Induced Systemic Resistance by a Plant Growth-Promoting Rhizobacterium Impacts Development and Feeding Behavior of Aphids

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants, Insects and Rhizobacteria
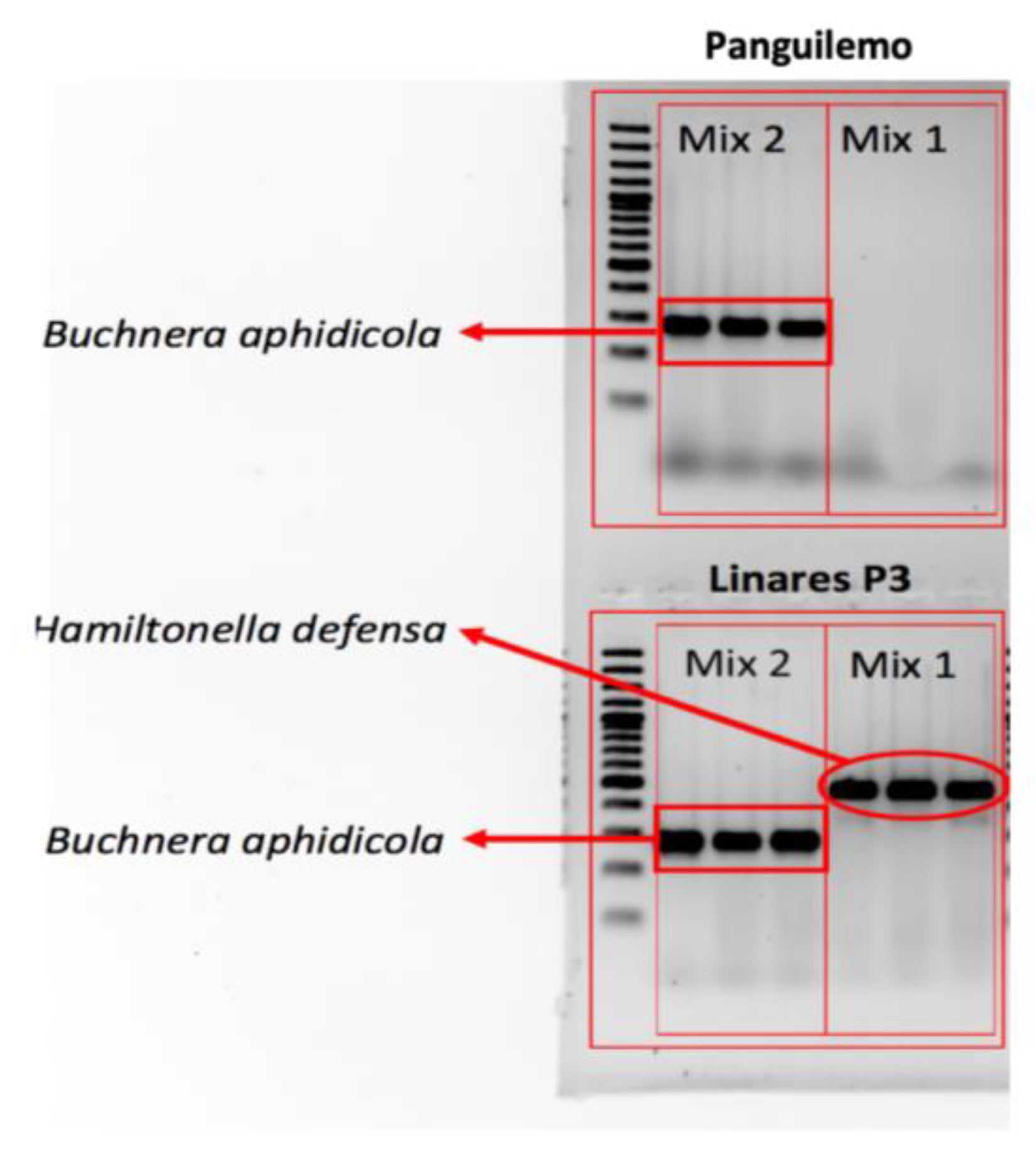
2.2. Aphid Genotype and Endosymbiont Screening
2.3. Aphid Population Growth According to PGPR Inoculation
2.4. Feeding Behavior of Aphids
2.5. Statistical Analyses
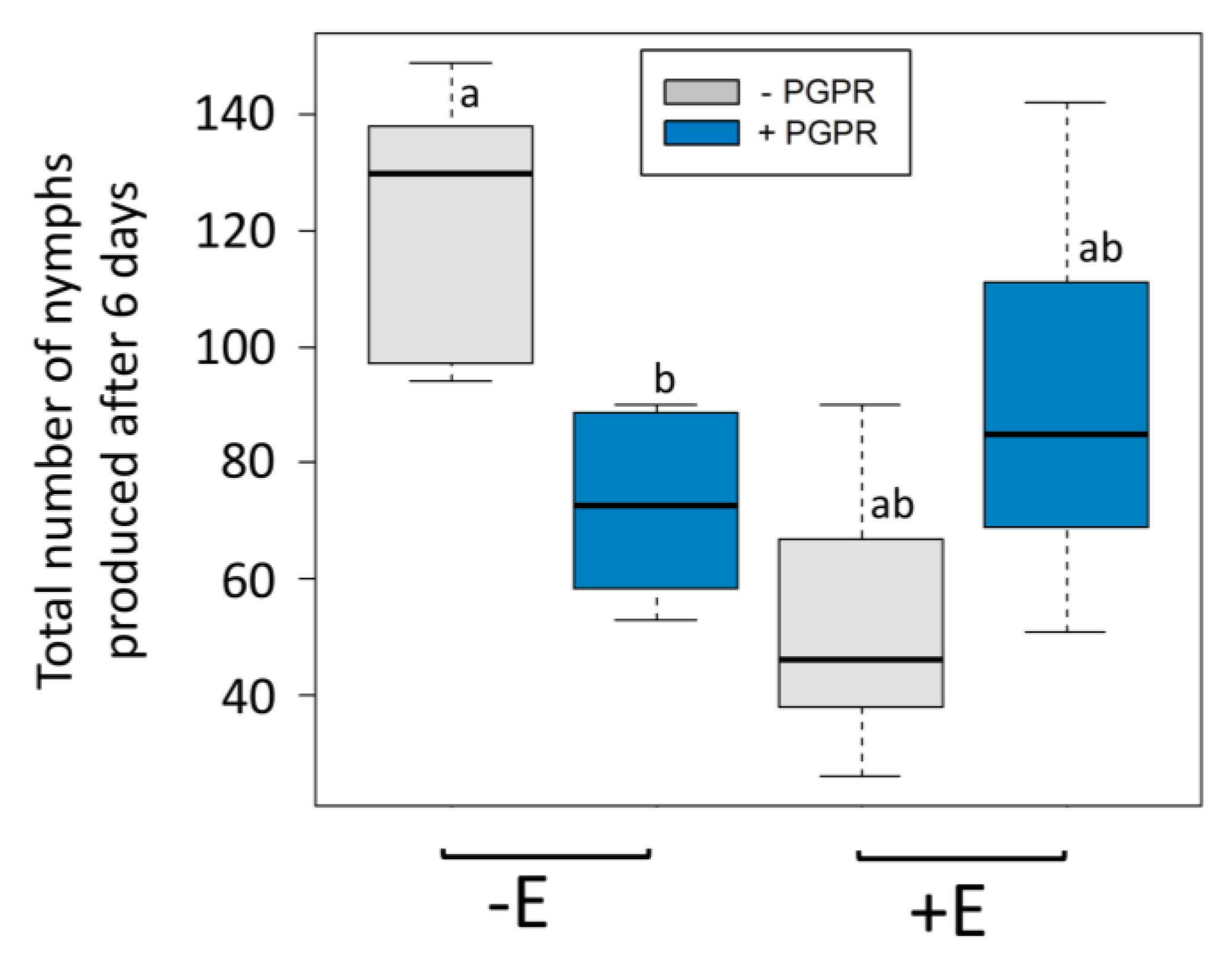
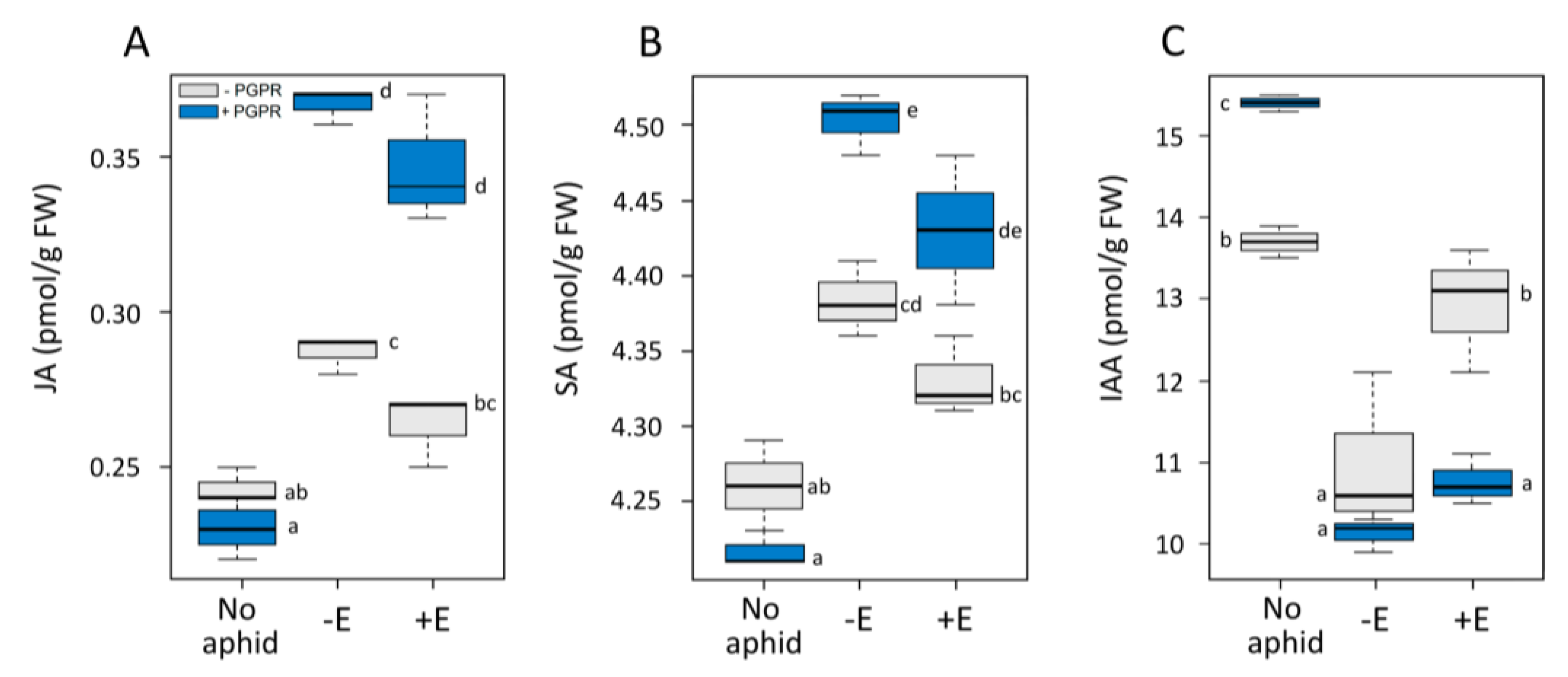
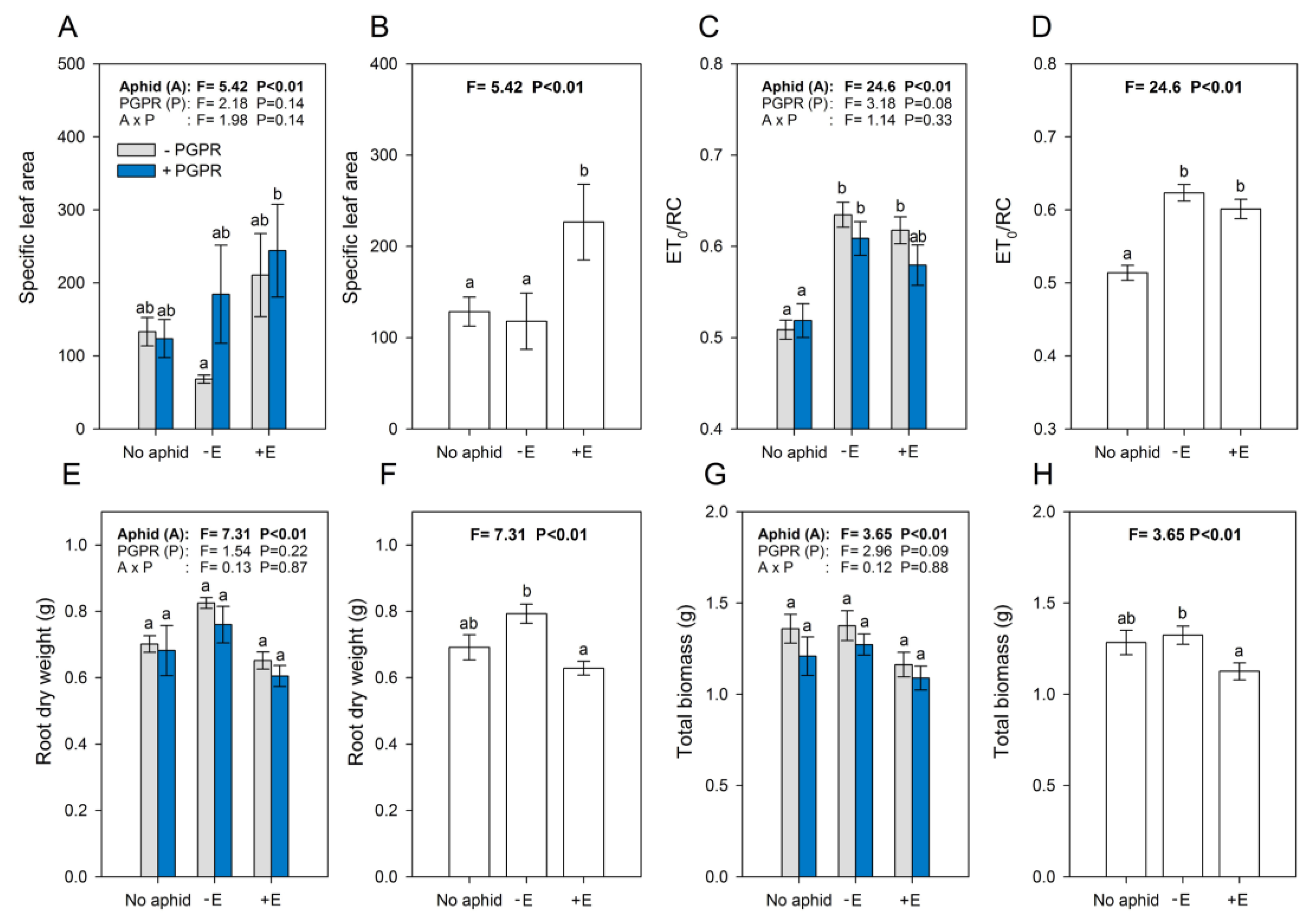
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Hartmann, A.; Rothballer, M.; Schmid, M. Lorenz Hiltner, a pioneer in rhizosphere microbial ecology and soil bacteriology research. Plant Soil 2008, 312, 7–14. [Google Scholar] [CrossRef]
- Dardanelli, M.S.; Medeot, D.B.; Paulucci, N.S.; Bueno, M.A.; Vicario, J.C.; García, M.; Bensi, N.H.; Niebylski, A.M. Biochemical Processes of Rhizobacteria and Their Application in Biotechnology; Malik, A., Grohmann, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; ISBN 9789400715912. [Google Scholar]
- Kloepper, J.W.; Rodríguez-Kábana, R.; Zehnder, G.W.; Murphy, J.F.; Sikora, E.; Fernández, C. Plant root-bacterial interactions in biological control of soilborne diseases and potential extension to systemic and foliar diseases. Australas. Plant Pathol. 1999, 28, 21–26. [Google Scholar] [CrossRef]
- Ramamoorthy, V.; Viswanathan, R.; Raguchander, T.; Prakasam, V.; Samiyappan, R. Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop Prot. 2001, 20, 1–11. [Google Scholar] [CrossRef]
- Walters, D.; Heil, M. Costs and trade-offs associated with induced resistance. Physiol. Mol. Plant Pathol. 2007, 71, 3–17. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Fan, B.; Blom, J.; Klenk, H.P.; Borriss, R. Bacillus amyloliquefaciens, Bacillus velezensis, and Bacillus siamensis form an “Operational group B. amyloliquefaciens” within the B. subtilis species complex. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Matilla, M.; Krell, T. Plant growth promotion and biocontrol mediated by plant-associated bacteria BT. In Plant Microbiome: Stress Response; Egamberdieva, D., Ahmad, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 45–80. [Google Scholar]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.W.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42-A review. Front. Microbiol. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zehnder, G.; Kloepper, J.; Tuzun, S.; Yao, C.; Wei, G.; Chambliss, O.; Shelby, R. Insect feeding on cucumber mediated by rhizobacteria-induced plant resistance. Entomol. Exp. Appl. 1997, 83, 81–85. [Google Scholar] [CrossRef]
