Predatory Earwigs are Attracted by Herbivore-Induced Plant Volatiles Linked with Plant Growth-Promoting Rhizobacteria

 ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture and Plant Treatments
2.2. Insect Rearing
2.2.1. Plutella Xylostella
2.2.2. Doru Luteipes
2.3. Plant Weight Measurements
2.4. Y-Tube Olfactometer Experiments
2.5. Statistical Analyses
3. Results
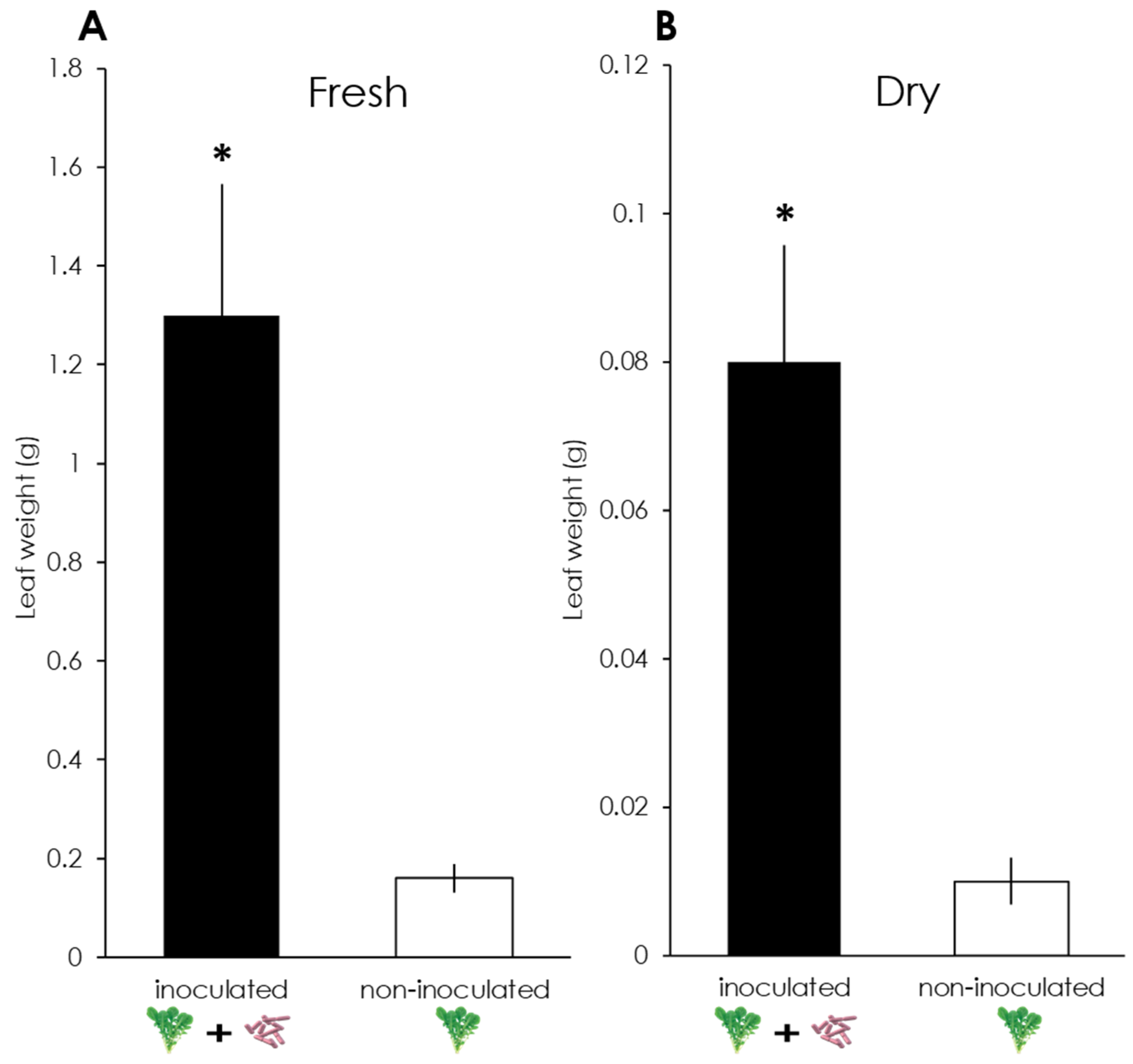
3.1. Plant Weight Measurement
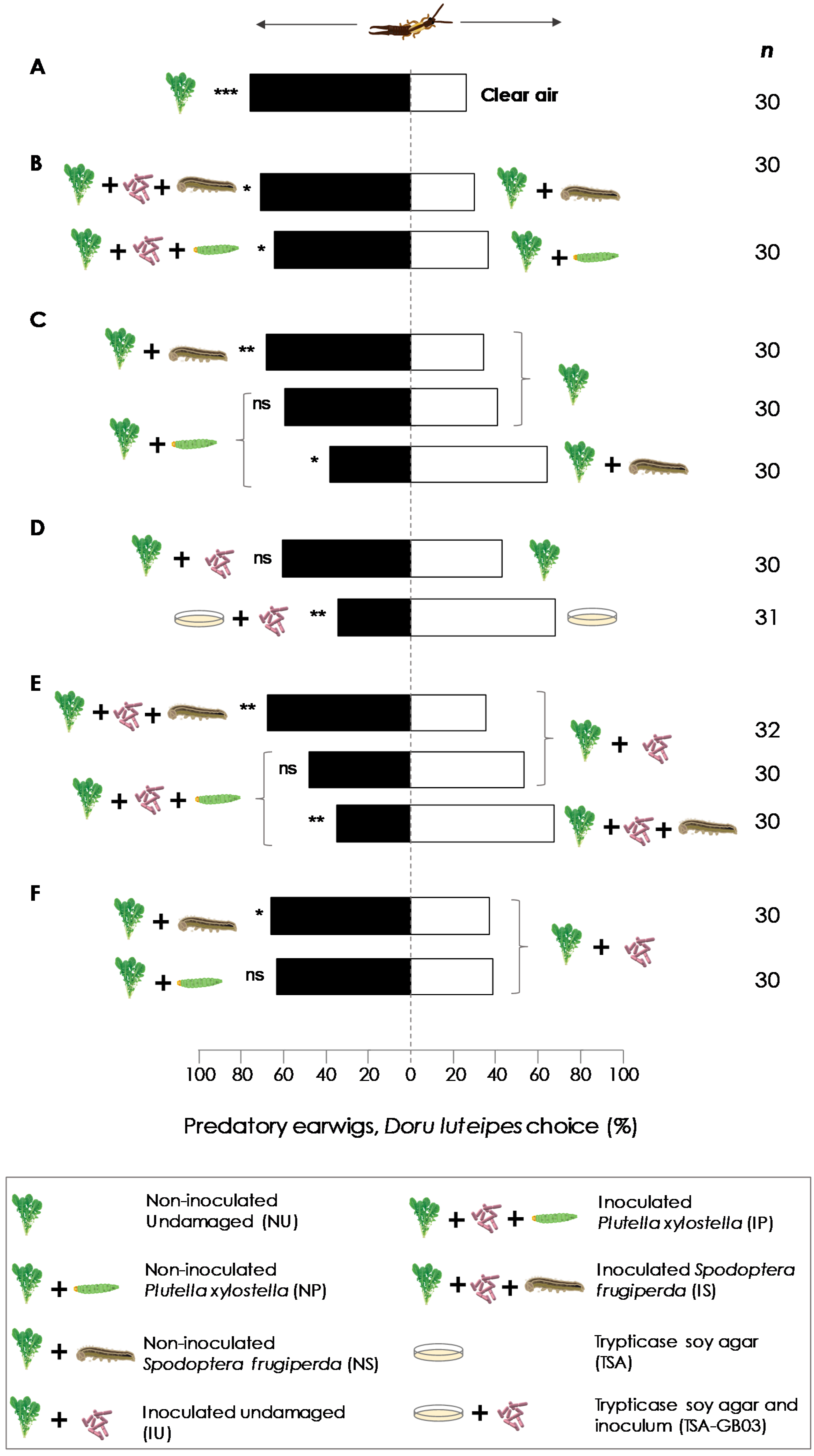
3.2. Y-Tube Olfactometer Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Karban, R.; Baldwin, I.T. Induced Responses to Herbivory, 1st ed.; The University of Chicago Press: Chicago, IL, USA, 1997; p. 330. [Google Scholar]
- Wittstock, U.; Gershenzon, J. Constitutive plant toxins and their role in defense against herbivores and pathogens. Curr. Opin. Plant. Biol. 2002, 5, 300–307. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect Plant Biology; Oxford University Press: New York, NY, USA, 2005; pp. 48–86. [Google Scholar]
- Price, P.W.; Bouton, C.E.; Gross, P.; McPheron, B.A.; Thompson, J.N.; Weis, A.E. Interactions among three trophic levels: Influence of plants on interactions between insect herbivores and natural enemies. Annu. Rev. Ecol. Syst. 1980, 11, 41–65. [Google Scholar] [CrossRef] [Green Version]
- Turlings, T.C.J.; Tumlinson, J.H.; Lewis, W.J. Exploitation of herbivore-induced plant odors by host-seeking parasitic wasps. Science 1990, 250, 1251–1253. [Google Scholar] [CrossRef] [Green Version]
- De Moraes, C.M.; Lewis, W.J.; Pare, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 394, 570. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Mescher, M.C.; Tumlinson, J.H. Caterpillar-induced nocturnal plant volatiles repel conspecific females. Nature 2001, 410, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M.; van Loon, J.J.A. Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context. Entomol. Exp. Appl. 2000, 97, 237–249. [Google Scholar] [CrossRef] [Green Version]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef]
