Biology and Prevalence in Northern Italy of Verrallia aucta (Diptera, Pipunculidae), a Parasitoid of Philaenus spumarius (Hemiptera, Aphrophoridae), the Main Vector of Xylella fastidiosa in Europe

 , , and
, , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Dissections
2.1.1. Field Collection
2.1.2. Dissections
2.2. Molecular Detection of Verrallia aucta
2.2.1. Primer Design
2.2.2. DNA Extraction
2.2.3. Diagnostic PCRs
2.3. Rearing of Verrallia aucta and Philaenus spumarius
2.4. Data Analysis
3. Results
3.1. Molecular Detection of Verrallia aucta
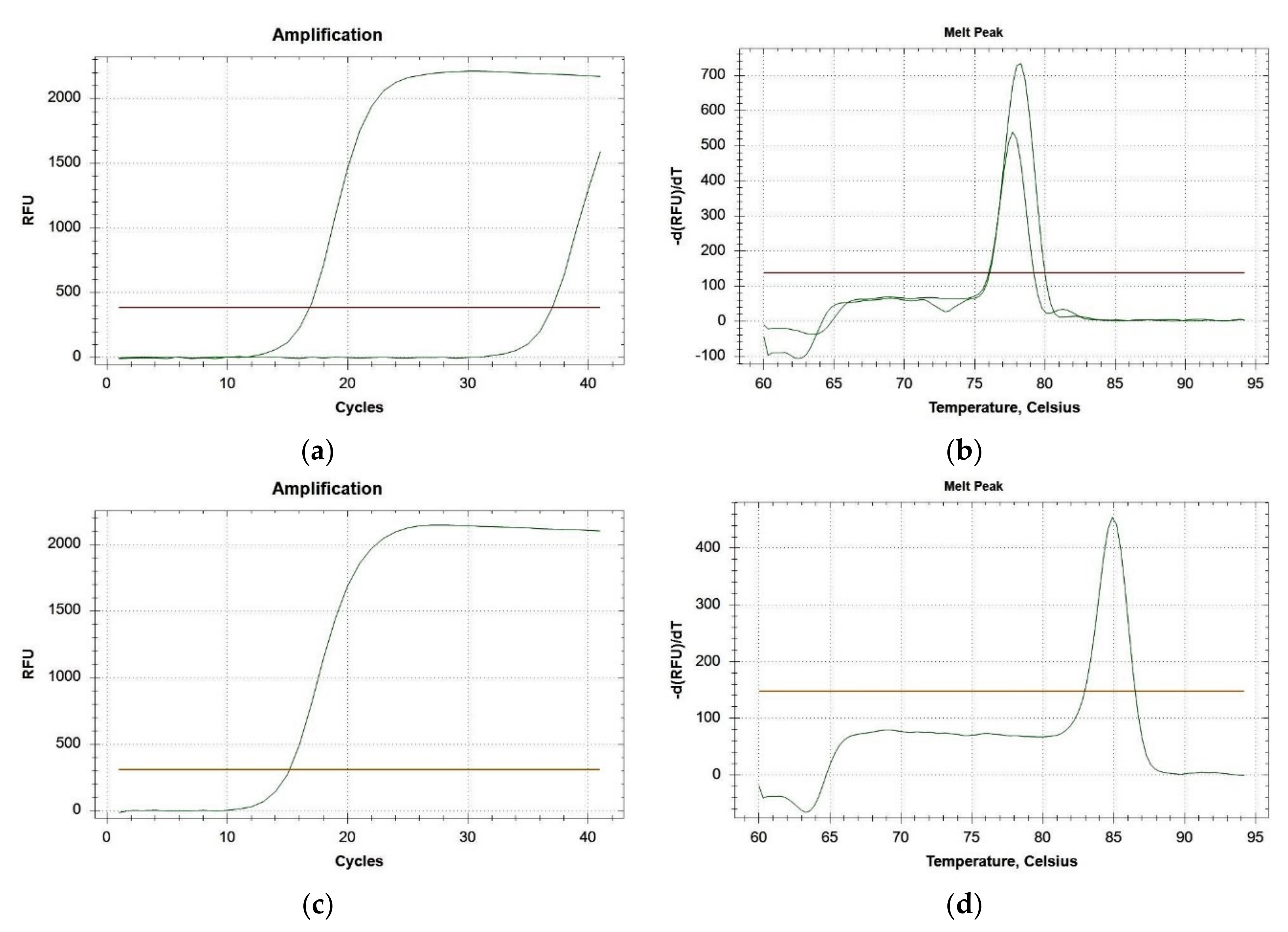
3.1.1. Primer Specificity
3.1.2. Real-Time PCRs
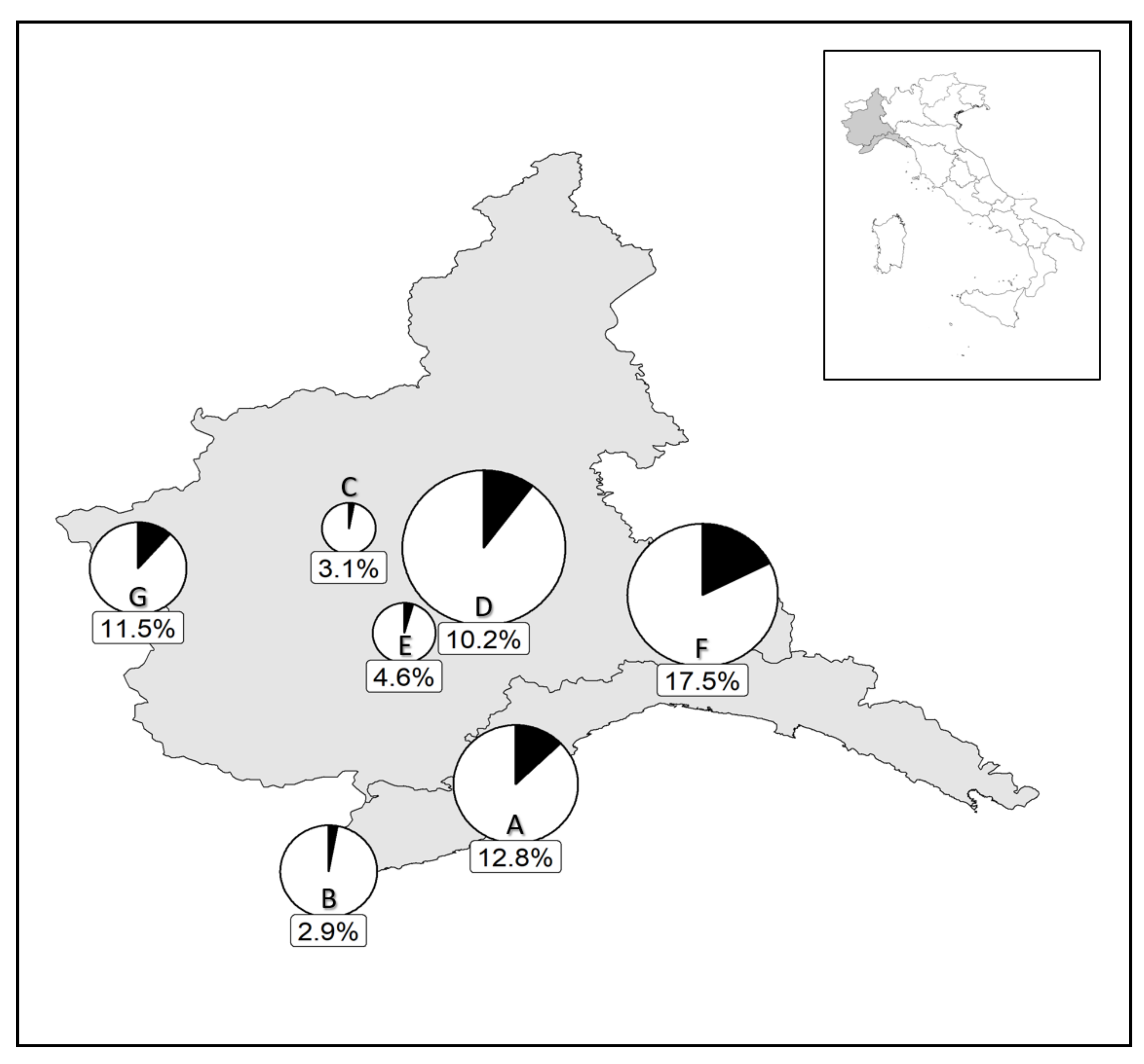
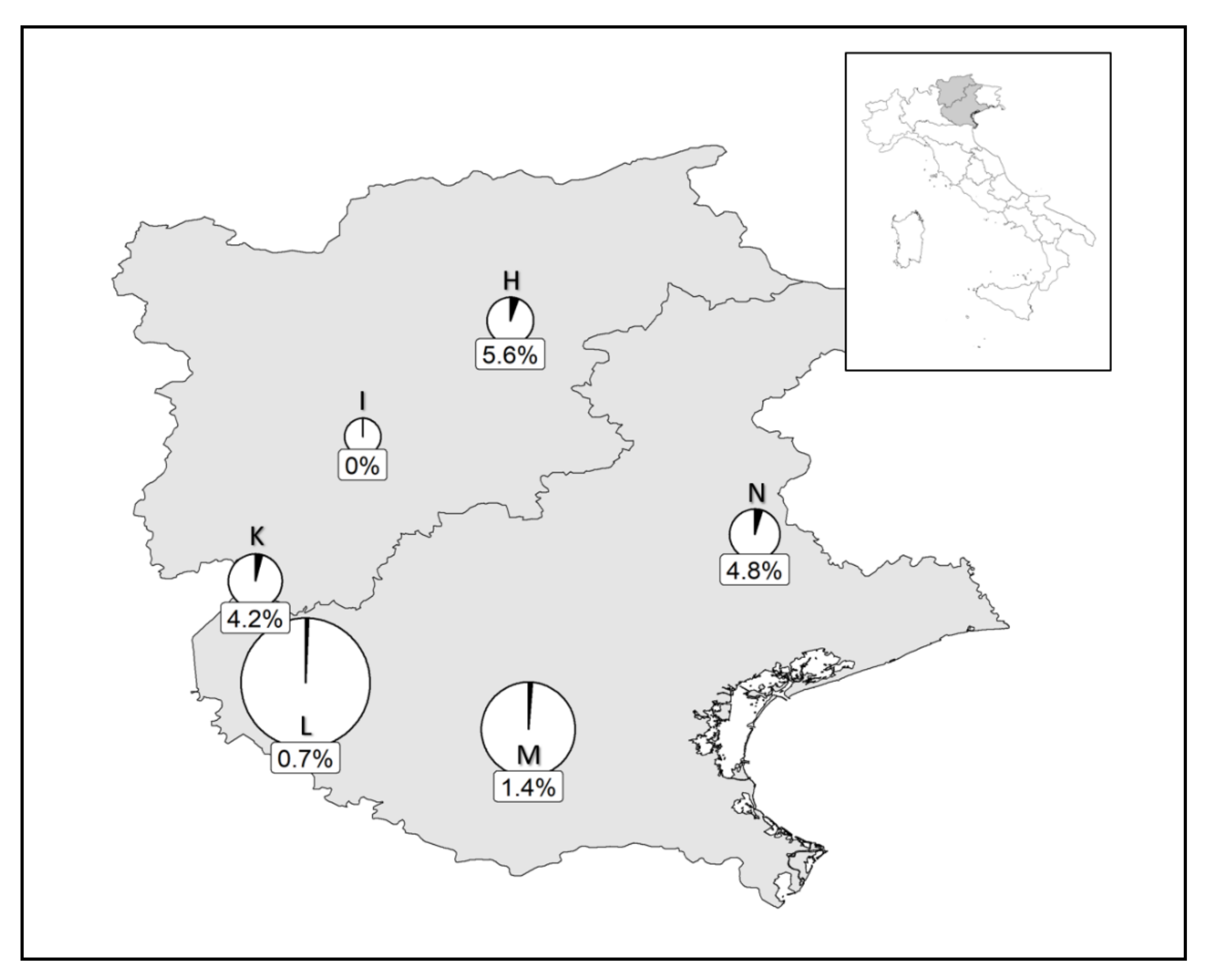
3.2. Prevalence of the Parasitoid in the Field
3.2.1. Philaenus spumarius
3.2.2. Neophilaenus campestris
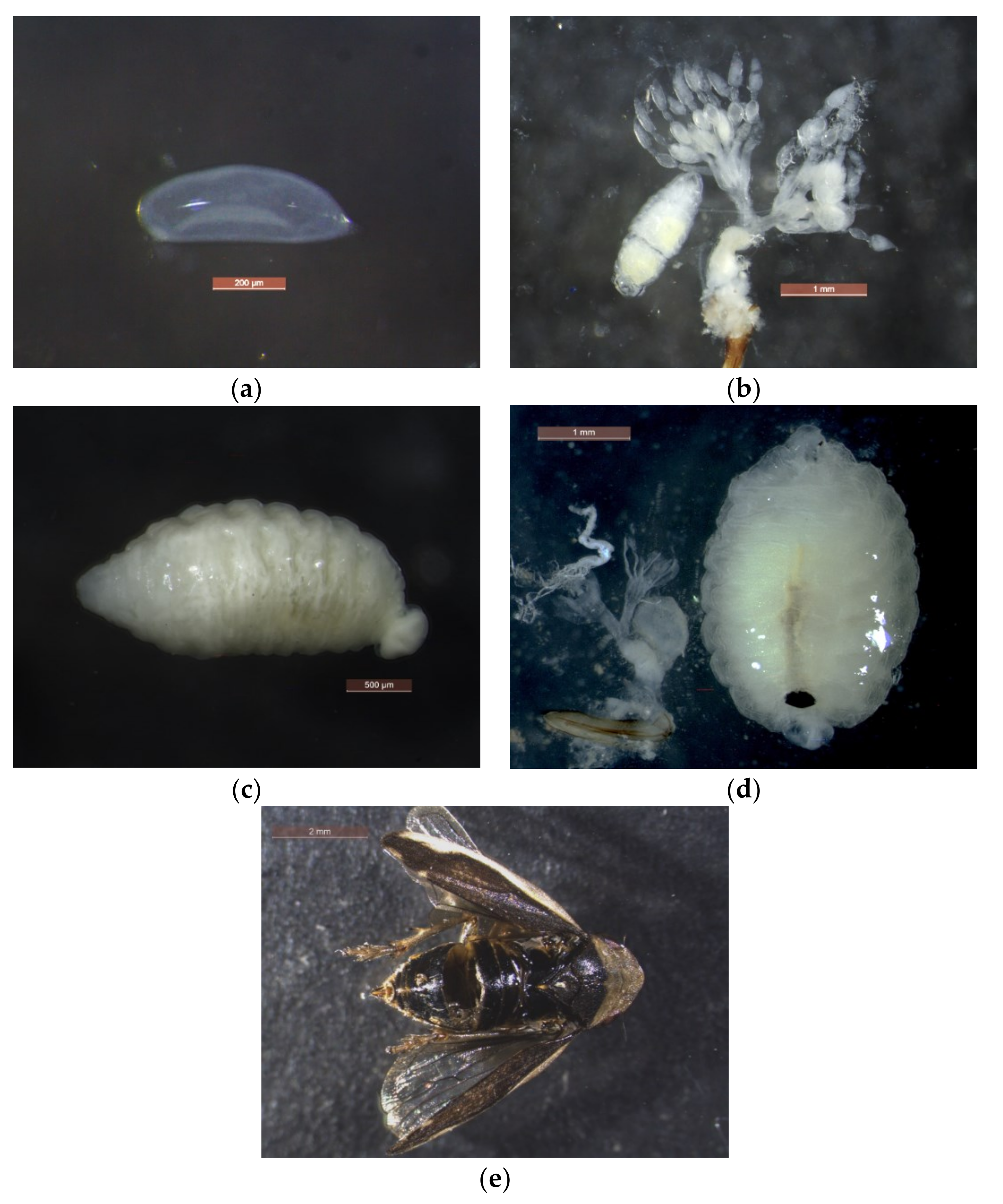
3.3. Biology of Verrallia aucta
3.3.1. Preimaginal Stages
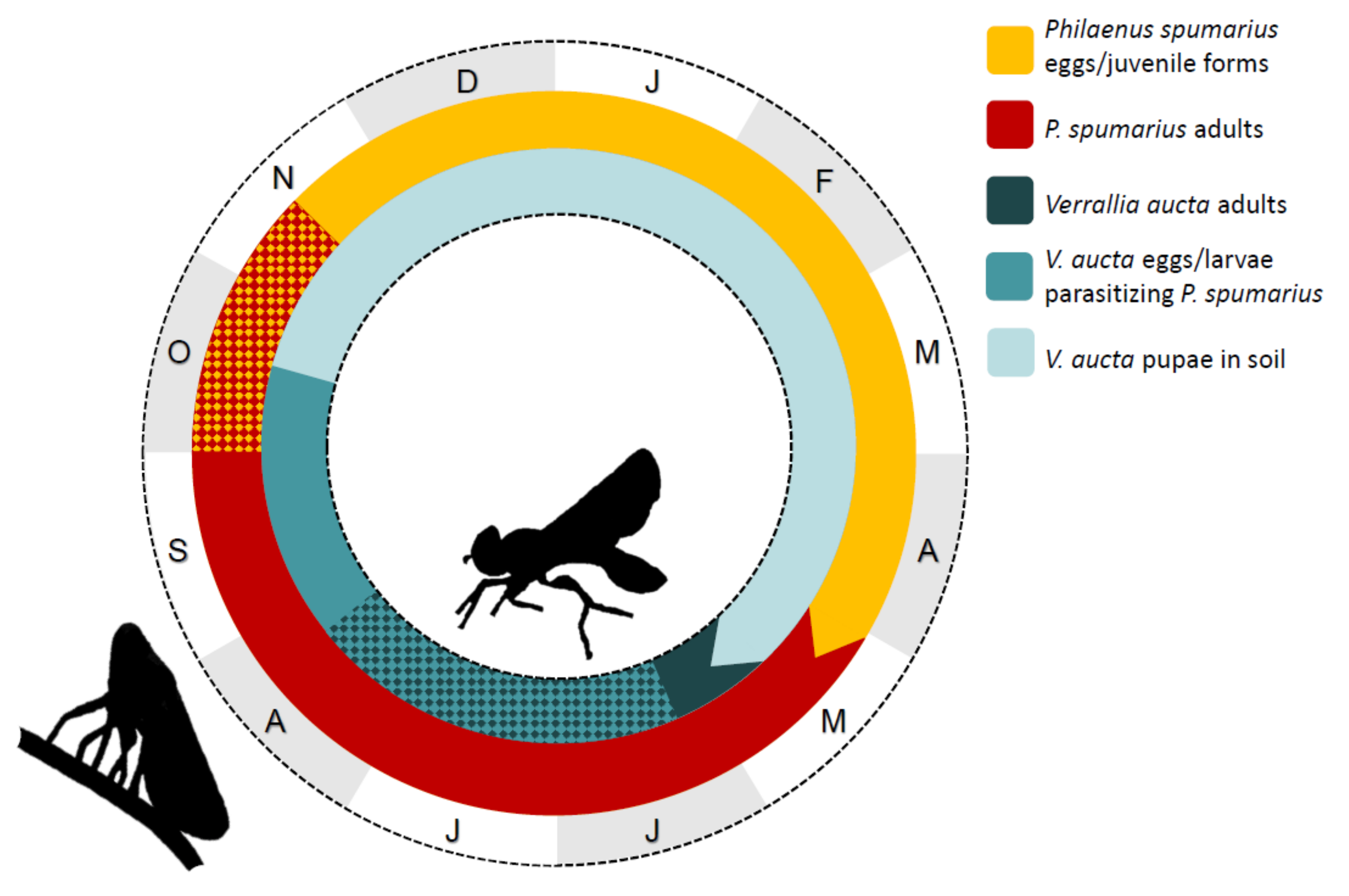
3.3.2. Life Cycle
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Redak, R.; Purcell, A.; Lopes, J.; Blua, M.; Mizell, R.; Andersen, P. The biology of xylem fluid-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology. Annu. Rev. Entomol. 2004, 49, 243–270. [Google Scholar] [CrossRef]
- Saponari, M.; Boscia, D.; Loconsole, G.; Palmisano, F.; Savino, V.; Potere, O.; Martelli, G. New hosts of Xylella fastidiosa strain CoDiRO in Apulia. J. Plant Pathol. 2014, 96, 611. [Google Scholar]
- Cavalieri, V.; Altamura, G.; Fumarola, G.; di Carolo, M.; Saponari, M.; Cornara, D.A.-O.; Bosco, D.; Dongiovanni, C.A.-O. Transmission of Xylella fastidiosa subspecies pauca sequence type 53 by different insect species. Insects 2019, 10, 324. [Google Scholar] [CrossRef] [PubMed]
- Zajac, M.A.; Wilson, M.C. The effects of nymphal feeding by the meadow spittlebug, Philaenus spumarius (L.) on strawberry yield and quality. Crop Prot. 1984, 3, 167–175. [Google Scholar] [CrossRef]
