Transgenic Ectopic Overexpression of Broad Complex (BrC-Z2) in the Silk Gland Inhibits the Expression of Silk Fibroin Genes of Bombyx mori


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Semi-RT-PCR and qRT-PCR
2.3. Transgenic Overexpression of BmBrC-Z2 in the PSG
2.4. Juvenile Hormone Treatment
2.5. Statistical Analysis
3. Results
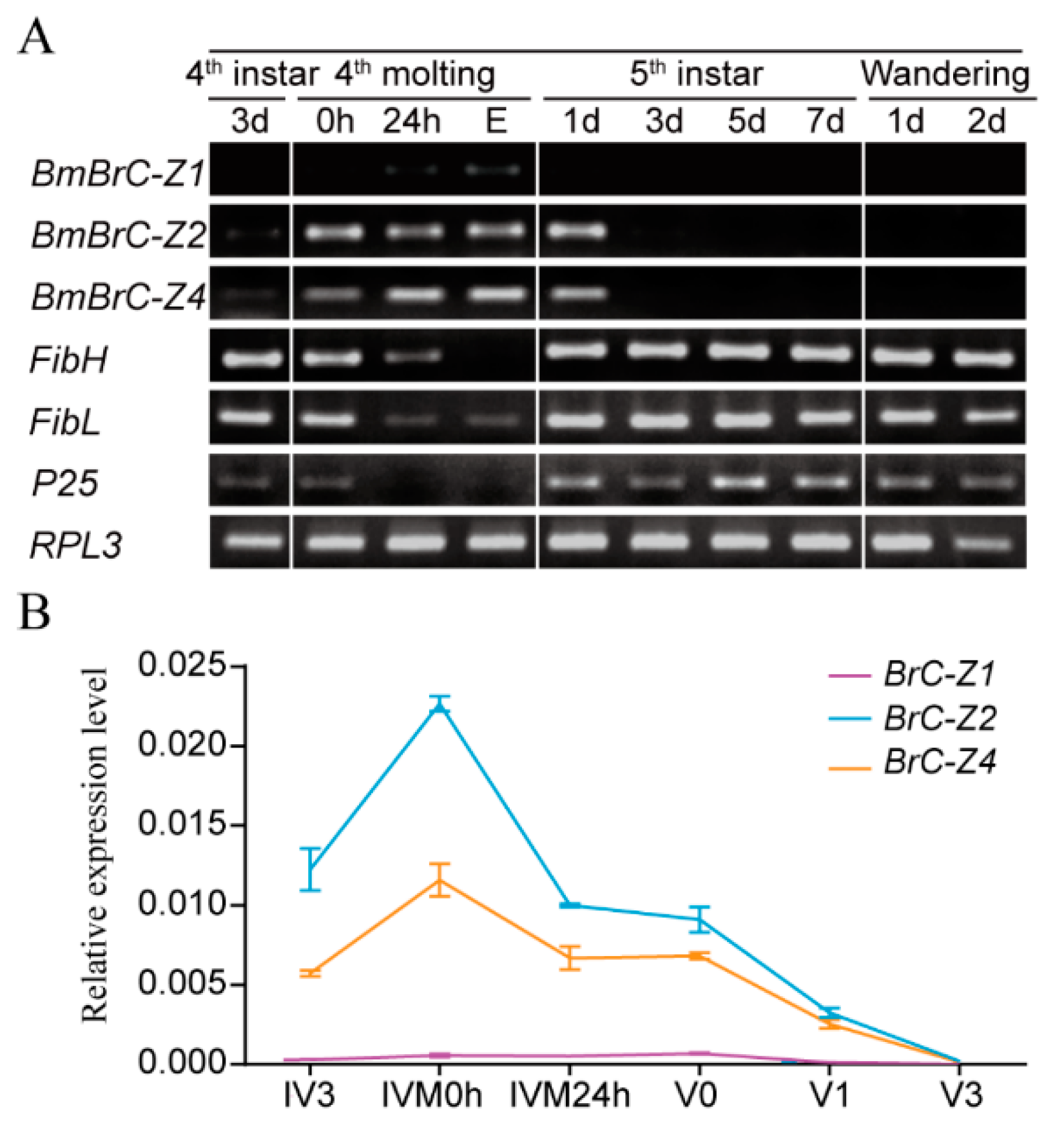
3.1. Expression Patterns of BmBrC-Zs and Fibroins in the Silk Gland
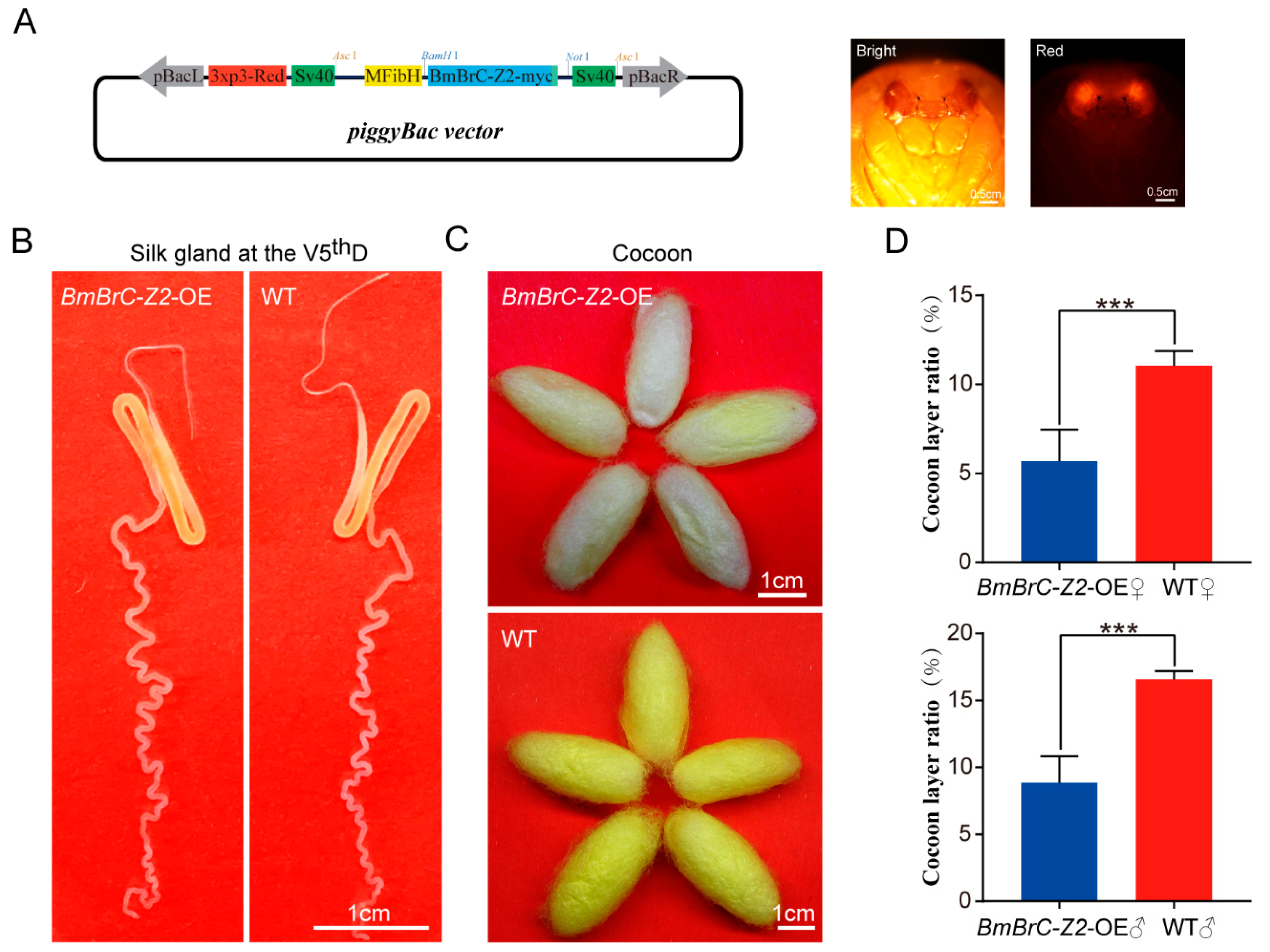
3.2. Transgenic Ectopic Overexpression of BmBrC-Z2 in the PSG
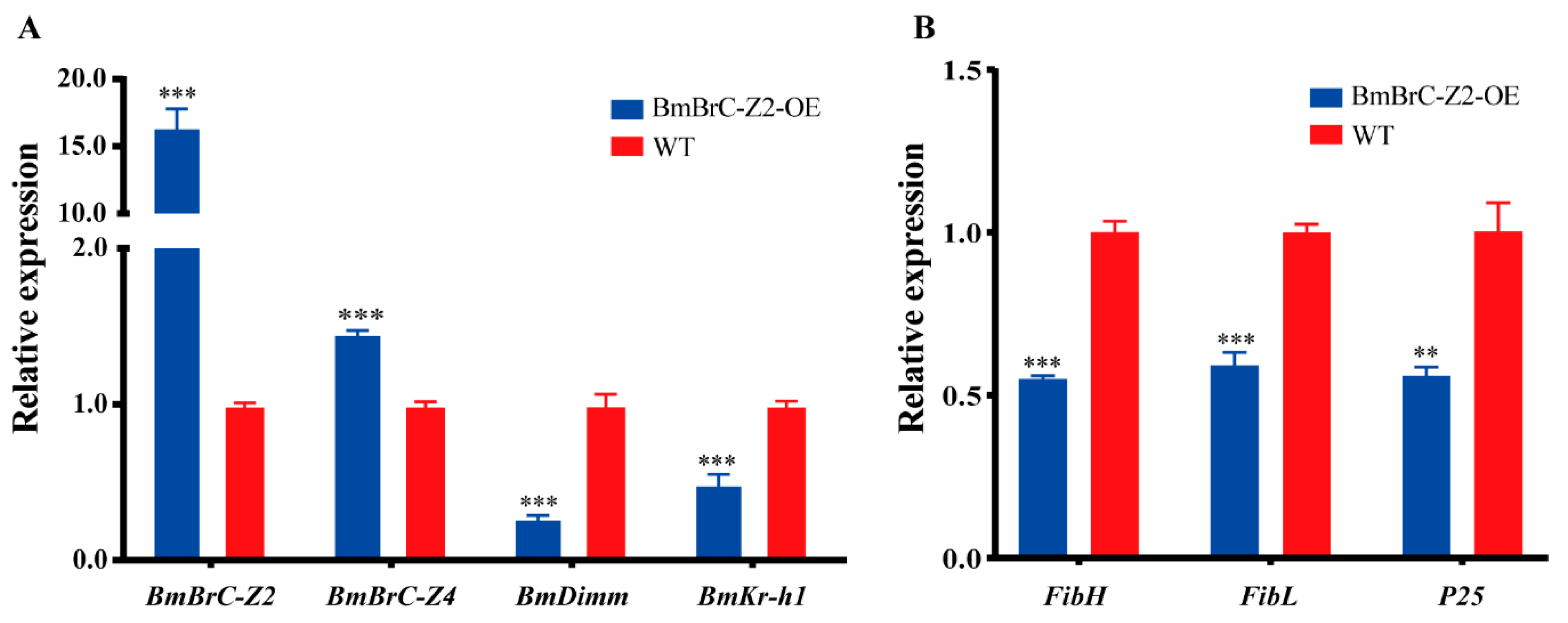
3.3. Transgenic Overexpression of BmBrC-Z2 Downregulates Gene Expression
3.4. Exogenous JH Analog Can Rescue the Phenotype of the Transgenic Line Overexpressing BmBrC-Z2
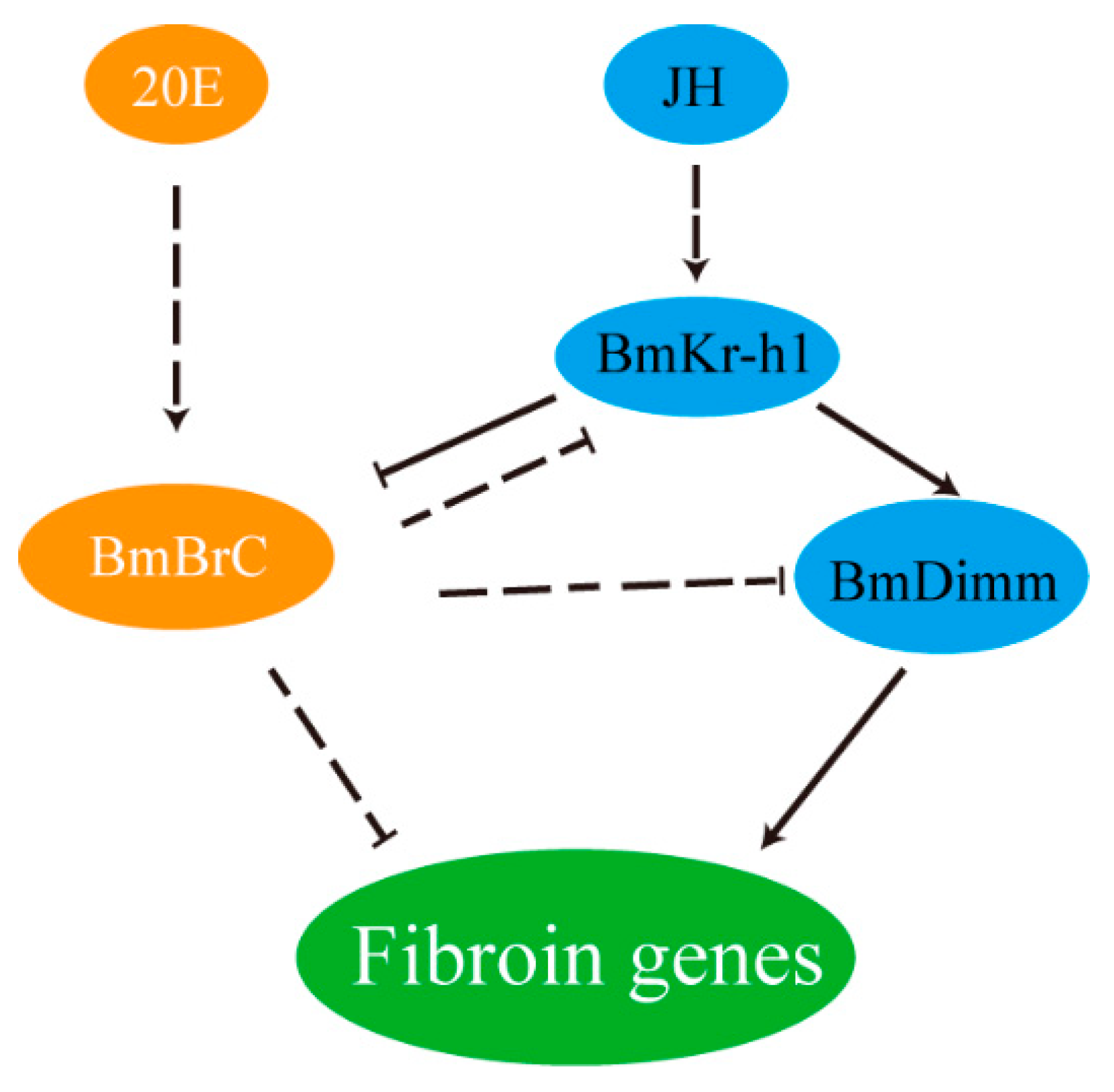
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Thummel, C.S. Ecdysone-regulated puff genes 2000. Insect Biochem. Mol. Biol. 2002, 32, 113–120. [Google Scholar] [CrossRef]
- Reza, A.M.; Kanamori, Y.; Shinoda, T.; Shimura, S.; Mita, K.; Nakahara, Y.; Kiuchi, M.; Kamimura, M. Hormonal control of a metamorphosis-specific transcriptional factor Broad-Complex in silkworm. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Dubrovsky, E.B. Hormonal cross talk in insect development. Trends Endocrinol. Metab. 2005, 16, 6–11. [Google Scholar] [CrossRef]
- Xia, Q.; Li, S.; Feng, Q. Advances in silkworm studies accelerated by the genome sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef] [PubMed]
- Takasu, Y.; Yamada, H.; Tsubouchi, K. Isolation of three main sericin components from the cocoon of the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2002, 66, 2715–2718. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Hino, R.; Ogawa, S.; Iizuka, M.; Adachi, T.; Shimizu, K.; Sotoshiro, H.; Yoshizato, K. A germline transgenic silkworm that secretes recombinant proteins in the sericin layer of cocoon. Transgenic Res. 2007, 16, 449–465. [Google Scholar] [CrossRef]