- Van der Ent, S.; Van Wees, S.C.M.; Pieterse, C.M.J. Jasmonate signaling in plant interactions with resistance-inducing beneficial microbes. Phytochemistry 2009, 70, 1581–1588. [Google Scholar] [CrossRef] [Green Version]
- Song, G.C.; Ryu, C.M. Two volatile organic compounds trigger plant self-defense against a bacterial pathogen and a sucking insect in cucumber under open field conditions. Int. J. Mol. Sci. 2013, 14, 9803–9819. [Google Scholar] [CrossRef] [Green Version]
- Conrath, U.; Pieterse, C.M.J.; Mauch-mani, B. Priming in plant-pathogen interactions. Trends Plant Sci. 2002, 7, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and plants-With special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef] [PubMed]
- Van Peer, R.; Niemann, G.J.; Schippers, B. Induced resistance and phytoalexin accumulation in biological control of Fusarium Wilt of Carnation by Pseudomonas sp. Strain WCS417r. Phytopathology 1991, 81, 728–734. [Google Scholar] [CrossRef]
- Pastor, V.; Luna, E.; Mauch-Mani, B.; Ton, J.; Flors, V. Primed plants do not forget. Environ. Exp. Bot. 2012, 94, 46–56. [Google Scholar] [CrossRef]
- Fahimi, A.; Ashouri, A.; Ahmadzadeh, M.; Hoseini Naveh, V.; Asgharzadeh, A.; Maleki, F.; Felton, G.W. Effect of PGPR on population growth parameters of cotton aphid. Arch. Phytopathol. Plant Prot. 2014, 47, 1274–1285. [Google Scholar] [CrossRef]
- Disi, J.O.; Zebelo, S.; Kloepper, J.W.; Fadamiro, H. Seed inoculation with beneficial rhizobacteria affects European corn borer (Lepidoptera: Pyralidae) oviposition on maize plants. Entomol. Sci. 2018, 21, 48–58. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Ammar, E.-D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [Green Version]
- Peccoud, J.; Figueroa, C.C.; Silva, A.X.; Ramirez, C.C.; Mieuzet, L.; Bonhomme, J.; Stoeckel, S.; Plantagenest, M.; Simon, J.-C. Host range expansion of an introduced insect pest through multiple colonizations of specialized clones. Mol. Ecol. 2008, 17, 4608–4618. [Google Scholar] [CrossRef]
- Guyomar, C.; Legeai, F.; Jousselin, E.; Mougel, C.; Lemaitre, C.; Simon, J.C. Multi-scale characterization of symbiont diversity in the pea aphid complex through metagenomic approaches. Microbiome 2018, 6, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Hatt, S.; He, K.; Chen, J.; Francis, F.; Wang, Z. Nine facultative endosymbionts in aphids. A review. J. Asia Pac. Entomol. 2017, 20, 794–801. [Google Scholar] [CrossRef]