- Yurtsever, S. On the polymorphic meadow spittlebug, Philaenus spumarius (L.)(Homoptera: Cercopidae). Turk. J. Zool. 2000, 24, 447–460. [Google Scholar]
- Cornara, D.; Saponari, M.; Zeilinger, A.; de Stradis, A.; Boscia, D.; Loconsole, G.; Bosco, D.; Martelli, G.; Almeida, R.; Porcelli, F. Spittlebugs as vectors of Xylella fastidiosa in olive orchards in Italy. J. Pest Sci. 2017, 90, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Moralejo, E.; Borras, D.; Gomila, M.; Montesinos, M.; Adrover, F.; Juan, A.; Nieto, A.; Olmo, D.; Segui, G.; Landa, B. Insights into the epidemiology of Pierce’s disease in vineyards of Mallorca, Spain. Plant Pathol. 2019, 68, 1458–1471. [Google Scholar] [CrossRef]
- Cornara, D.; Bosco, D.; Fereres, A. Philaenus spumarius: When an old acquaintance becomes a new threat to European agriculture. J. Pest Sci. 2018, 91, 957–972. [Google Scholar] [CrossRef]
- Purcell, A.H. Homopteran transmission of xylem-inhabiting bacteria. In Advances in Disease Vector Research; Springer: Berlin, Germany, 1990; pp. 243–266. [Google Scholar]
- Cruaud, A.; Gonzalez, A.; Godefroid, M.; Nidelet, S.; Streito, J.; Thuillier, J.; Rossi, J.; Santoni, S.; Rasplus, J. Using insects to detect, monitor and predict the distribution of Xylella fastidiosa: A case study in Corsica. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Dongiovanni, C.; Altamura, G.; Di Carolo, M.; Fumarola, G.; Saponari, M.; Cavalieri, V. Evaluation of efficacy of different insecticides against Philaenus spumarius L., vector of Xylella fastidiosa in olive orchards in Southern Italy, 2015–2017. Arthropod Manag. Tests 2018, 43, 34. [Google Scholar] [CrossRef][Green Version]