- Stenberg, J.A.; Heil, M.; Ahman, I.; Bjorkman, C. Optimizing crops for biocontrol of pests and disease. Trends Plant Sci. 2015, 20, 698–712. [Google Scholar] [CrossRef] [Green Version]
- De Boer, J.G.; Hordijk, C.A.; Posthumus, M.A.; Dicke, M. Prey and non-prey arthropods sharing a host plant: Effects on induced volatile emission and predator attraction. J. Chem. Ecol. 2008, 34, 281. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.; Kroymann, J.; Mitchell-Olds, T. Different transcript patterns in response to specialist and generalist herbivores in the wild Arabidopsis relative Boechera divaricarpa. PLoS ONE 2007, 2, 1081. [Google Scholar] [CrossRef] [Green Version]
- Travers-Martin, N.; Müller, C. Matching plant defence syndromes with performance and preference of a specialist herbivore. Funct. Ecol. 2008, 22, 1033–1043. [Google Scholar] [CrossRef]
- Mooney, E.H.; Tiedeken, E.J.; Muth, N.Z.; Niesenbaum, R.A. Differential induced response to generalist and specialist herbivores by Lindera benzoin (Lauraceae) in sun and shade. Oikos 2009, 118, 1181–1189. [Google Scholar] [CrossRef]
- Du, Y.; Poppy, G.M.; Powell, W.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Identification of semiochemicals released during aphid feeding that attract parasitoid Aphidius ervi. J. Chem. Ecol. 1998, 24, 1355–1368. [Google Scholar] [CrossRef]
- Rasmann, S.; Turlings, T.C.J. Simultaneous feeding by aboveground and belowground herbivores attenuates plant-mediated attraction of their respective natural enemies. Ecol. Lett. 2007, 10, 926–936. [Google Scholar] [CrossRef]
- Ali, J.G.; Agrawal, A.A. Specialist versus generalist insect herbivores and plant defense. Trends Plant Sci. 2012, 17, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Van Poecke, R.M.; Roosjen, M.; Pumarino, L.; Dicke, M. Attraction of the specialist parasitoid Cotesia rubecula to Arabidopsis thaliana infested by host or non-host herbivore species. Entomol. Exp. Appl. 2003, 107, 229–236. [Google Scholar] [CrossRef]