- Takasu, Y.; Hata, T.; Uchino, K.; Zhang, Q. Identification of Ser2 proteins as major sericin components in the non-cocoon silk of Bombyx mori. Insect Biochem. Mol. Biol. 2010, 40, 339–344. [Google Scholar] [CrossRef]
- Peng, Z.; Yang, X.; Liu, C.; Dong, Z.; Wang, F.; Wang, X.; Hu, W.; Zhang, X.; Zhao, P.; Xia, Q. Structural and Mechanical Properties of Silk from Different Instars of Bombyx mori. Biomacromolecules 2019, 20, 1203–1216. [Google Scholar] [CrossRef]
- Inoue, S.; Tanaka, K.; Arisaka, F.; Kimura, S.; Ohtomo, K.; Mizuno, S. Silk fibroin of Bombyx mori is secreted, assembling a high molecular mass elementary unit consisting of H-chain, L-chain, and P25, with a 6:6:1 molar ratio. J. Biol. Chem. 2000, 275, 40517–40528. [Google Scholar] [CrossRef]
- Maekawa, H.; Suzuki, Y. Repeated turn-off and turn-on of fibroin gene transcription during silk gland development of Bombyx mori. Dev. Biol. 1980, 78, 394–406. [Google Scholar] [CrossRef]
- Hu, W.; Liu, C.; Cheng, T.; Li, W.; Wang, N.; Xia, Q. Histomorphometric and transcriptomic features characterize silk glands’ development during the molt to intermolt transition process in silkworm. Insect Biochem. Mol. Biol. 2016, 76, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Tsuda, M.; Takiya, S.; Hirose, S.; Suzuki, E.; Kameda, M.; Ninaki, O. Tissue-specific transcription enhancement of the fibroin gene characterized by cell-free systems. Proc. Natl. Acad. Sci. USA 1986, 83, 9522–9526. [Google Scholar] [CrossRef]