- Morente, M.; Cornara, D.; Plaza, M.; Duran, J.; Capiscol, C.; Trillo, R.; Ruiz, M.; Ruz, C.; Sanjuan, S.; Pereira, J.; et al. Distribution and relative abundance of insect vectors of Xylella fastidiosa in olive groves of the Iberian peninsula. Insects 2018, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Tsagkarakis, A.; Afentoulis, D.; Matared, M.; Thanou, Z.; Stamatakou, G.; Kalaitzaki, A.; Tzobanoglou, D.; Goumas, D.; Trantas, E.; Zarboutis, I.; et al. Identification and seasonal abundance of Auchenorrhyncha with a focus on potential insect vectors of Xylella fastidiosa in olive orchards in three regions of Greece. J. Econ. Entomol. 2018, 111, 2536–2545. [Google Scholar] [CrossRef] [PubMed]
- Di Serio, F.; Bodino, N.; Cavalieri, V.; Demichelis, S.; Di Carolo, M.; Dongiovanni, C.; Fumarola, G.; Gilioli, G.; Guerrieri, E.; Picciotti, U. Collection of data and information on biology and control of vectors of Xylella fastidiosa. EFSA Support. Publ. 2019, 16. [Google Scholar] [CrossRef]
- Bodino, N.; Cavalieri, V.; Dongiovanni, C.; Plazio, E.; Saladini, M.; Volani, S.; Simonetto, A.; Fumarola, G.; Di Carolo, M.; Porcelli, F.; et al. Phenology, seasonal abundance and stage-structure of spittlebug (Hemiptera: Aphrophoridae) populations in olive groves in Italy. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Bodino, N.; Cavalieri, V.; Dongiovanni, C.; Saladini, M.; Simonetto, A.; Volani, S.; Plazio, E.; Altamura, G.; Tauro, D.; Gilioli, G.; et al. Spittlebugs of mediterranean olive groves: Host-plant exploitation throughout the year. Insects 2020, 11, 130. [Google Scholar] [CrossRef]
- Panzavolta, T.; Bracalini, M.; Croci, F.; Ghelardini, L.; Luti, S.; Campigli, S.; Goti, E.; Marchi, R.; Tiberi, R.; Marchi, G. Philaenus italosignus a potential vector of Xylella fastidiosa: Occurrence of the spittlebug on olive trees in Tuscany (Italy). Bull. Insectol. 2019, 72, 317–320. [Google Scholar]
- Santoiemma, G.; Tamburini, G.; Sanna, F.; Mori, N.; Marini, L. Landscape composition predicts the distribution of Philaenus spumarius, vector of Xylella fastidiosa, in olive groves. J. Pest Sci. 2019, 92, 1101–1109. [Google Scholar] [CrossRef]