- Guerrieri, E.; Lingua, G.; Digilio, M.C.; Massa, N.; Berta, G. Do interactions between plant roots and the rhizosphere affect parasitoid behaviour? Ecol. Entomol. 2004, 29, 753–756. [Google Scholar] [CrossRef]
- Godschalx, A.L.; Schädler, M.; Trisel, J.A.; Balkan, M.A.; Ballhorn, D.J. Ants are less attracted to the extrafloral nectar of plants with symbiotic, nitrogen-fixing rhizobia. Ecology 2015, 96, 348–354. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, M.; Erb, M.; Ton, J.; Brandenburg, A.; Karlen, D.; Zopfi, J.; Turlings, T.C. Volatiles produced by soil-borne endophytic bacteria increase plant pathogen resistance and affect tritrophic interactions. Plant Cell Environ. 2014, 37, 813–826. [Google Scholar] [CrossRef]
- Ryu, C.M.; Murphy, J.F.; Mysore, K.S.; Kloepper, J.W. Plant growth-promoting rhizobacteria systemically protect Arabidopsis thaliana against Cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway. Plant J. 2004, 39, 381–392. [Google Scholar] [CrossRef]
- Blom, D.; Fabbri, C.; Connor, E.C.; Schiestl, F.P.; Klauser, D.R.; Boller, T.; Eberl, L.; Weisskopf, L. Production of plant growth modulating volatiles is widespread among rhizosphere bacteria and strongly depends on culture conditions. Environ. Microbiol. 2011, 13, 3047–3058. [Google Scholar] [CrossRef] [PubMed]
- Bailly, A.; Weisskopf, L. The modulating effect of bacterial volatiles on plant growth: Current knowledge and future challenges. Plant Signal Behav. 2012, 7, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Pineda, A.; Soler, R.; Weldegergis, B.T.; Shimwela, M.M.; van Loon, J.J.A.; Dicke, M. Non-Pathogenic rhizobacteria interfere with the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid signalling. Plant Cell Environ. 2013, 36, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Van der Putten, W.H.; Vet, L.E.M.; Harvey, J.A.; Wackers, F.L. Linking above and belowground multitrophic interactions of plants, herbivores, pathogens, and their antagonists. Trends Ecol. Evol. 2001, 16, 547–554. [Google Scholar] [CrossRef]
