Does the Infectious Status of Aphids Influence Their Preference Towards Healthy, Virus-Infected and Endophytically Colonized Plants?

 , ,
, ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Cultures
2.2. Virus and EEPF Cultures
2.3. Insect Rearing
2.4. EEPF-Inoculated and Virus-Infected Plants
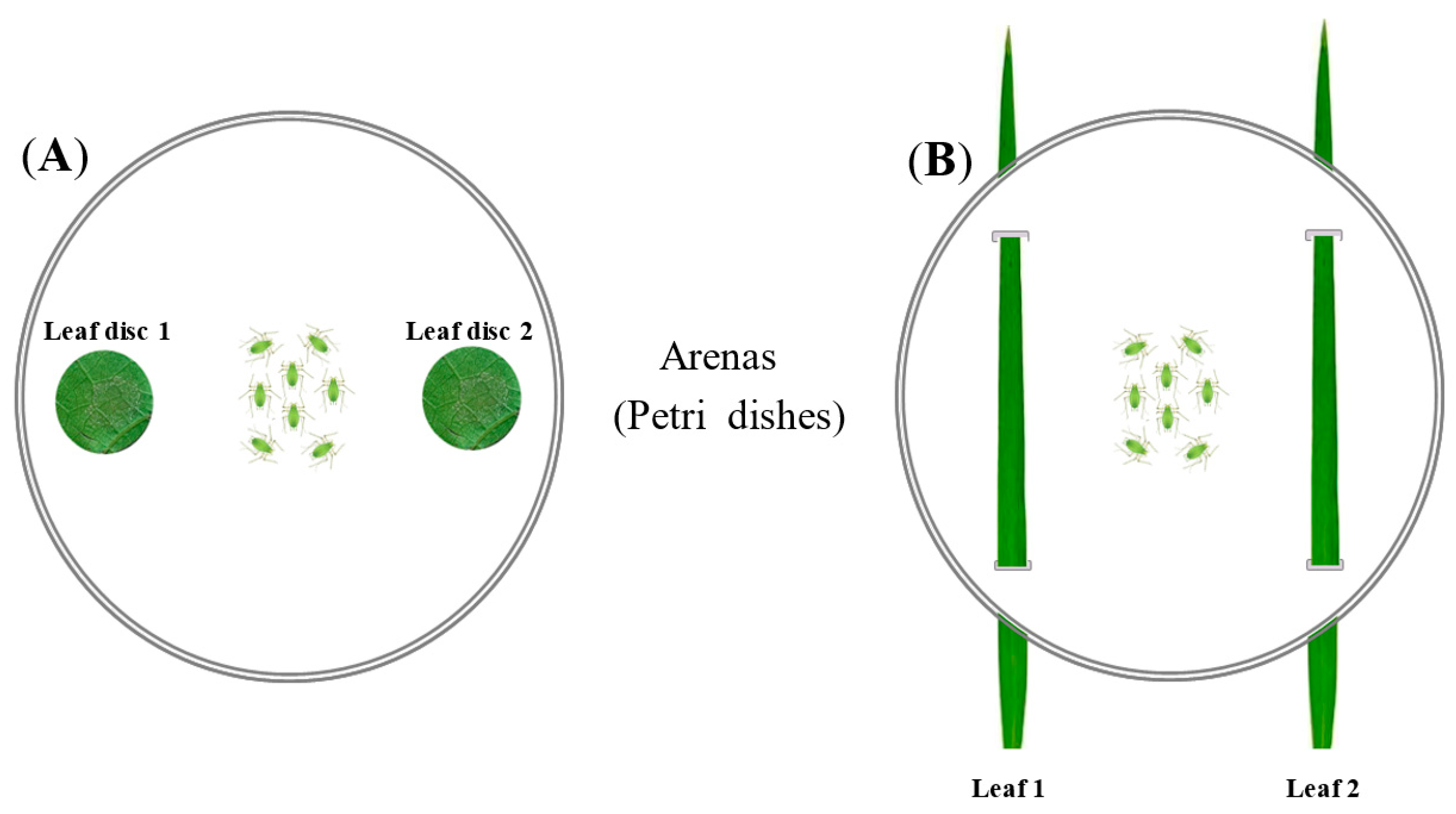
2.5. Design of the Preference Bioassay
2.6. VOC Sampling and Analysis
2.7. Statistical Analyses
3. Results
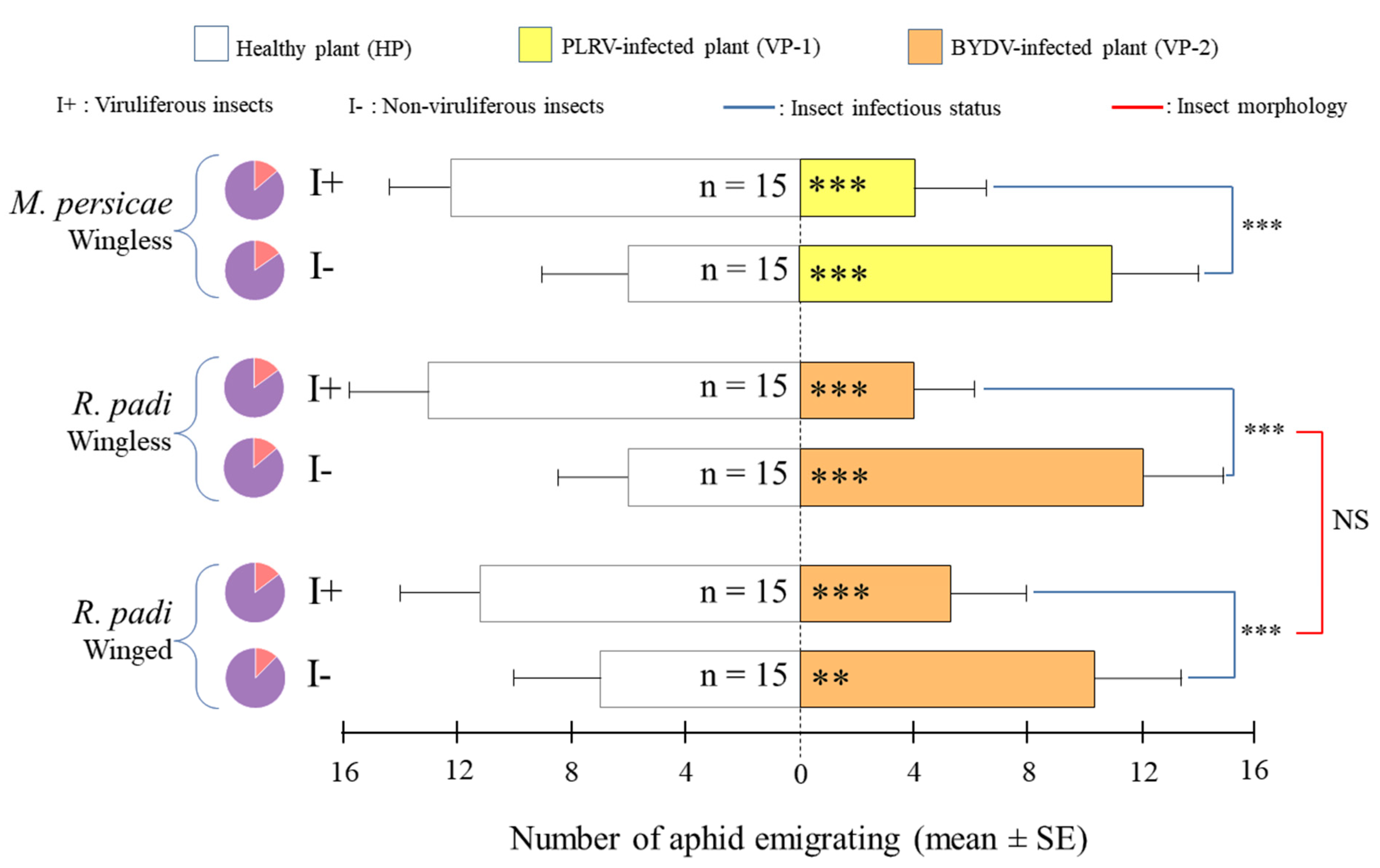
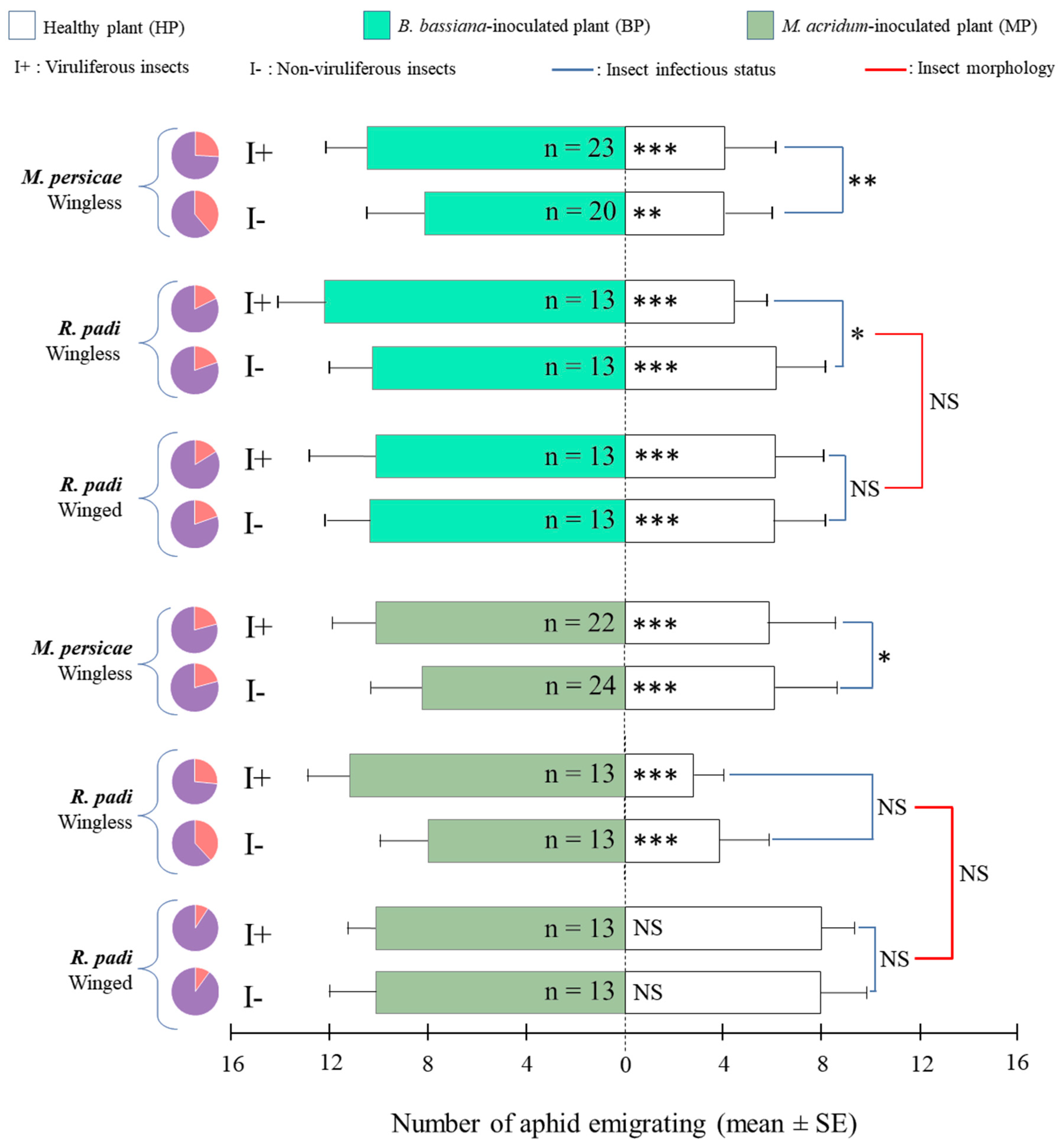
3.1. Aphid Preference
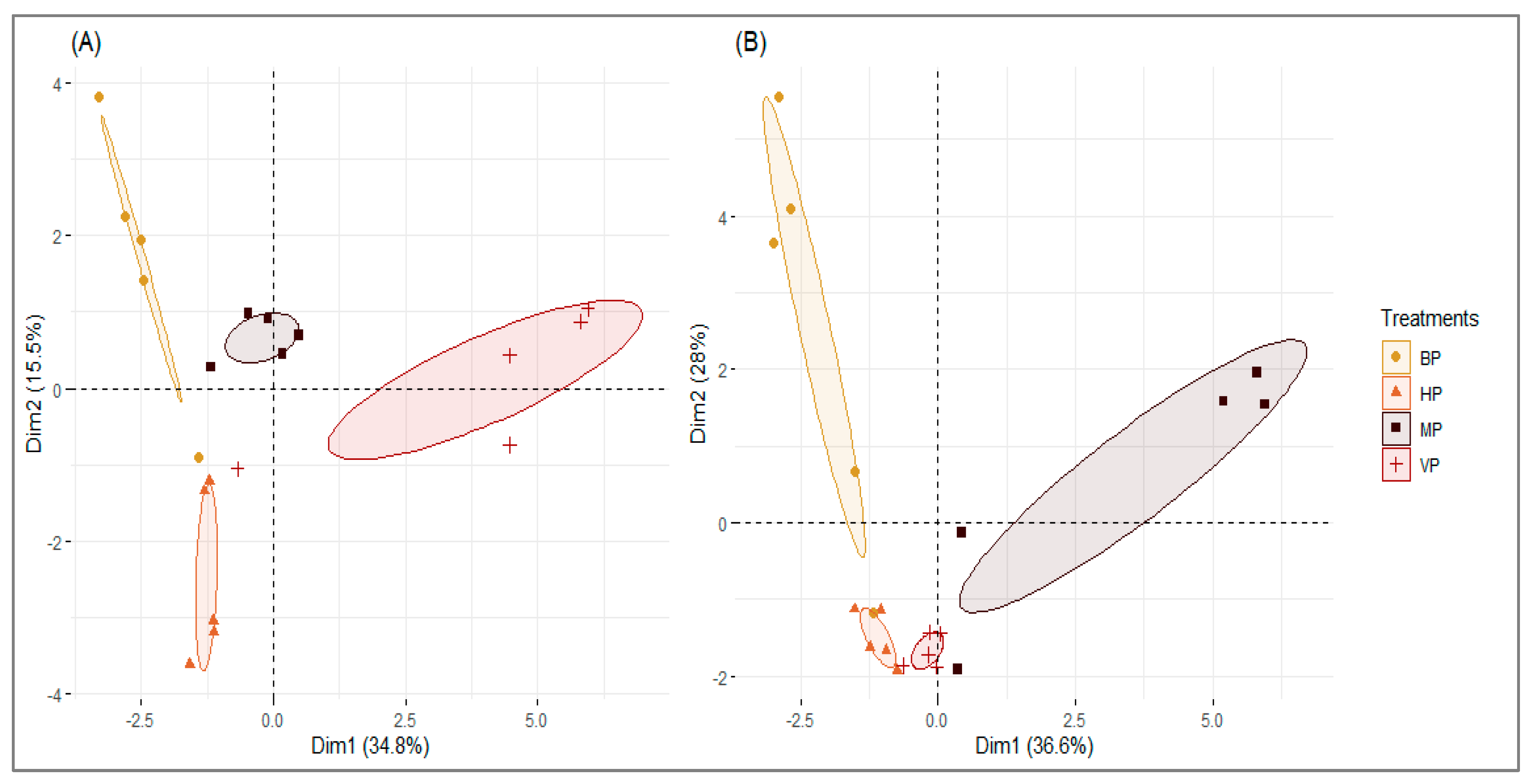
3.2. Organic Compounds of Volatiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Llewellyn, K.S.; Loxdale, H.D.; Harrington, R.; Brookes, C.P.; Clark, S.J.; Sunnucks, P. Migration and genetic structure of the grain aphid (Sitobion avenae) in Britain related to climate and clonal fluctuation as revealed using microsatellites. Mol. Ecol. 2002, 12, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Nault, L.R. Arthropod Transmission of Plant Viruses: A New Synthesis. Ann. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Kamphuis, L.G.; Zulak, K.; Gao, L.-L.; Anderson, J.; Singh, K.B. Plant–aphid interactions with a focus on legumes. Funct. Plant Biol. 2013, 40, 1271. [Google Scholar] [CrossRef] [PubMed]
- Powell, G.; Tosh, C.R.; Hardie, J. Host plant selection by aphids: Behavioral, Evolutionary, and Applied Perspectives. Annu. Rev. Entomol. 2006, 51, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Hussain, M.; Avery, P.B.; Qasim, M.; Fang, D.; Wang, L. Volatiles from plants induced by multiple aphid attacks promote conidial performance of Lecanicillium lecanii. PLoS ONE 2016, 11, e0151844. [Google Scholar] [CrossRef] [PubMed]