- Hui, C.C.; Suzuki, Y.; Kikuchi, Y.; Mizuno, S. Homeodomain binding sites in the 5’ flanking region of the Bombyx mori silk fibroin light-chain gene. J. Mol. Biol. 1990, 213, 395–398. [Google Scholar] [CrossRef]
- Takiya, S.; Hui, C.C.; Suzuki, Y. A contribution of the core-promoter and its surrounding regions to the preferential transcription of the fibroin gene in posterior silk gland extracts. EMBO J. 1990, 9, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Durand, B.; Drevet, J.; Couble, P. P25 gene regulation in Bombyx mori silk gland: Two promoter-binding factors have distinct tissue and developmental specificities. Mol. Cell Biol. 1992, 12, 5768–5777. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nony, P.; Prudhomme, J.C.; Couble, P. Regulation of the P25 gene transcription in the silk gland of Bombyx. Biol. Cell. 1995, 84, 43–52. [Google Scholar] [CrossRef]
- Takiya, S.; Kokubo, H.; Suzuki, Y. Transcriptional regulatory elements in the upstream and intron of the fibroin gene bind three specific factors POU-M1, Bm Fkh and FMBP-1. Biochem. J. 1997, 321 Pt 3, 645–653. [Google Scholar] [CrossRef]
- Guo, T.Q.; Wang, J.Y.; Wang, S.P.; Guo, X.Y.; Huang, K.W.; Huang, J.T.; Lu, C.D. Loss of posterior silk gland transcription specificity of fibroin light chain promoter due to absence of 41 bp sequence containing possible inhibitor binding sites. Acta Biochim. Biophys. Sin. (Shanghai) 2005, 37, 819–825. [Google Scholar] [CrossRef]
- Shimizu, K.; Ogawa, S.; Hino, R.; Adachi, T.; Tomita, M.; Yoshizato, K. Structure and function of 5’-flanking regions of Bombyx mori fibroin heavy chain gene: Identification of a novel transcription enhancing element with a homeodomain protein-binding motif. Insect Biochem. Mol. Biol. 2007, 37, 713–725. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, T.; Jin, S.; Guo, Y.; Wu, Y.; Liu, D.; Xu, X.; Sun, Y.; Li, Z.; He, H.; et al. Genome-wide open chromatin regions and their effects on the regulation of silk protein genes in Bombyx mori. Sci. Rep. 2017, 7, 12919. [Google Scholar] [CrossRef]