- Pineda, A.; Zheng, S.J.; Van Loon, J.J.; Pieterse, C.M.; Dicke, M. Helping plants to deal with insects: The role of beneficial soil-borne microbes. Trends Plant Sci. 2010, 15, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.; Nadipalli, R.K.; Xie, X.; Sun, Y.; Surowiec, K.; Zhang, J.; Pare, P.W. Augmenting Sulfur Metabolism and Herbivore Defense in Arabidopsis by Bacterial Volatile Signaling. Front. Plant Sci. 2016, 7, 458. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.T.; Backman, P.A. Factors relating to peanut yield increases after seed treatment with Bacillus subtilis. Plant Dis. 1991, 75, 347–353. [Google Scholar] [CrossRef]
- Backman, P.; Brannen, P.M.; Mahaffee, W.F. Plant response and disease control following seed inoculation with Bacillus subtilis. In Improving Plant Productivity with Rhizosphere Bacteria; Ryder, M.H., Stephens, P.M., Bowen, G.D., Eds.; CSIRO Division of Soils: Glen Osmond, Australia, 1994; pp. 3–8. [Google Scholar]
- Fernando, W.D.; Ramarathnam, R.; Krishnamoorthy, A.S.; Savchuk, S.C. Identification and use of potential bacterial organic antifungal volatiles in biocontrol. Soil Biol. Biochem. 2005, 37, 955–964. [Google Scholar] [CrossRef]
- Liu, C.; Yin, X.; Wang, Q.; Peng, Y.; Ma, Y.; Liu, P.; Shi, J. Antagonistic activities of volatiles produced by two Bacillus strains against Monilinia fructicola in peach fruit. J. Sci. Food Agric. 2018, 98, 5756–5763. [Google Scholar] [CrossRef]
- Massawe, V.C.; Rao, A.H.; Farzand, A.; Mburu, D.K.; Gao, X. Volatile Organic Compounds of Endophytic Bacillus spp. have Biocontrol Activity Against Sclerotinia sclerotiorum. Phytopathology 2018, 108. [Google Scholar]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Weyens, N.; Van Der Lelie, D.; Taghavi, S.; Newman, L.; Vangrondsveld, J. Exploiting plant-microbe partnerships to improve biomass production and remediation. Trends Biotechnol. 2009, 27, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore-induced plant volatiles: Beyond the ‘cry for help’. Trends Plant Sci. 2010, 627, 167–175. [Google Scholar] [CrossRef]