- Hasbroucq, S.; Casarin, N.; Czwienczek, E.; Bragard, C.; Grégoire, J.-C. Distribution, adult phenology and life history traits of potential insect vectors of Xylella fastidiosa in Belgium. Belg. J. Entomol. 2020, 92, 1–21. [Google Scholar]
- Mesmin, X.; Chartois, M.; Genson, G.; Rossi, J.; Cruaud, A.; Rasplus, J. Ooctonus vulgatus (Hymenoptera, Mymaridae), a potential biocontrol agent to reduce populations of Philaenus spumarius (Hemiptera, Aphrophoridae ) the main vector of Xylella fastidiosa in Europe. PeerJ 2020, 8, 8591. [Google Scholar] [CrossRef]
- Liccardo, A.; Fierro, A.; Garganese, F.; Picciotti, U.; Porcelli, F. A biological control model to manage the vector and the infection of Xylella fastidiosa on olive trees. PLoS ONE 2020, 15, 232363. [Google Scholar] [CrossRef]
- Ables, J.R. Feeding behavior of an assassin bug, Zelus renardii. Ann. Entomol. Soc. Am. 1978, 71, 476–478. [Google Scholar] [CrossRef]
- Benhadi-Marín, J.; Villa, M.; Pereira, L.F.; Rodrigues, I.; Morente, M.; Baptista, P.; Pereira, J.A. A guild-based protocol to target potential natural enemies of Philaenus spumarius (Hemiptera: Aphrophoridae), a vector of Xylella fastidiosa (Xanthomonadaceae): A case study with spiders in the olive grove. Insects 2020, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, J.B. The Biology of Pipunculidae (Diptera) Parasitising Some British Cercopidae (Homoptera). Physiol. Entomol. 1969, 44, 17–24. [Google Scholar] [CrossRef]
- Whittaker, J.B. Density regulation in a population of Philaenus spumarius (L.)(Homoptera: Cercopidae). J. Anim. Ecol. 1973, 42, 163–172. [Google Scholar] [CrossRef]
- de Jong, Y.; Verbeek, M.; Michelsen, V.; de Place Bjørn, P.; Los, W.; Steeman, F.; Bailly, N.; Basire, C.; Chylarecki, P.; Stloukal, E. Fauna Europaea–all European animal species on the web. Biodivers. Data J. 2014, 2, 4034. [Google Scholar] [CrossRef]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, 67–72. [Google Scholar] [CrossRef]