- Su, Q.; Oliver, K.M.; Xie, W.; Wu, Q.; Wang, S.; Zhang, Y. The whitefly-associated facultative symbiont Hamiltonella defensa suppresses induced plant defences in tomato. Funct. Ecol. 2015, 29, 1007–1018. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degnan, P.H.; Hunter, M.S.; Moran, N.A. Bacteriophages encode factors required for protection in a symbiotic mutualism. Science (80-) 2009, 325, 992–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frago, E.; Mala, M.; Weldegergis, B.T.; Yang, C.; McLean, A.; Godfray, H.C.J.; Gols, R.; Dicke, M. Symbionts protect aphids from parasitic wasps by attenuating herbivore-induced plant volatiles. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelella, G.; Nalam, V.; Nachappa, P.; White, J.; Kaplan, I. Endosymbionts differentially alter exploratory probing behavior of a nonpersistent plant virus vector. Microb. Ecol. 2018, 76, 453–458. [Google Scholar] [CrossRef]
- Hackett, S.C.; Karley, A.J.; Bennett, A.E. Unpredicted impacts of insect endosymbionts on interactions between soil organisms, plants and aphids. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131275. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Kim, S.J.; Kwon, S.W.; Rooney, A.P. Bacillus velezensis is not a later heterotypic synonym of Bacillus amyloliquefaciens; Bacillus methylotrophicus, Bacillus amyloliquefaciens subsp. Plantarum and ‘Bacillus oryzicola’ are later heterotypic synonyms of Bacillus velezensis based on phylogenom. Int. J. Syst. Evol. Microbiol. 2016, 66, 1212–1217. [Google Scholar] [CrossRef]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef]
- Borodovsky, M.; McIninch, J. GenMark: Parallel gene recognition for both DNA strands. Comput. Chem. 1993, 17, 123–133. [Google Scholar] [CrossRef]
- Peccoud, J.; Bonhomme, J.; Mahéo, F.; de la Huerta, M.; Cosson, O.; Simon, J.C. Inheritance patterns of secondary symbionts during sexual reproduction of pea aphid biotypes. Insect Sci. 2014, 21, 291–300. [Google Scholar] [CrossRef]
- Ramos, P.; Rivas, N.; Pollmann, S.; Casati, P.; Molina-Montenegro, M.A. Hormonal and physiological changes driven by fungal endophytes increase Antarctic plant performance under UV-B radiation. Fungal Ecol. 2018, 34, 76–82. [Google Scholar] [CrossRef]
- Carrasco Loba, V.; Pérez Alonso, M.-M.; Pollmann, S. Monitoring of crosstalk between jasmonate and auxin in the framework of plant stress responses of roots. In Auxins and Cytokinins in Plant Biology: Methods and Protocols; Dandekar, T., Naseem, M., Eds.; Humana Press: New York, NY, USA, 2017; pp. 175–185. ISBN 978-1-4939-6829-9. [Google Scholar]