- Kessler, A.; Heil, M. The multiple faces of indirect defences and their agents of natural selection. Funct. Ecol. 2011, 25, 348–357. [Google Scholar] [CrossRef]
- Stam, J.M.; Kroes, A.; Li, Y.; Gols, R.; van Loon, J.J.; Poelman, E.H.; Dicke, M. Plant interactions with multiple insect herbivores: From community to genes. Annu. Rev. Plant Biol. 2014, 65, 689–713. [Google Scholar] [CrossRef] [PubMed]
- Rasmann, S.; Bennett, A.; Biere, A.; Karley, A.; Guerrieri, E. Root symbionts: Powerful drivers of plant above-and belowground indirect defenses. Insect Sci. 2017, 24, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, D.; Bossi, S.; Cascone, P.; Digilio, M.C.; Prieto, J.D.; Fanti, P.; Guerrieri, E.; Iodice, L.; Lingua, G.; Lorito, M.; et al. Tomato below ground-above ground interactions: Trichoderma longibrachiatum affects the performance of Macrosiphum euphorbiae and its natural antagonists. Mol. Plant-Microbe Interact. 2013, 26, 1249–1256. [Google Scholar] [CrossRef] [Green Version]
- Saravanakumar, D.; Lavanya, N.; Muthumeena, B.; Raguchander, T.; Suresh, S.; Samiyappan, R. Pseudomonas fluorescens enhances resistance and natural enemy population in rice plants against leaffolder pest. J. Appl. Entomol. 2008, 132, 469–479. [Google Scholar] [CrossRef]
- Pangesti, N.; Pineda, A.; Pieterse, C.M.J.; Dicke, M.; van Loon, J.J.A. Two-way plant-mediated interactions between root-associated microbes and insects: From ecology to mechanisms. Front. Plant Sci. 2013, 4, 414. [Google Scholar] [CrossRef] [Green Version]
- U.S. EPA. Pesticide Product Label, Kodiak Concentrate Biological Fungicide. Available online: https://www3.epa.gov/pesticides/chem_search/ppls/000264-00951-20050705.pdf (accessed on 7 May 2015).