- Mach, V.; Takiya, S.; Ohno, K.; Handa, H.; Imai, T.; Suzuki, Y. Silk gland factor-1 involved in the regulation of Bombyx sericin-1 gene contains fork head motif. J. Biol. Chem. 1995, 270, 9340–9346. [Google Scholar] [CrossRef] [PubMed]
- Ohno, K.; Sawada, J.; Takiya, S.; Kimoto, M.; Matsumoto, A.; Tsubota, T.; Uchino, K.; Hui, C.C.; Sezutsu, H.; Handa, H.; et al. Silk gland factor-2, involved in fibroin gene transcription, consists of LIM homeodomain, LIM-interacting, and single-stranded DNA-binding proteins. J. Biol. Chem. 2013, 288, 31581–31591. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Fukuta, M.; Takiya, S.; Matsuno, K.; Xu, X.; Suzuki, Y. Promoter of the POU-M1/SGF-3 gene involved in the expression of Bombyx silk genes. J. Biol. Chem. 1994, 269, 2733–2742. [Google Scholar] [PubMed]
- Liu, L.; Wang, Y.; Li, Y.; Guo, P.; Liu, C.; Li, Z.; Wang, F.; Zhao, P.; Xia, Q.; He, H. Insights into the repression of fibroin modulator binding protein-1 on the transcription of fibroin H-chain during molting in Bombyx mori. Insect Biochem. Mol. Biol. 2019, 104, 39–49. [Google Scholar] [CrossRef]
- Zhou, C.; Zha, X.; Shi, P.; Zhao, P.; Wang, H.; Zheng, R.; Xia, Q. Nuclear hormone receptor BmFTZ-F1 is involved in regulating the fibroin heavy chain gene in the silkworm, Bombyx mori. Biochim. Biophys. Acta 2016, 1860, 2529–2536. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, C.; Li, Q.; Hu, W.; Zhou, M.; Nie, H.; Zhang, Y.; Peng, Z.; Zhao, P.; Xia, Q. Basic helix-loop-helix transcription factor Bmsage is involved in regulation of fibroin H-chain gene via interaction with SGF1 in Bombyx mori. PLoS ONE 2014, 9, e94091. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, C.; Jiang, L.; Li, Q.Y.; Zhou, M.; Cheng, T.; Mita, K.; Xia, Q. A juvenile hormone transcription factor Bmdimm-fibroin H chain pathway is involved in the synthesis of silk protein in silkworm, Bombyx mori. J. Biol. Chem. 2015, 290, 972–986. [Google Scholar] [CrossRef]
- Konopova, B.; Jindra, M. Broad-Complex acts downstream of Met in juvenile hormone signaling to coordinate primitive holometabolan metamorphosis. Development 2008, 135, 559–568. [Google Scholar] [CrossRef]
- Huang, J.H.; Lozano, J.; Belles, X. Broad-complex functions in postembryonic development of the cockroach Blattella germanica shed new light on the evolution of insect metamorphosis. Biochim. Biophys. Acta 2013, 1830, 2178–2187. [Google Scholar] [CrossRef]
- Chaharbakhshi, E.; Jemc, J.C. Broad-complex, tramtrack, and bric-a-brac (BTB) proteins: Critical regulators of development. Genesis 2016, 54, 505–518. [Google Scholar] [CrossRef]
- Kayukawa, T.; Nagamine, K.; Ito, Y.; Nishita, Y.; Ishikawa, Y.; Shinoda, T. Kruppel Homolog 1 Inhibits Insect Metamorphosis via Direct Transcriptional Repression of Broad-Complex, a Pupal Specifier Gene. J. Biol. Chem. 2016, 291, 1751–1762. [Google Scholar] [CrossRef] [PubMed]
- Nishita, Y.; Takiya, S. Structure and expression of the gene encoding a Broad-Complex homolog in the silkworm, Bombyx mori. Gene 2004, 339, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Ijiro, T.; Urakawa, H.; Yasukochi, Y.; Takeda, M.; Fujiwara, Y. cDNA cloning, gene structure, and expression of Broad-Complex (BR-C) genes in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2004, 34, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Lin, Y.; Liu, H.; Shen, G.; Luo, J.; Zhang, H.; Peng, Z.; Chen, E.; Xing, R.; Han, C.; et al. The Broad Complex isoform 2 (BrC-Z2) transcriptional factor plays a critical role in vitellogenin transcription in the silkworm Bombyx mori. Biochim. Biophys. Acta 2014, 1840, 2674–2684. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, H.; Yang, C.; Gu, J.; Shen, G.; Zhang, H.; Chen, E.; Han, C.; Zhang, Y.; Xu, Y.; et al. The POU homeodomain transcription factor POUM2 and broad complex isoform 2 transcription factor induced by 20-hydroxyecdysone collaboratively regulate vitellogenin gene expression and egg formation in the silkworm Bombyx mori. Insect Mol. Biol. 2017, 26, 496–506. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, S. 20-hydroxyecdysone enhances the expression of the chitinase 5 via Broad-Complex Zinc-Finger 4 during metamorphosis in silkworm, Bombyx mori. Insect Mol. Biol. 2017, 26, 243–253. [Google Scholar] [CrossRef]
- Wang, H.; Iwanaga, M.; Kawasaki, H. Activation of BMWCP10 promoter and regulation by BR-C Z2 in wing disc of Bombyx mori. Insect Biochem. Mol. Biol. 2009, 39, 615–623. [Google Scholar] [CrossRef]
- Deng, H.; Niu, K.; Zhang, J.; Feng, Q. BmBR-C Z4 is an upstream regulatory factor of BmPOUM2 controlling the pupal specific expression of BmWCP4 in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2015, 66, 42–50. [Google Scholar] [CrossRef]
- Mai, T.; Chen, S.; Lin, X.; Zhang, X.; Zou, X.; Feng, Q.; Zheng, S. 20-hydroxyecdysone positively regulates the transcription of the antimicrobial peptide, lebocin, via BmEts and BmBR-C Z4 in the midgut of Bombyx mori during metamorphosis. Dev. Comp. Immunol. 2017, 74, 10–18. [Google Scholar] [CrossRef]