- Boquel, S.; Giordanengo, P.; Ameline, A. Vector activity of three aphid species (Hemiptera: Aphididae) modulated by host plant selection behaviour on potato (Solanales: Solanaceae). Ann. Soc. Entomol. Fr. 2014, 50, 141–148. [Google Scholar] [CrossRef]
- Döring, T.F. How aphids find their host plants, and how they don’t. Ann. Appl. Biol. 2014, 165, 3–26. [Google Scholar] [CrossRef]
- Ngumbi, E.; Eigenbrode, S.D.; Bosque-Pérez, N.A.; Ding, H.; Rodriguez, A. Myzus persicae is arrested more by blends than by individual compounds elevated in headspace of plrv-infected potato. J. Chem. Ecol. 2007, 33, 1733–1747. [Google Scholar] [CrossRef]
- He, X.; Xu, H.; Gao, G.; Zhou, X.; Zheng, X.; Sun, Y.; Yang, Y.; Tian, J.; Lu, Z. Virus-mediated chemical changes in rice plants impact the relationship between non-vector planthopper Nilaparvata lugens Stål and its egg parasitoid Anagrus nilaparvatae Pang et Wang. PLoS ONE 2014, 9, e105373. [Google Scholar] [CrossRef]
- Ingwell, L.L.; Eigenbrode, S.D.; Bosque-Pérez, N.A. Plant viruses alter insect behavior to enhance their spread. Sci. Rep. 2012, 2, 1–6. [Google Scholar] [CrossRef]
- Rajabaskar, D.; Bosque-Pérez, N.A.; Eigenbrode, S.D. Preference by a virus vector for infected plants is reversed after virus acquisition. Virus Res. 2014, 186, 32–37. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.L.; Moscardini, V.F.; Gontijo, P.C.; Sâmia, R.R.; Marucci, R.C.; Budia, F.; Carvalho, G.A. Life history parameters and feeding preference of the green lacewing Ceraeochrysa cubana fed with virus-free and potato leafroll virus-infected Myzus persicae. BioControl 2016, 61, 671–679. [Google Scholar] [CrossRef]
- Carmo-Sousa, M.; Moreno, A.; Plaza, M.; Garzo, E.; Fereres, A. Cucurbit aphid-borne yellows virus (CABYV) modifies the alighting, settling and probing behaviour of its vector Aphis gossypii favouring its own spread. Ann. Appl. Biol. 2016, 169, 284–297. [Google Scholar] [CrossRef]