- Choi, S.-K.; Jeong, H.; Kloepper, J.W.; Ryu, C.-M. Genome sequence of Bacillus amyloliquefaciens GB03, an active ingredient of the first commercial biological control product. Genome Announc. 2014, 2, e01092-14. [Google Scholar] [CrossRef] [Green Version]
- Shelton, A.M.; Collins, H.L.; Zhang, Y.J.; Wu, Q.J. The management of diamondback moth and other crucifer pests: Proceedings of the fifth international workshop. In Proceedings of the 5th International Workshop on the Management of Diamondback Moth and Other Crucifer Pests, Beijing, China, 21–24 October 2006; China Agricultural Science and Technology Press: Beijing, China, 2008. [Google Scholar]
- Alvarenga, C.D.; Vendramim, J.D.; Cruz, I. Efeito do Predador Doru luteipes (Scud.) Sobre o Crescimento Populacional de Schizaphis graminum (Rond.) em Diferentes Genótipos de Sorgo. Soc. Entomol. Bras. 1996, 25, 137–140. [Google Scholar]
- Bacci, L.; Picanço, M.C.; Gusmo, M.R.; Barreto, R.W.; Galvan, T.L. Selective insecticides to the earwing Doru luteipes (Scudder) used for controlling the green peach aphid in Brassicae. Hortic. Bras. 2002, 2, 174–179. [Google Scholar] [CrossRef] [Green Version]
- Reis, L.L.; Oliveira, L.J.; Cruz, I. Biologia e potencial de Doru luteipes no controle de Spodoptera frugiperda. Pesqui. Agropecuária Bras. 1988, 23, 333–342. [Google Scholar]
- Fenoglio, M.S.; Trumper, E.V. Influence of Weather Conditions and Density of Doru luteipes (Dermaptera: Forficulidae) on Diatraea saccharalis (Lepidoptera: Crambidae) Egg Mortality. Environ. Entomol. 2007, 36, 1159–1165. [Google Scholar] [CrossRef]
- Parra, J.R.P. Técnicas de Criação de Insetos Para Programas de Controle Biológico, 6th ed.; FEALQ: Piracicaba, Brasil, 2001; p. 134. [Google Scholar]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Pare, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharboutli, M.S.; Mack, T.P. Effect of temperature, humidity, and prey density on feeding rate of the striped earwig (Dermaptera: Labiduridae). Environ. Entomol. 1993, 22, 1134–1139. [Google Scholar] [CrossRef]
- Weiss, M.J.; McDonald, G. European earwig, Forficula auricularia L. (Dermaptera: Forficulidae), as a predator of the redlegged Earth mite, Halotydeus destructor (Tucker) (Acarina: Penthaleidae). Aust. Entomol. 1998, 37, 183–185. [Google Scholar]
- Naranjo-Guevara, N.; Peñaflor, M.F.G.; Cabezas-Guerrero, M.F.; Bento, J.M.S. Nocturnal herbivore-induced plant volatiles attract the generalist predatory earwig Doru luteipes Scudder. Sci. Nat. 2017, 104, 77. [Google Scholar] [CrossRef]
- Vessey, J. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Rodriguez, A.; Sanders, I.R. The role of community and population ecology in applying mycorrhizal fungi for improved food security. IMSE J. 2015, 9, 1053–1061. [Google Scholar] [CrossRef]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G.A. An underground revolution: Biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Bacci, L.; Picanco, M.C.; Gusmão, M.R.; Crespo, A.; Pereira, J.C. Seletividade de inseticidas a Brevicoryne brassica (L.) (Hemiptera: Aphididae) e ao predador Doru luteipes (Dermaptera: Forficulidae). Neotrop. Entomol. 2001, 30, 407–713. [Google Scholar] [CrossRef] [Green Version]
- Sueldo, M.R.; Bruzzone, O.A.; Virla, E.G. Characterization of the earwig, Doru lineare, as a predator of larvae of the fall armyworm, Spodoptera frugiperda: A functional response study. J. Insect Sci. 2010, 10, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva Nunes, G.; Dantas, T.A.V.; Figueiredo, W.R.S.; de Souza, M.D.S.; do Nascimento, I.N.; de Luna Batista, J. Predation of diamondback moth larvae and pupae by Euborellia annulipes. RBCA 2018, 8. [Google Scholar] [CrossRef]