- Manschadi, A.M.; Sauerborn, J.; Stützel, H.; Göbel, W.; Saxena, M.C. Simulation of faba bean (Vicia faba L.) root system development under Mediterranean conditions. Eur. J. Agron. 1998, 9, 259–272. [Google Scholar] [CrossRef]
- Adasme-Carreño, F.; Muñoz-Gutiérrez, C.; Salinas-Cornejo, J.; Ramírez, C.C. A2EPG: A new software for the analysis of electrical penetration graphs to study plant probing behaviour of hemipteran insects. Comput. Electron. Agric. 2015, 113, 128–135. [Google Scholar] [CrossRef]
- Sarria, E.; Cid, M.; Garzo, E.; Fereres, A. Excel Workbook for automatic parameter calculation of EPG data. Comput. Electron. Agric. 2009, 67, 35–42. [Google Scholar] [CrossRef]
- Buensanteai, N.; Yuen, G.Y.; Prathuangwong, S. Priming, signaling, and protein production associated with induced resistance by Bacillus amyloliquefaciens KPS46. World J. Microbiol. Biotechnol. 2009, 25, 1275–1286. [Google Scholar] [CrossRef]
- Beris, D.; Theologidis, I.; Skandalis, N.; Vassilakos, N. Bacillus amyloliquefaciens strain MBI600 induces salicylic acid dependent resistance in tomato plants against Tomato spotted wilt virus and Potato virus y. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asari, S.; Ongena, M.; Debois, D.; De Pauw, E.; Chen, K.; Bejai, S.; Meijer, J. Insights into the molecular basis of biocontrol of Brassica pathogens by Bacillus amyloliquefaciens UCMB5113 lipopeptides. Ann. Bot. 2017, 120, 551–562. [Google Scholar] [CrossRef]
- Brock, A.K.; Berger, B.; Schreiner, M.; Ruppel, S.; Mewis, I. Plant growth-promoting bacteria Kosakonia radicincitans mediate anti-herbivore defense in Arabidopsis thaliana. Planta 2018, 248, 1383–1392. [Google Scholar] [CrossRef]
- Xie, S.; Jiang, H.; Ding, T.; Xu, Q.; Chai, W.; Cheng, B. Bacillus amyloliquefaciens FZB42 represses plant miR846 to induce systemic resistance via a jasmonic acid-dependent signalling pathway. Mol. Plant Pathol. 2018, 19, 1612–1623. [Google Scholar] [CrossRef] [Green Version]
- Kloth, K.J.; Wiegers, G.L.; Busscher-Lange, J.; Van Haarst, J.C.; Kruijer, W.; Bouwmeester, H.J.; Dicke, M.; Jongsma, M.A. AtWRKY22 promotes susceptibility to aphids and modulates salicylic acid and jasmonic acid signalling. J. Exp. Bot. 2016, 67, 3383–3396. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.B.; Nault, B.A.; Smart, C.D. Effects of plant growth-promoting rhizobacteria on bell pepper production and green peach aphid infestations in New York. Crop Prot. 2008, 27, 996–1002. [Google Scholar] [CrossRef]