- dos Santos, R.C.; Peñaflor, M.F.G.V.; Sanches, P.A.; Nardi, C.; Bento, J.M.S. The effects of Gibberella zeae, Barley Yellow Dwarf Virus, and co-infection on Rhopalosiphum padi olfactory preference and performance. Phytoparasitica 2016, 44, 47–54. [Google Scholar] [CrossRef]
- Calvo, D.; Fereres, A. The performance of an aphid parasitoid is negatively affected by the presence of a circulative plant virus. BioControl 2011, 56, 747–757. [Google Scholar] [CrossRef]
- Jahan, S.M.H.; Lee, G.-S.; Lee, S.; Lee, K.-Y. Acquisition of Tomato yellow leaf curl virus enhances attraction of Bemisia tabaci to green light emitting diodes. J. Asia Pac. Entomol. 2014, 17, 79–82. [Google Scholar] [CrossRef]
- Vu, V.H.; Hong, S., Il; Kim, K. Selection of entomopathogenic fungi for aphid control. J. Biosci. Bioeng. 2007, 104, 498–505. [Google Scholar] [CrossRef]
- Seye, F.; Bawin, T.; Boukraa, S.; Zimmer, J.Y.; Ndiaye, M.; Delvigne, F.; Francis, F. Effect of entomopathogenic Aspergillus strains against the pea aphid, Acyrthosiphon pisum (Hemiptera: Aphididae). Appl. Entomol. Zool. 2014, 49, 453–458. [Google Scholar] [CrossRef]
- Elliot, S.L.; Sabelis, M.W.; Janssen, A.; Van der Geest, L.P.S.; Beerling, E.A.M.; Fransen, J. Can plants use entomopathogens as bodyguards? Ecol. Lett. 2000, 3, 228–235. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Catherine Aime, M.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control 2008, 46, 72–82. [Google Scholar] [CrossRef]
- Parsa, S.; Ortiz, V.; Vega, F.E. Establishing Fungal Entomopathogens as Endophytes: Towards Endophytic Biological Control. J. Vis. Exp. 2013, e50360. [Google Scholar] [CrossRef] [PubMed]