- Takabayashi, J.; Sabelis, M.W.; Janssen, A.; Shiojiri, K.; van Wijk, M. Can plants betray the presence of multiple herbivore species to predators and parasitoids? The role of learning in phytochemical information networks. Ecol. Res. 2006, 21, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Painter, R.H. Insects on corn and teosinte in Guatemala. J. Econ. Entomol. 1955, 48, 36–42. [Google Scholar] [CrossRef]
- De Boer, J.G.; Dicke, M. Olfactory learning by predatory arthropods. Anim. Biol. 2006, 56, 143–155. [Google Scholar] [CrossRef]
- Dukas, R. Evolutionary biology of insect learning. Annu. Rev. Entomol. 2008, 53, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Vet, L.E.; Dicke, M. Ecology of infochemical use by natural enemies in a tritophic context. Annu. Rev. Entomol. 1992, 37, 141–172. [Google Scholar] [CrossRef]
- Ngumbi, E.N. Mechanisms of Olfaction in Parasitic Wasps: Analytical and Behavioral Studies of Response of a Specialist (Microplitis croceipes) and a Generalist (Cotesia marginiventris) Parasitoid to Host-Related Odor. Doctoral Dissertation, Auburn University, Auburn, AL, USA, 2011. [Google Scholar]
- Pangesti, N.; Weldegergis, B.T.; Langendorf, B.; van Loon, J.J.; Dicke, M.; Pineda, A. Rhizobacterial colonization of roots modulates plant volatile emission and enhances the attraction of a parasitoid wasp to host-infested plants. Oecologia 2015, 178, 1169–1180. [Google Scholar] [CrossRef] [Green Version]
- Ruther, J.; Kleier, S. Plant-plant signaling: Ethylene synergizes volatile emission in Zea mays induced by exposure to (Z)-3-hexen-1-ol. J. Chem. Ecol. 2005, 31, 2217–2222. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, J.I.; Arimura, G.I.; Ozawa, R.; Shimoda, T.; Takabayashi, J.; Nishioka, T. Exogenous ACC enhances volatiles production mediated by jasmonic acid in lima bean leaves. FEBS Lett. 2001, 509, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.H.; Song, G.C.; Ryu, C.M. Sweet scents from good bacteria: Case studies on bacterial volatile compounds for plant growth and immunity. Plant Mol. Biol. 2016, 90, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial Volatile Emissions as Insect Semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell, K.; Naranjo-Guevara, N.; Santos, R.C.d.; Meadow, R.; Bento, J.M.S. Predatory Earwigs are Attracted by Herbivore-Induced Plant Volatiles Linked with Plant Growth-Promoting Rhizobacteria. Insects 2020, 11, 271. https://doi.org/10.3390/insects11050271
Bell K, Naranjo-Guevara N, Santos RCd, Meadow R, Bento JMS. Predatory Earwigs are Attracted by Herbivore-Induced Plant Volatiles Linked with Plant Growth-Promoting Rhizobacteria. Insects. 2020; 11(5):271. https://doi.org/10.3390/insects11050271
Chicago/Turabian StyleBell, Kim, Natalia Naranjo-Guevara, Rafaela C. dos Santos, Richard Meadow, and José M. S. Bento. 2020. "Predatory Earwigs are Attracted by Herbivore-Induced Plant Volatiles Linked with Plant Growth-Promoting Rhizobacteria" Insects 11, no. 5: 271. https://doi.org/10.3390/insects11050271
APA StyleBell, K., Naranjo-Guevara, N., Santos, R. C. d., Meadow, R., & Bento, J. M. S. (2020). Predatory Earwigs are Attracted by Herbivore-Induced Plant Volatiles Linked with Plant Growth-Promoting Rhizobacteria. Insects, 11(5), 271. https://doi.org/10.3390/insects11050271