- Ma, H.; Abbas, M.N.; Zhang, K.; Hu, X.; Xu, M.; Liang, H.; Kausar, S.; Yang, L.; Cui, H. 20-Hydroxyecdysone regulates the transcription of the lysozyme via Broad-Complex Z2 gene in silkworm, Bombyx mori. Dev. Comp. Immunol. 2019, 94, 66–72. [Google Scholar] [CrossRef]
- Ali, A.; Bovilla, V.R.; Mysarla, D.K.; Siripurapu, P.; Pathak, R.U.; Basu, B.; Mamillapalli, A.; Bhattacharya, S. Knockdown of Broad-Complex Gene Expression of Bombyx mori by Oligopyrrole Carboxamides Enhances Silk Production. Sci. Rep. 2017, 7, 805. [Google Scholar] [CrossRef] [PubMed]
- Zhao, A.; Zhao, T.; Zhang, Y.; Xia, Q.; Lu, C.; Zhou, Z.; Xiang, Z.; Nakagaki, M. New and highly efficient expression systems for expressing selectively foreign protein in the silk glands of transgenic silkworm. Transgenic Res. 2010, 19, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Sun, Y.; Wu, Y.; Cheng, T.; Liu, C. Bmsage is involved in the determination of cell number in the silk gland of Bombyx mori. Insect Biochem. Mol. Biol. 2019, 113, 103205. [Google Scholar] [CrossRef] [PubMed]
- Kayukawa, T.; Jouraku, A.; Ito, Y.; Shinoda, T. Molecular mechanism underlying juvenile hormone-mediated repression of precocious larval-adult metamorphosis. Proc. Natl. Acad. Sci. USA 2017, 114, 1057–1062. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purpose | Gene | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|---|
| RT-PCR | BmBrC-Z1 | CCCAAGAAGATTACAGATGCG | AGGTGGCTGGTTAGGGTG |
| RT-PCR | BmBrC-Z2 | TCGCTGACAAACACGCTG | ATGGTAAGAACGGCGGAC |
| RT-PCR | BmBrC-Z4 | GCCACAAGGTCTTCCGCA | AAGAGCCAGCGGAAGGAT |
| RT-PCR | FibH | CAGCATCAGTTCGGTTCC | GACTCGTTACCGTCGGAATC |
| RT-PCR | FibL | ATACCGATTGGTCACATAACAG | GCAGATAGATGGGCGATAA |
| RT-PCR | P25 | AGCCGCTGTGGCAGTTTTG | TAGGTGGCGTTGAAGTATGG |