- Biswas, C.; Dey, P.; Satpathy, S.; Satya, P. Establishment of the fungal entomopathogen Beauveria bassiana as a season long endophyte in jute (Corchorus olitorius) and its rapid detection using SCAR marker. BioControl 2012, 57, 565–571. [Google Scholar] [CrossRef]
- Russo, M.L.; Pelizza, S.A.; Cabello, M.N.; Stenglein, S.A.; Scorsetti, A.C. Endophytic colonisation of tobacco, corn, wheat and soybeans by the fungal entomopathogen Beauveria bassiana (Ascomycota, Hypocreales). Biocontrol Sci. Technol. 2015, 25, 475–480. [Google Scholar] [CrossRef]
- Behie, S.W.; Jones, S.J.; Bidochka, M.J. Plant tissue localization of the endophytic insect pathogenic fungi Metarhizium and Beauveria. Fungal Ecol. 2015, 13, 112–119. [Google Scholar] [CrossRef]
- McKinnon, A.C. Plant Tissue Preparation for the Detection of an Endophytic Fungus In Planta. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2016; Volume 1477, pp. 167–173. [Google Scholar] [CrossRef]
- Gurulingappa, P.; Sword, G.A.; Murdoch, G.; McGee, P.A. Colonization of crop plants by fungal entomopathogens and their effects on two insect pests when in planta. Biol. Control 2010, 55, 34–41. [Google Scholar] [CrossRef]
- González-Mas, N.; Sánchez-Ortiz, A.; Valverde-García, P.; Quesada-Moraga, E. Effects of Endophytic Entomopathogenic Ascomycetes on the Life-History Traits of Aphis gossypii Glover and Its Interactions with Melon Plants. Insects 2019, 10, 165. [Google Scholar] [CrossRef]
- Gurulingappa, P.; McGee, P.A.; Sword, G. Endophytic Lecanicillium lecanii and Beauveria bassiana reduce the survival and fecundity of Aphis gossypii following contact with conidia and secondary metabolites. Crop Prot. 2011, 30, 349–353. [Google Scholar] [CrossRef]
- Clifton, E.H.; Jaronski, S.T.; Coates, B.S.; Hodgson, E.W.; Gassmann, A.J. Effects of endophytic entomopathogenic fungi on soybean aphid and identification of Metarhizium isolates from agricultural fields. PLoS ONE 2018, 13, e0194815. [Google Scholar] [CrossRef]
