 26, 53. [Google Scholar]
- Picoli, P.R.F.; Vieira, M.R. Primeiro relato de atividade patogênica de Hirsutella thompsonii (Fischer) sobre o ácaro-da-erinose-da-lichia Aceria litchii (Keifer). Semin. Agrar. 2013, 34, 187–190. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographic Origin | Coordinates | Collector | Collection Date | GenBank Sequences | ||
|---|---|---|---|---|---|---|
| COI | ITS1-5.8 | ITS2-28S 5’ | ||||
| Australia | 24.87 S 56.78 E | E. Dunn & J.J. Beard | 3-Apr-19 | MT027817 | MN594262 | MN594268 |
| Brazil | 22.71 S 43.63 W | G.J de Moraes | 25-Mar-19 | MT027818 | MN594263 | MN594269 |
| Florida | 26.6 N 82.1 W | D. Carrillo & A.M Revynthi | 20-Feb-18 | MT027819 | MN594264 | MN594270 |
| Hawaii | 21.30 N 157.82 W | B. Azama & M. Ramadan | 15-Mar-18 | MT027820 | MN594265 | MN594271 |
| India | 22.94 N 88.53 E | K. Karmakar | 25-Mar-19 | MT027821 | MN594266 | MN594272 |
| Taiwan | 22.64 N 120.35 E | C.F. Hong | 22-Apr-19 | MT027822 | MN594267 | MN594273 |
| Molecular Marker | Number of Sites | No. of Alleles/Haplotypes | No. of Segregating Sites | Nucleotide Diversity Per Site (π) |
|---|---|---|---|---|
| ITS1 | 492 | 2 | 62 | 0.04665 |
| ITS2 | 936 | 4 | 64 | 0.03874 |
| COI | 363 | 4 | 6 | 0.00724 |
| ITS1 | ITS2 | COI | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FL | Au | Br | HW | Ind | FL | Au | Br | HW | Ind | FL | Au | Br | HW | Ind | |
| Au | 0.140 | 0.066 | 0.008 | ||||||||||||
| Br | 0.000 | 0.140 | 0.004 | 0.069 | 0.008 | 0.000 | |||||||||
| HW | 0.000 | 0.140 | 0.000 | 0.066 | 0.000 | 0.069 | 0.008 | 0.000 | 0.000 | ||||||
| Ind | 0.000 | 0.140 | 0.000 | 0.000 | 0.009 | 0.067 | 0.009 | 0.067 | 0.017 | 0.008 | 0.008 | 0.008 | |||
| Tw | 0.000 | 0.140 | 0.000 | 0.000 | 0.000 | 0.000 | 0.066 | 0.004 | 0.066 | 0.009 | 0.017 | 0.008 | 0.008 | 0.008 | 0.000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrillo, D.; Cruz, L.F.; Revynthi, A.M.; Duncan, R.E.; Bauchan, G.R.; Ochoa, R.; Kendra, P.E.; Bolton, S.J. Detection of the Lychee Erinose Mite, Aceria litchii (Keifer) (Acari: Eriophyidae) in Florida, USA: A Comparison with Other Alien Populations. Insects 2020, 11, 235. https://doi.org/10.3390/insects11040235
Carrillo D, Cruz LF, Revynthi AM, Duncan RE, Bauchan GR, Ochoa R, Kendra PE, Bolton SJ. Detection of the Lychee Erinose Mite, Aceria litchii (Keifer) (Acari: Eriophyidae) in Florida, USA: A Comparison with Other Alien Populations. Insects. 2020; 11(4):235. https://doi.org/10.3390/insects11040235
Chicago/Turabian StyleCarrillo, Daniel, Luisa F. Cruz, Alexandra M. Revynthi, Rita E. Duncan, Gary R. Bauchan, Ronald Ochoa, Paul E. Kendra, and Samuel J. Bolton. 2020. "Detection of the Lychee Erinose Mite, Aceria litchii (Keifer) (Acari: Eriophyidae) in Florida, USA: A Comparison with Other Alien Populations" Insects 11, no. 4: 235. https://doi.org/10.3390/insects11040235
APA StyleCarrillo, D., Cruz, L. F., Revynthi, A. M., Duncan, R. E., Bauchan, G. R., Ochoa, R., Kendra, P. E., & Bolton, S. J. (2020). Detection of the Lychee Erinose Mite, Aceria litchii (Keifer) (Acari: Eriophyidae) in Florida, USA: A Comparison with Other Alien Populations. Insects, 11(4), 235. https://doi.org/10.3390/insects11040235