- Boutard-Hunt, C.; Smart, C.D.; Thaler, J.; Nault, B.A. Impact of plant growth-promoting rhizobacteria and natural enemies on Myzus persicae (Hemiptera: Aphididae) infestations in pepper. J. Econ. Entomol. 2009, 102, 2183–2191. [Google Scholar] [CrossRef]
- Martinuz, A.; Schouten, A.; Menjivar, R.D.; Sikora, R.A. Effectiveness of systemic resistance toward Aphis gossypii (Hom., Aphididae) as induced by combined applications of the endophytes Fusarium oxysporum Fo162 and Rhizobium etli G12. Biol. Control 2012, 62, 206–212. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Gange, A.C. Plant-associated Bacillus spp. alter life-history traits of the specialist insect Brevicoryne brassicae L. Agric. For. Entomol. 2016, 18, 35–42. [Google Scholar] [CrossRef]
- Naeem, M.; Aslam, Z.; Khaliq, A.; Ahmed, J.N.; Nawaz, A.; Hussain, M. Plant growth promoting rhizobacteria reduce aphid population and enhance the productivity of bread wheat. Braz. J. Microbiol. 2018, 49, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Pineda, A.; Zheng, S.J.; van Loon, J.J.A.; Dicke, M. Rhizobacteria modify plant-aphid interactions: A case of induced systemic susceptibility. Plant Biol. 2012, 14, 83–90. [Google Scholar] [CrossRef]
- Blubaugh, C.K.; Carpenter-Boggs, L.; Reganold, J.P.; Schaeffer, R.N.; Snyder, W.E. Bacteria and competing herbivores weaken top–down and bottom–up aphid suppression. Front. Plant Sci. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Stewart, S.A.; Hodge, S.; Bennett, M.; Mansfield, J.W.; Powell, G. Aphid induction of phytohormones in Medicago truncatula is dependent upon time post-infestation, aphid density and the genotypes of both plant and insect. Arthropod Plant Interact. 2016, 10, 41–53. [Google Scholar] [CrossRef]
- Oliver, K.M.; Higashi, C.H. Variations on a protective theme: Hamiltonella defensa infections in aphids variably impact parasitoid success. Curr. Opin. Insect Sci. 2019, 32, 1–7. [Google Scholar] [CrossRef]
- Sochard, C.; Morlière, S.; Toussaint, G.; Outreman, Y.; Sugio, A.; Simon, J.C. Examination of the success rate of secondary symbiont manipulation by microinjection methods in the pea aphid system. Entomol. Exp. Appl. 2020, 1–10. [Google Scholar] [CrossRef]