- Jaber, L.R.; Araj, S.-E. Interactions among endophytic fungal entomopathogens (Ascomycota: Hypocreales), the green peach aphid Myzus persicae Sulzer (Homoptera: Aphididae), and the aphid endoparasitoid Aphidius colemani Viereck (Hymenoptera: Braconidae). Biol. Control 2018, 116, 53–61. [Google Scholar] [CrossRef]
- Meister, B.; Krauss, J.; Härri, S.A.; Victoria Schneider, M.; Müller, C.B. Fungal endosymbionts affect aphid population size by reduction of adult life span and fecundity. Basic Appl. Ecol. 2006, 7, 244–252. [Google Scholar] [CrossRef]
- Chauzat, M.; Faucon, J. Pesticide residues in beeswax samples collected from honey bee colonies (Apis mellifera L.) in France. Pest Manag. Sci. 2007, 1106, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Tzeng, Y. Development and applications of destruxins: A review. Biotechnol. Adv. 2012, 30, 1242–1254. [Google Scholar] [CrossRef] [PubMed]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2017, 107, 50–59. [Google Scholar] [CrossRef]
- Thakur, A.; Kaur, S.; Kaur, A.; Singh, V. Enhanced Resistance to Spodoptera litura in Endophyte Infected Cauliflower Plants. Environ. Entomol. 2013, 42, 240–246. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, A.R.; Raya-Díaz, S.; Zamarreño, Á.M.; García-Mina, J.M.; del Campillo, M.C.; Quesada-Moraga, E. An endophytic Beauveria bassiana strain increases spike production in bread and durum wheat plants and effectively controls cotton leafworm (Spodoptera littoralis) larvae. Biol. Control 2018, 116, 90–102. [Google Scholar] [CrossRef]
- Van Wees, S.C.; Van der Ent, S.; Pieterse, C.M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef]