- Kehlmaier, C.; Assmann, T. Molecular analysis meets morphology-based systematics-a synthetic approach for Chalarinae (Insecta: Diptera: Pipunculidae). Syst. Entomol. 2010, 35, 181–195. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, 115. [Google Scholar] [CrossRef]
- Marzachi, C.; Bosco, D. Relative quantification of chrysanthemum yellows (16Sr I) phytoplasma in its plant and insect host using real-time polymerase chain reaction. Mol. Biotechnol. 2005, 30, 117–127. [Google Scholar] [CrossRef]
- Swallow, W.H. Group testing for estimating infection rates and probabilities of disease transmission. Phytopathology 1985, 75, 882–889. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Coe, R.L. Handbooks for the Identification of British Insects: Vol. X, Part 2(c). Diptera Pipunculidae; Royal Entomological Society of London: London, UK, 1966; pp. 8–10. [Google Scholar]
- de Meyer, M. Preliminary database on the distribution of Pipunculidae (Diptera) in Europe. Pros. 81h Airfoil. E.]. S. Bruss. 1992, 91–100. [Google Scholar]
- de Meyer, M.; Bruyn, L.d. On the phenology of some Pipunculidae (Diptera) in Belgium. Bull. Ann. Société R. Belge d’Entomologie 1984, 120, 123–131. [Google Scholar]
- Wiegert, R.G. Population energetics of meadow spittlebugs (Philaenus spumarius L.) as affected by migration and habitat. Ecol. Monogr. 1964, 34, 218–241. [Google Scholar] [CrossRef]
- Prior, K.M.; Hellmann, J.J. Does enemy loss cause release? A biogeographical comparison of parasitoid effects on an introduced insect. Ecology 2013, 94, 1015–1024. [Google Scholar] [CrossRef]
- Weaver, C.R.; King, D.R. Meadow spittlebug, Philaenus leucophthalmus (L.). Ohio ARDC Res. Bull. 1954, 71, 1–99. [Google Scholar]
- Grant, J.F.; Lambdin, P.L.; Follum, R.A. Infestation levels and seasonal incidence of the meadow spittlebug (Homoptera: Cercopidae) on musk thistle in Tennessee. J. Agric. Entomol. 1998, 15, 83–91. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Region | Latitude | Longitude | Site Code | Altitudinal Zonation |