- Blakeslee, J.J.; Spatola Rossi, T.; Kriechbaumer, V. Auxin biosynthesis: Spatial regulation and adaptation to stress. J. Exp. Bot. 2019, 70, 5041–5049. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Park, J.M.; Lazarovits, G.; Bernards, M.A. Burkholderia phytofirmans-induced shoot and root growth promotion is associated with endogenous changes in plant growth hormone levels. Plant Growth Regul. 2014, 75, 199–207. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, J.S.; Meena, V.S.; Ramteke, P.W. Plant growth-promoting rhizobacteria: Strategies to improve abiotic stresses under sustainable agriculture. J. Plant Nutr. 2019, 42, 1402–1415. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, J.S.; Meena, V.S.; Srivastava, R. Recent advances of PGPR based approaches for stress tolerance in plants for sustainable agriculture. Biocatal. Agric. Biotechnol. 2019, 20, 101271. [Google Scholar] [CrossRef]
- He, L.; Li, C.; Liu, R. Indirect interactions between arbuscular mycorrhizal fungi and Spodoptera exigua alter photosynthesis and plant endogenous hormones. Mycorrhiza 2017, 27, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Radwan, S.S.; Dashti, N.; El-Nemr, I.M. Enhancing the growth of Vicia faba plants by microbial inoculation to improve their phytoremediation potential for oily desert areas. Int. J. Phytoremed. 2005, 7, 19–32. [Google Scholar] [CrossRef]
- Elbadry, M.; Taha, R.M.; Eldougdoug, K.A.; Gamal−Eldin, H. Induction of systemic resistance in faba bean (Vicia faba L.) to bean yellow mosaic potyvirus (BYMV) via seed bacterization with plant growth promoting rhizobacteria. J. Plant Dis. Prot. 2006, 113, 247–251. [Google Scholar] [CrossRef]
- Nalam, V.; Louis, J.; Shah, J. Plant defense against aphids, the pest extraordinaire. Plant Sci. 2019, 279, 96–107. [Google Scholar] [CrossRef]
- Alvarez, A.E.; Tjallingii, W.F.; Garzo, E.; Vleeshouwers, V.; Dicke, M.; Vosman, B. Location of resistance factors in the leaves of potato and wild tuber-bearing Solanum species to the aphid Myzus persicae. Entomol. Exp. Appl. 2006, 121, 145–157. [Google Scholar] [CrossRef]
- Le Roux, V.; Dugravot, S.; Brunissen, L.; Vincent, C.; Pelletier, Y.; Giordanengo, P. Antixenosis phloem-based resistance to aphids: Is it the rule? Ecol. Entomol. 2010, 35, 407–416. [Google Scholar] [CrossRef]
- Will, T.; Kornemann, S.R.; Furch, A.C.U.; Tjallingii, W.F.; van Bel, A.J.E. Aphid watery saliva counteracts sieve-tube occlusion: A universal phenomenon? J. Exp. Biol. 2009, 212, 3305–3312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paprocka, M.; Gliszczyńska, A.; Dancewicz, K.; Gabryś, B. Novel hydroxy- and epoxy-cis-jasmone and dihydrojasmone derivatives affect the foraging activity of the peach potato aphid Myzus persicae (Sulzer) (Homoptera: Aphididae). Molecules 2018, 23, 2362. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Dai, H.; Zhang, Y.; Chandrasekar, R.; Luo, L.; Hiromasa, Y.; Sheng, C.; Peng, G.; Chen, S.; Tomich, J.M.; et al. Armet is an effector protein mediating aphid-plant interactions. FASEB J. 2015, 29, 2032–2045. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Luo, L.; Lu, H.; Chen, S.; Kang, L.; Cui, F. Angiotensin-converting enzymes modulate aphid–plant interactions. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Will, T.; Tjallingii, W.F.; Thonnessen, A.; van Bel, A.J.E. Molecular sabotage of plant defense by aphid saliva. Proc. Natl. Acad. Sci. USA 2007, 104, 10536–10541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naessens, E.; Dubreuil, G.; Giordanengo, P.; Baron, O.L.; Minet-Kebdani, N.; Keller, H.; Coustau, C. A secreted MIF cytokine enables aphid feeding and represses plant immune responses. Curr. Biol. 2015, 25, 1898–1903. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | Locus | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| ApF08 | ALB04 | ALB08 | ALB12 | ALB07 | ApH10 | Ap03 | ApH08 | ALA12 | |
| +E | 183/189 | 267/269 | 277/290 | 321/345 | 136/152 | 203/213 | 261/261 | 268/284 | 435/459 |
| −E | 181/193 | 266/268 | 294/302 | 329/335 | 135/171 | 215/217 | 254/261 | 266/268 | 449/457 |
| EPG Parameters | Localization of Resistance Factors | −E | +E | ||||
|---|---|---|---|---|---|---|---|
| Surface | Mesophyll | Phloem | −PGPR | +PGPR | −PGPR | +PGPR | |
| Mean duration of NP (min) | X | 6.41 ± 5.95 d | 5.59 ± 9.00 c | 2.97 ± 1.76 a | 4.27 ± 5.21 b | ||
| Duration of the 2nd non-probe period (min) | X | X | 4.31 ± 5.50 d | 2.99 ± 3.74 c | 2.69 ± 3.80 b | 1.88 ± 2.01 a | |
| Number of short probes (C < 3 min) | X | X | 5.59 ± 5.02 a | 5.97 ± 5.33 a | 10.15 ± 10.84 b | 8.15 ± 9.59 b | |
| Time from start of EPG to 1st E2 (min) | X | X | X | 129.37 ± 77.09 d | 123.35 ± 76.93 b | 127.98 ± 86.23 c | 103.35 ± 68.26 a |
| Duration of NP just after the probe of the 1st sustained E2 (min) | X | X | 1.39 ± 3.90 c | 1.82 ± 4.82 d | 0.73 ± 2.12 b | 0.32 ± 0.83 a | |
| Mean duration of F (min) | X | 0.10 ± 0.49 a | 0.65 ± 3.28 b | 4.40 ± 16.31 c | 0.00 ± 0.00ab c | ||
| Time from the beginning of the 1st probe to 1st pd (min) | X | 6.98 ± 18.80 d | 4.80 ± 12.57 c | 0.79 ± 1.75 a | 1.06 ± 2.57 b | ||
| Duration of 1st probe (min) | X | X | 10.90 ± 44.98 b | 33.57 ± 76.30 d | 18.43 ± 63.67 c | 1.17 ± 1.95 a | |