- Aragón, S.M. How Entomopathogenic Endophytic Fungi Modulate Plant-Insect Interactions; Georg-August-University Goettingen: Göttingen, Germany, 2016. [Google Scholar]
- Jaber, L.R.; Vidal, S. Fungal endophyte negative effects on herbivory are enhanced on intact plants and maintained in a subsequent generation. Ecol. Entomol. 2010, 35, 25–36. [Google Scholar] [CrossRef]
- Saikkonen, K.; Gundel, P.E.; Helander, M. Chemical Ecology Mediated by Fungal Endophytes in Grasses. J. Chem. Ecol. 2013, 39, 962–968. [Google Scholar] [CrossRef]
- Li, T.; Blande, J.D.; Gundel, P.E.; Helander, M.; Saikkonen, K. Epichloë Endophytes Alter Inducible Indirect Defences in Host Grasses. PLoS ONE 2014, 9, e101331. [Google Scholar] [CrossRef]
- Hodge, S.; Powell, G. Complex interactions between a plant pathogen and insect parasitoid via the shared vector-host: Consequences for host plant infection. Oecologia 2008, 157, 387–397. [Google Scholar] [CrossRef]
- Dáder, B.; Moreno, A.; Viñuela, E.; Fereres, A. Spatio-temporal dynamics of viruses are differentially affected by parasitoids depending on the mode of transmission. Viruses 2012, 4, 3069–3089. [Google Scholar] [CrossRef]
- Lin, F.-J.; Bosquée, E.; Liu, Y.-J.; Chen, J.-L.; Yong, L.; Francis, F. Impact of aphid alarm pheromone release on virus transmission efficiency: When pest control strategy could induce higher virus dispersion. J. Virol. Methods 2016, 235, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Woodford, J.A.T.; Jolly, C.A.; Aveyard, C.S. Biological factors influencing the transmission of potato leafroll virus by different aphid species. Potato Res. 1995, 38, 133–141. [Google Scholar] [CrossRef]
- LUBILOSA. Locust Control and the Making of “Green Muscle”. Available online: http://www.dropdata.org/biopesticides/green_muscle.htm (accessed on 17 June 2020).