| RT-PCR | RPL3 | TCGTCATCGTGGTAAGGTCAA | TTTGTATCCTTTGCCCTTGGT |
| qRT-PCR | BmBrC-Z2 | TCGCTGACAAACACGCTG | ATGGTAAGAACGGCGGAC |
| qRT-PCR | BmBrC-Z4 | GCCACAAGGTCTTCCGCA | AAGAGCCAGCGGAAGGAT |
| qRT-PCR | FibH | TCTGTGTCATCTGCTTCATCTCG | TATCCAGGACGAAGTAAGAAACAA |
| qRT-PCR | FibL | ATACCGATTGGTCACATAACAG | GCAGATAGATGGGCGATAA |
| qRT-PCR | P25 | AGCCGCTGTGGCAGTTTTG | TAGGTGGCGTTGAAGTATGG |
| qRT-PCR | BmKr-h1 | CATCGTTTTCAACATTTTGGCGAG | CACATCACTTTACCATCGGCAGC |
| qRT-PCR | BmDimm | CGTGGAACCCGCATTTGTA | AACCTCGGCAATCCAGTCG |
| qRT-PCR | RPL3 | TTCGTACTGGCTCTTCTCGT | CAAAGTTGATAGCAATTCCCT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cong, J.; Tao, C.; Zhang, X.; Zhang, H.; Cheng, T.; Liu, C. Transgenic Ectopic Overexpression of Broad Complex (BrC-Z2) in the Silk Gland Inhibits the Expression of Silk Fibroin Genes of Bombyx mori. Insects 2020, 11, 374. https://doi.org/10.3390/insects11060374
Cong J, Tao C, Zhang X, Zhang H, Cheng T, Liu C. Transgenic Ectopic Overexpression of Broad Complex (BrC-Z2) in the Silk Gland Inhibits the Expression of Silk Fibroin Genes of Bombyx mori. Insects. 2020; 11(6):374. https://doi.org/10.3390/insects11060374
Chicago/Turabian StyleCong, Jiangshan, Cuicui Tao, Xuan Zhang, Hui Zhang, Tingcai Cheng, and Chun Liu. 2020. "Transgenic Ectopic Overexpression of Broad Complex (BrC-Z2) in the Silk Gland Inhibits the Expression of Silk Fibroin Genes of Bombyx mori" Insects 11, no. 6: 374. https://doi.org/10.3390/insects11060374
APA StyleCong, J., Tao, C., Zhang, X., Zhang, H., Cheng, T., & Liu, C. (2020). Transgenic Ectopic Overexpression of Broad Complex (BrC-Z2) in the Silk Gland Inhibits the Expression of Silk Fibroin Genes of Bombyx mori. Insects, 11(6), 374. https://doi.org/10.3390/insects11060374