|---|---|---|---|---|---|
| Arnasco | Liguria | 44.0766 | 8.1173 | A | plain–foothill |
| Finale Ligure | Liguria | 44.1811 | 8.3634 | A | plain–foothill |
| Ventimiglia | Liguria | 43.8073 | 7.5858 | B | plain–foothill |
| Chieri | Piemonte | 45.0154 | 7.7931 | C | plain–foothill |
| Druento | Piemonte | 45.1290 | 7.5880 | C | plain–foothill |
| Grugliasco | Piemonte | 45.0730 | 7.5874 | C | plain–foothill |
| Asti | Piemonte | 44.9194 | 8.1982 | D | plain–foothill |
| Cocconato | Piemonte | 45.0826 | 8.0594 | D | plain–foothill |
| Cisterna d’Asti | Piemonte | 44.8241 | 8.0117 | E | plain–foothill |
| Monteu Roero | Piemonte | 44.7784 | 7.9402 | E | plain–foothill |
| Vezza d’Alba | Piemonte | 44.7586 | 8.0220 | E | plain–foothill |
| Paderna | Piemonte | 44.8261 | 8.8947 | F | plain–foothill |
| Prali | Piemonte | 44.8759 | 7.0575 | G | montane |
| Cesana | Piemonte | 44.9530 | 6.7936 | G | montane |
| Sestriere | Piemonte | 44.9472 | 6.9054 | G | montane |
| Castelrotto | Trentino-Alto Adige | 46.5672 | 11.5560 | H | montane |
| Castelrotto San Michele | Trentino-Alto Adige | 46.5787 | 11.6021 | H | montane |
| Compaccio | Trentino-Alto Adige | 46.5410 | 11.6170 | H | montane |
| Mezzocorona | Trentino-Alto Adige | 46.2132 | 11.1455 | I | plain–foothill |
| Malcesine | Veneto | 45.7777 | 10.8206 | K | plain–foothill |
| Bussolengo | Veneto | 45.4475 | 10.8628 | L | plain–foothill |
| Montorio | Veneto | 45.4741 | 11.0768 | L | plain–foothill |
| Mezzane di Sotto | Veneto | 45.4837 | 11.1187 | L | plain–foothill |
| Nanto | Veneto | 45.4350 | 11.5791 | M | plain–foothill |
| Arquà Petrarca | Veneto | 45.2701 | 11.7395 | M | plain–foothill |
| Lozzo Atestino | Veneto | 45.2874 | 11.6195 | M | plain–foothill |
| Castello Roganzuolo | Veneto | 45.9182 | 12.3310 | N | plain–foothill |