- Inglis, G.D.; Enkerli, J.; Goettel, M.S. Laboratory Techniques Used for Entomopathogenic Fungi BT—Manual of Techniques in Invertebrate Pathology, 2nd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Rondot, Y.; Reineke, A. Endophytic Beauveria bassiana in grapevine Vitis vinifera (L.) reduces infestation with piercing-sucking insects. Biol. Control 2018, 116, 82–89. [Google Scholar] [CrossRef]
- Humber, R.A. Fungi. In Manual of Techniques in Insect Pathology; Elsevier: Amsterdam, The Netherlands, 1997; pp. 153–185. [Google Scholar] [CrossRef]
- Driver, F.; Milner, R.J.; Trueman, J.W.H. A taxonomic revision of Metarhizium based on a phylogenetic analysis of rDNA sequence data. Mycol. Res. 2000, 104, 134–150. [Google Scholar] [CrossRef]
- Haas, J.; Lozano, E.R.; Poppy, G.M. A simple, light clip-cage for experiments with aphids. Agric. For. Entomol. 2018, 20, 589–592. [Google Scholar] [CrossRef]
- Nisbet, A.J.; Woodford, J.A.; Strang, R.H.C. The effects of azadirachtin on the acquisition and inoculation of potato leafroll virus by Myzus persicae. Crop Prot. 1996, 15, 9–14. [Google Scholar] [CrossRef]
- Sadeghi, E.; Dedryver, C.; Gauthier, J. Role of acquisition and inoculation time in the expression of clonal variation for BYDV-PAV transmission in the aphid species Rhopalosiphum padi. Plant Pathol. 1997, 46, 502–508. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Kazantsev, T.; Talts, E.; Tosens, T.; Niinemets, Ü. Emission Timetable and Quantitative Patterns of Wound-Induced Volatiles Across Different Leaf Damage Treatments in Aspen (Populus Tremula). J. Chem. Ecol. 2015, 41, 1105–1117. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Haubruge, E.; De Moraes, C.M.; Mescher, M.C. Aphid responses to volatile cues from turnip plants (Brassica rapa) infested with phloem-feeding and chewing herbivores. Arthropod. Plant. Interact. 2013, 7, 567–577. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 2 March 2020).
- The Pherobase: Database of Pheromones and Semiochemicals. Available online: https://www.pherobase.com/ (accessed on 2 March 2020).
- NIST Chemistry WebBook, NIST Standard Reference Database Number 69; Linstrom, P.J., Mallard, W.G., Eds.; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2017. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M.A.S.S. The vegan package. Community Ecol. Packag. 2007, 10, 631–637. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Package ‘factoextra’: Extract and Visualize the Results of Multivariate Data Analyse. Available online: http://www.sthda.com/english/rpkgs/factoextra (accessed on 2 March 2020).