| Time from the beginning of that probe to 1st E (min) | X | 22.09 ± 14.57 b | 31.02 ± 31.60 d | 21.36 ± 17.44 a | 25.13 ± 12.45 c | ||
| Duration of 1st E (min) | X | 71.97 ± 73.17 d | 56.78 ± 61.83 a | 70.14 ± 80.36 c | 60.10 ± 68.13 b | ||
| Time from the beginning of that probe to 1st sustained E2 | X | X | 25.05 ± 17.09 b | 31.39 ± 32.46 d | 22.40 ± 19.16 a | 25.79 ± 13.09 c | |
| Mean duration of E1 (min) | X | 1.14 ± 0.86 a | 1.10 ± 1.58 a | 1.05 ± 1.14 a | 1.38 ± 1.49 b | ||
| Number of E1 | X | 1.22 ± 0.97 a | 1.42 ± 1.06a b | 1.33 ± 1.18 a | 2.31 ± 2.04 b | ||
| Duration of E1 followed by the 1st E2 (min) | X | 1.04 ± 0.72 c | 0.92 ± 0.64 b | 0.75 ± 0.51 a | 0.93 ± 0.52 b | ||
| Duration of E1 followed by the 1st sustained E2 (min) | X | 1.12 ± 0.90 b | 0.86 ± 0.53 a | 0.93 ± 1.09 a | 1.14 ± 1.10 b | ||
| Number of sustained E2 (<10 min) | X | 1.00 ± 0.78 a | 1.03 ± 0.75 a | 0.89 ± 0.75 a | 1.65 ± 1.35 a | ||
| Duration of G (min) | 13.65 ± 30.82 c | 5.98 ± 15.32 a | 18.85 ± 46.92 d | 6.94 ± 19.64 b | |||
| Number of G | 0.30 ± 0.67 a | 0.29 ± 0.78 a | 0.37 ± 0.69 a | 0.12 ± 0.33 a | |||
| % of probing spent in G | 7.44 ± 16.92 b | 3.35 ± 8.32 a | 8.70 ± 21.19 b | 3.31 ± 9.45 a | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serteyn, L.; Quaghebeur, C.; Ongena, M.; Cabrera, N.; Barrera, A.; Molina-Montenegro, M.A.; Francis, F.; Ramírez, C.C. Induced Systemic Resistance by a Plant Growth-Promoting Rhizobacterium Impacts Development and Feeding Behavior of Aphids. Insects 2020, 11, 234. https://doi.org/10.3390/insects11040234
Serteyn L, Quaghebeur C, Ongena M, Cabrera N, Barrera A, Molina-Montenegro MA, Francis F, Ramírez CC. Induced Systemic Resistance by a Plant Growth-Promoting Rhizobacterium Impacts Development and Feeding Behavior of Aphids. Insects. 2020; 11(4):234. https://doi.org/10.3390/insects11040234
Chicago/Turabian StyleSerteyn, Laurent, Céleste Quaghebeur, Marc Ongena, Nuri Cabrera, Andrea Barrera, Marco A. Molina-Montenegro, Frédéric Francis, and Claudio C. Ramírez. 2020. "Induced Systemic Resistance by a Plant Growth-Promoting Rhizobacterium Impacts Development and Feeding Behavior of Aphids" Insects 11, no. 4: 234. https://doi.org/10.3390/insects11040234
APA StyleSerteyn, L., Quaghebeur, C., Ongena, M., Cabrera, N., Barrera, A., Molina-Montenegro, M. A., Francis, F., & Ramírez, C. C. (2020). Induced Systemic Resistance by a Plant Growth-Promoting Rhizobacterium Impacts Development and Feeding Behavior of Aphids. Insects, 11(4), 234. https://doi.org/10.3390/insects11040234