| Primer | Sequence (5′–3′) | Target | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| VaITS2 Fw | TGCTGCTTGGACTACATATGG | ITS2 | 183 | This work |
| VaITS2 Rv | AACGCATGGCACTAAACGAA | |||
| VaCOI Fw | TGGAGGATTCGGAAACTGAC | COI | 308 | This work |
| VaCOI Rv | AGGTGATTCCTGTAGACCGC | |||
| Mq Fw | AACGGCTACCACATCCAAGG | 18S | 98 | [30] |
| Mq Rv | GCCTCGGATGAGTCCCG |
| Region | Site | Year | MAY | JUN | JUL | AUG | SEP | OCT | NOV | DEC | Altitudinal Zonation |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Liguria | A | 2016 | - | - | - | - | 16.7% d (6) | 0% d (8) | 0% d (6) | 0% d (3) | plain–foothill |
| 2017 | 0% d (5) | 0% d (13) | 0% d (10) | 16.7% d (12) | 10% d (10) | 0% d (11) | 0% d (10) | 0% d (7) | |||
| 2019 | - | - | 17.6% p (102) | - | - | - | - | - | |||
| B | 2018 | - | - | - | 2.9% p (104) | - | - | - | - | ||
| Piemonte | C | 2016 | - | - | - | - | 0% d (1) | - | - | - | |
| 2017 | - | 0% d (6) | 0% d (1) | - | - | - | - | - | |||
| 2018 | - | 0% d (5) | - | - | - | - | - | - | |||
| 2019 | 0% d (5) | 33.3% d (3) | 0% d (7) | 0% d (2) | 0% dp (7) | - | - | - | |||
| D | 2016 | - | - | - | 8.7% d (23) | 0% d (15) | 7.7% d (13) | 0% d (33) | 0% d (3) | ||
| 2017 | 0% d (30) | 0% d (11) | 15.4% d (13) | 14.3% d (7) | 0% d (11) | 0% d (16) | 0% d (12) | - | |||
| 2018 | - | 0% d (3) | - | - | 15.6% dp (77) | - | - | - | |||
| 2019 | - | 15.9%dp (69) | 0% p (2) | - | 2.9% dp (35) | - | - | - | |||
| E | 2018 | - | - | - | - | 0% p (27) | - | - | - | ||
| 2019 | - | 40% dp (5) | 0% p (4) | - | 0% dp (8) | - | - | - | |||
| F | 2018 | - | 0% d (3) | - | - | 19.9% dp (136) | - | - | - | ||
| 2019 | - | 14.1% dp (64) | - | - | 14.6% dp (48) | - | - | - | |||
| G | 2016 | - | - | 0% d (9) | 0% d (6) | 0% d (2) | - | - | - | montane | |
| 2017 | - | - | 7.1% d (14) | 25% d (12) | 16.7% d (12) | - | - | - | |||
| 2018 | - | 0% d (1) | 4.5% d (22) | 14.3% d (14) | 0% d (2) | - | - | - | |||