- Hodge, S.; Powell, G. Conditional Facilitation of an Aphid Vector, Acyrthosiphon pisum, by the Plant Pathogen, Pea Enation Mosaic Virus. J. Insect Sci. 2010, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.W.; Wang, X.F.; Chen, D.; Wang, X.W.; Fan, X.J.; Liu, X.D. Potato virus Y-infected tobacco affects the growth, reproduction, and feeding behavior of a vector aphid, Myzus persicae (Hemiptera: Aphididae). Appl. Entomol. Zool. 2015, 50, 239–243. [Google Scholar] [CrossRef]
- Ghosh, A.; Das, A.; Vijayanandraj, S.; Mandal, B. Cardamom Bushy Dwarf Virus Infection in Large Cardamom Alters Plant Selection Preference, Life Stages, and Fecundity of Aphid Vector, Micromyzus kalimpongensis (Hemiptera: Aphididae). Environ. Entomol. 2016, 45, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Congdon, B.S.; Coutts, B.A.; Renton, M.; Flematti, G.R.; Jones, R.A.C. Establishing alighting preferences and species transmission differences for Pea seed-borne mosaic virus aphid vectors. Virus Res. 2017, 241, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Effects of pathogens on sensory-mediated interactions between plants and insect vectors. Curr. Opin. Plant Biol. 2016, 32, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Nikolakakis, N.N.; Margaritopoulos, J.T.; Tsitsipis, J.A. Performance of Myzus persicae (Hemiptera: Aphididae) clones on different host-plants and their host preference. Bull. Entomol. Res. 2003, 93, 235–242. [Google Scholar] [CrossRef]
- Sharifi, R.; Lee, S.-M.; Ryu, C.-M. Microbe-induced plant volatiles. New Phytol. 2018, 220, 684–691. [Google Scholar] [CrossRef]
- Quiroz, A.; Niemeyer, H.M. Olfactometer-assesed responses of aphid Rhopalosiphum padi to wheat and oat volatiles. J. Chem. Ecol. 1998, 24, 113–124. [Google Scholar] [CrossRef]
- Dheivasigamani, R.; Ding, H.; Food, U.S.; Eigenbrode, S.D. Behavioral Responses of Green Peach Aphid, Myzus persicae (Sulzer), to the Volatile Organic Compound Emissions from Four Potato Varieties. Am. J. Potato Res. 2013, 90, 171–178. [Google Scholar] [CrossRef]
- González-Mas, N.; Quesada-Moraga, E.; Plaza, M.; Fereres, A.; Moreno, A. Changes in feeding behaviour are not related to the reduction in the transmission rate of plant viruses by Aphis gossypii (Homoptera: Aphididae) to melon plants colonized by Beauveria bassiana (Ascomycota: Hypocreales). Biol. Control 2019, 130, 95–103. [Google Scholar] [CrossRef]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant–endophyte–herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–54. [Google Scholar] [CrossRef]
- de Oliveira, C.F.; Long, E.Y.; Finke, D.L. A negative effect of a pathogen on its vector? A plant pathogen increases the vulnerability of its vector to attack by natural enemies. Oecologia 2014, 174, 1169–1177. [Google Scholar] [CrossRef][Green Version]
- González-Mas, N.; Medina, M.C.; Sánchez, F.G.; Moraga, E.Q. Bottom—Up effects of endophytic Beauveria bassiana on multitrophic interactions between the cotton aphid, Aphis gossypii, and its natural enemies in melon. J. Pest Sci. 2019, 92, 1271–1281. [Google Scholar] [CrossRef]
- Pedrini, N.; Crespo, R.; Juárez, M.P. Biochemistry of insect epicuticle degradation by entomopathogenic fungi. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 124–137. [Google Scholar] [CrossRef]
- Bawin, T.; Seye, F.; Boukraa, S.; Zimmer, J.-Y.; Delvigne, F.; Francis, F. La lutte contre les moustiques (Diptera: Culicidae): Diversité des approches et application du contrôle biologique. Can. Entomol. 2015, 147, 476–500. [Google Scholar] [CrossRef]
- Jaronski, S.T.; Mascarin, G.M. Mass Production of Fungal Entomopathogens. In Microbial Control of Insect and Mite Pests; Elsevier: Amsterdam, The Netherlands, 2017; pp. 141–155. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fingu-Mabola, J.C.; Martin, C.; Bawin, T.; Verheggen, F.J.; Francis, F. Does the Infectious Status of Aphids Influence Their Preference Towards Healthy, Virus-Infected and Endophytically Colonized Plants? Insects 2020, 11, 435. https://doi.org/10.3390/insects11070435
Fingu-Mabola JC, Martin C, Bawin T, Verheggen FJ, Francis F. Does the Infectious Status of Aphids Influence Their Preference Towards Healthy, Virus-Infected and Endophytically Colonized Plants? Insects. 2020; 11(7):435. https://doi.org/10.3390/insects11070435
Chicago/Turabian StyleFingu-Mabola, Junior Corneille, Clément Martin, Thomas Bawin, François Jean Verheggen, and Frédéric Francis. 2020. "Does the Infectious Status of Aphids Influence Their Preference Towards Healthy, Virus-Infected and Endophytically Colonized Plants?" Insects 11, no. 7: 435. https://doi.org/10.3390/insects11070435
APA StyleFingu-Mabola, J. C., Martin, C., Bawin, T., Verheggen, F. J., & Francis, F. (2020). Does the Infectious Status of Aphids Influence Their Preference Towards Healthy, Virus-Infected and Endophytically Colonized Plants? Insects, 11(7), 435. https://doi.org/10.3390/insects11070435