| 2019 | - | - | 0% d (3) | 42.9% d (7) | - | - | - | - |
| Region | Year | Site | JUL | AUG | SEP | Altitudinal Zonation |
|---|---|---|---|---|---|---|
| Trentino-Alto Adige | 2018 | H | 0%d (5) | 7.7%d (13) | - | montane |
| I | 0%d (11) | - | - | plain–foothill | ||
| Veneto | 2019 | K | 0%p (3) | - | 4.8%p (21) | |
| L | 3.7%p (27) | 0%p (43) | 0%p (68) | |||
| M | 0%p (5) | 0%p (32) | 2.8%p (36) | |||
| N | - | 8.3%p (12) | 0%p (9) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molinatto, G.; Demichelis, S.; Bodino, N.; Giorgini, M.; Mori, N.; Bosco, D. Biology and Prevalence in Northern Italy of Verrallia aucta (Diptera, Pipunculidae), a Parasitoid of Philaenus spumarius (Hemiptera, Aphrophoridae), the Main Vector of Xylella fastidiosa in Europe. Insects 2020, 11, 607. https://doi.org/10.3390/insects11090607
Molinatto G, Demichelis S, Bodino N, Giorgini M, Mori N, Bosco D. Biology and Prevalence in Northern Italy of Verrallia aucta (Diptera, Pipunculidae), a Parasitoid of Philaenus spumarius (Hemiptera, Aphrophoridae), the Main Vector of Xylella fastidiosa in Europe. Insects. 2020; 11(9):607. https://doi.org/10.3390/insects11090607
Chicago/Turabian StyleMolinatto, Giulia, Stefano Demichelis, Nicola Bodino, Massimo Giorgini, Nicola Mori, and Domenico Bosco. 2020. "Biology and Prevalence in Northern Italy of Verrallia aucta (Diptera, Pipunculidae), a Parasitoid of Philaenus spumarius (Hemiptera, Aphrophoridae), the Main Vector of Xylella fastidiosa in Europe" Insects 11, no. 9: 607. https://doi.org/10.3390/insects11090607
APA StyleMolinatto, G., Demichelis, S., Bodino, N., Giorgini, M., Mori, N., & Bosco, D. (2020). Biology and Prevalence in Northern Italy of Verrallia aucta (Diptera, Pipunculidae), a Parasitoid of Philaenus spumarius (Hemiptera, Aphrophoridae), the Main Vector of Xylella fastidiosa in Europe. Insects, 11(9), 607. https://doi.org/10.3390/insects11090607